DNA VIRAL INFECTIONS
Adenovirus Infection
Adenoviral enteritis has been documented in commercial rabbits in Hungary. Peak losses occurred at 6-8 weeks of age. Profuse diarrhea was observed in severely affected animals, with low mortality.
There was a dramatic increase in the numbers of E. coli in the small intestine and cecum in rabbits that succumbed to the disease, suggesting a contributing role in pathogenesis. Severely affected animals were dehydrated, with fluid contents in the cecum. The adenovirus was isolated from the intestinal wall and gut contents, spleen, kidney, and lung when inoculated onto rabbit kidney cell cultures. A significant rise in adenoviral antibody levels was detected in convalescent sera. To date, confirmed cases of adenoviral enteritis in rabbits appear to be confined to Europe, although naturally occurring seroconversion to bovine adenovirus (type 1) antigen has been detected in meat rabbits from multiple commercial colonies in Quebec, Canada.Herpesvirus Infections
There are 4 known herpesviruses of rabbits, known as leporid herpesvirus (LHV) -1, -2, -3, and -4. LHV-1, -2, and -3 belong to the subfamily Gammaherpesvirinae, whereas LHV-4 belongs to the genus Simplex of the subfamily Alphaherpesvirinae. In addition, rabbits are naturally susceptible to Human herpesvirus 1 (Herpes simplex 1). Among the 4 rabbit herpesviruses, LHV-1 (cottontail herpesvirus) and LHV-3 (Herpesvirus sylvilagus) are indigenous to Sylvilagus rabbits and do not infect Oryctolagus rabbits. They are covered in this chapter because they are traditionally listed among rabbit viruses. LHV-2 and LHV-4 naturally infect domestic rabbits, but only LHV-4 has clinical significance.
Leporid Herpesvirus 1 and 3 Infection
Both LHV-1 and LHV-3 were isolated from primary kidney cell cultures of weanling cottontail (Sylvilagus) rabbits. LHV-3 enjoyed a brief period of interest as a model of human Epstein-Barr virus infection.
Inoculation of young cottontail rabbits with H. sylvilagus (LHV- 3) by the parenteral route produces a chronic infection with persistent viremia and atypical lymphocytosis. Within 6-8 weeks following inoculation, changes consistent with a lymphoproliferative disease are observed grossly and microscopically. Histological examination reveals alterations that vary from lymphoid hyperplasia to lymphoma. Other lesions include myocarditis, interstitial pneumonia, and myositis. In the malignant form of the disease, diffuse infiltration of various tissues with immature lymphocytes commonly occurs. Although H. sylvilagus replicates in kidney cells prepared from the domestic rabbit, attempts to infect New Zealand White rabbits with the virus have been unsuccessful. No disease has been reported in cottontail rabbits naturally infected with LHV-1, and no attempts to infect domestic rabbits with LHV-1 have been reported.Leporid Herpesvirus 2 Infection
LHV-2 has also been termed “Virus III" and Herpesvirus cuniculi. The virus was originally discovered during serial testicular transfer following inoculation of blood from a human varicella patient, and subsequently isolated from rabbit cell kidney cultures. It appears to be a subclinical infection among seropositive rabbits under natural conditions, but experimental intracerebral inoculation results in nonsuppurative encephalitis with intranuclear inclusion bodies.
Leporid Herpesvirus 4 Infection
Systemic infections with a LHV-4 have been observed in several commercial rabbitries in Canada. Outbreaks have mysteriously occurred among isolated rabbit populations, as well as following introduction of outside rabbits to rabbitries. The disease, which affects animals of various ages, is characterized by sudden onset with mortality up to 30%, with highest mortality among young rabbits.
Animals are frequently found dead, with no previous evidence of illness. Clinical signs include anorexia, conjunctivitis, periocular and facial swelling, subcutaneous swellings, respiratory distress, diarrhea, and abortion.
Additional gross findings include disseminated cutaneous hemorrhagic macules, pulmonary congestion and edema, hydrothorax, hydropericardium, and multifocal hemorrhage in spleen, kidney, stomach, and intestine. Microscopic findings include multifocal necrohemor- rhagic lesions in skin (Fig. 6.6), spleen, adrenals, stomach, intestine, heart, kidney, uterus, and liver. Pulmonary hemorrhage and edema are prominent. Intranuclear eosinophilic and amphophilic inclusions are present in multiple tissues, particularly respiratory epithelium, and multinucleate syncytia are present in some affected tissues (Fig. 6.7). Experimental intranasal inoculation resulted in periocular swelling, facial dermatitis, oculonasal discharge, and severe necrohemorrhagic bronchopneumonia. Inclusion bodies and syncytia were noted in multiple tissues. Notably, encephalitis has not been found in affected rabbits. Differential diagnosis must rule out caliciviral infection (rabbit hemorrhagic disease), but LHV-4 can be readily differentiated by the presence of characteristic intranuclear inclusion bodies and syncytia. Periocular swelling and acute death may also mimic myxomatosis, which is associated with intra- cytoplasmic pox viral inclusions. The sporadic incidence and severity of the disease suggest that rabbits may not be the natural host for this virus. LHV-4 is closely related to bovine herpesvirus 2, which infects a variety of ruminant species. No serosurveys have been performed among domestic rabbits.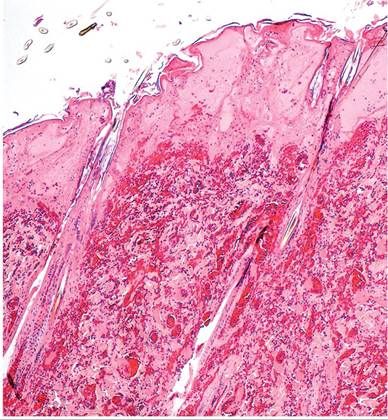
FIG. 6.6. Integument from a rabbit with leporid herpesvirus 4 infection, depicting necrosis of the epidermis and papillary dermis with underlying dermal hemorrhage. (Source: Brash et al. 2010. Reproduced with permission from Canadian Veterinary Medical Association.)
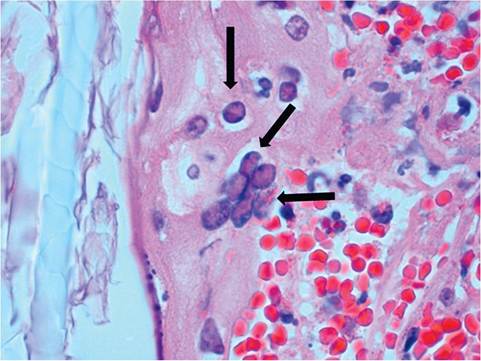
FIG. 6.7. Leporid herpesvirus 4 infection of the skin from a rabbit, depicting a multinucleate syncytium and intranuclear inclusion bodies (arrows) in the follicular epithelium.
(Source: Brash et al. 2010. Reproduced with permission from Canadian Veterinary Medical Association.)Herpes Simplex Virus Infection
For decades, domestic rabbits have served as an animal model for experimental Human herpesvirus-1 (Herpes simplex-1 virus) encephalitis. Sporadic naturally occurring cases of fatal encephalitis due to H. simplex virus infection have been observed in pet rabbits. Affected rabbits may present with conjunctivitis (Fig. 6.8) and neurologic signs. Necropsy reveals nonsuppurative meningoencephalitis with neuronal necrosis and prominent amphophilic intranuclear inclusion bodies in neurons and astroglial cells (Fig. 6.9). Lesions in other organs have not been described. Typical herpesvirus particles have been observed in affected cells by electron microscopy, and H. simplex-1 has been confirmed as the
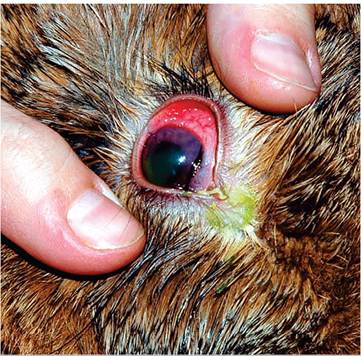
FIG. 6.8. Acute conjunctivitis in a rabbit naturally infected with herpes simplex virus infection following transmission from an infected human. (Source: Muller et al. 2009. Reproduced with permission from the American Veterinary Medical Association.)
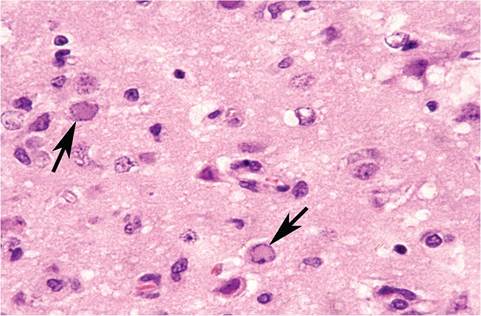
FIG. 6.9. Cerebrum from a rabbit with naturally acquired herpes simplex virus encephalitis. Note the prominent intranuclear inclusion bodies in neurons (arrows) and astroglia.
causative agent. Contact with humans infected with H. simplex virus has been documented in most reported cases in rabbits.
Lapine Parvovirus Infection
A parvovirus has been isolated from clinically normal rabbits in Japan. In a serological survey of commercial rabbits, approximately 60% of animals evaluated had antibody to lapine parvovirus. Oral or intravenous inoculation of 1 month old rabbits with lapine parvovirus resulted in transient depression and anorexia, with no mortality. The virus could be isolated from a variety of organs for up to 2 weeks postinoculation and from the small intestine on day 30 postinoculation.
On microscopic examination, a mild to moderate enteritis was present in the small intestine, with exfoliation of enter- ocytes. In a survey of laboratory rabbits from commercial and private sources in the United States, the majority had relatively high antibody titers to lapine parvovirus. In addition, lapine parvovirus has been isolated from the kidneys of neonatal rabbits. The role (if any) of lapine parvovirus in the enteritis complex is currently unknown.Necrotic hepatitis associated with a parvovirus was reported in Mexico in 1989. Rabbits also had splenic necrosis, and microinfarctions in the myocardium, kidney, and lung believed to be associated with intravascular coagulation. These lesions were reminiscent of rabbit hemorrhagic disease (RHD), but affected rabbits also had severe segmental necrosis of the small intestinal crypts and villi, which is not a feature of RHD. Furthermore, liver lesions contained intranuclear paracrystalline arrays of parvoviral-like virions by electron microscopy, and intranuclear staining with monoclonal antibodies against porcine parvovirus and murine parvovirus (MVM) was present. No other reports of this syndrome have been published, and this case may have been due to activation of parvovirus during the course of an RHD epizootic.
Papillomavirus Infections
Papilloma viruses were once classified as members of the family Papoviridae that included 2 genera, Papillomaviruses and Polyomaviruses, which differ in size, biology, and DNA sequence. That family has been eliminated and papilloma viruses are now classified within the family Papillomaviridae. Rabbits can be infected with 2 papilloma viruses, cottontail rabbit papillomavirus (CRPV) and rabbit oral papilloma virus (ROPV), both of which belong to the genus Kappapapillomavirus. Both of these viruses are used as experimental models of papilloma virus pathogenesis and immunity in the laboratory rabbit.
Cottontail Rabbit Papillomavirus (CRPV) Infection
CRPV, which is also known as Shope papilloma virus, is native to New World Sylvilagus spp.
rabbits, although Oryctolagus is experimentally and naturally susceptible to infection. Papillomaviruses cannot be propagated in cell culture, but infectious clones of viral DNA are extensively used to induce experimental infection of laboratory rabbits.Epizootiology and Pathogenesis
Primarily a benign disease of cottontail rabbits, papillomatosis due to CRPV has occurred in spontaneous outbreaks among domestic rabbits. Insect vectors are the usual means of mechanical spread of the virus from cottontail rabbits to domestic rabbits, and ticks are important for infection among Sylvilagus rabbits. When the CRPV is inoculated into Oryctolagus rabbits, it produces papillomas with a high incidence (75%) of progression to squamous cell carcinomas. Transformation to carcinomas also occurs in Sylvilagus, but to a lesser degree. Papillomas induced by this virus in Oryc- tolagus rabbits produce minimal or no infectious virus, in contrast to papillomas induced in the natural Sylvilagus host or in hares (Lepus spp.). Papillomas typically undergo immune-mediated resolution if they do not progress to carcinomas. Host immunity has been shown to have 2 distinct targets: virus structural antigens, invoking protection against virus reinfection, and tumor antigens, invoking papilloma regression. If rabbits are immunized against the virus, viral immunity does not affect papilloma status, and rabbits remain susceptible to induction of papillomas with infectious DNA. Once tumors have undergone immune-mediated regression, rabbits resist both virus and DNA challenge. Virus DNA can remain latent, with no histologic changes in the epidermis, until nonspecific irritants activate the formation of papillomas.
Pathology
Naturally occurring papillomas on Orycolagus rabbits occur most frequently on the eyelids and ears, which are sites most often accessed by insects. Papillomas

FIG. 6.10. Multiple cutaneous papillomas (horns) projecting from
the lips of a Sylvilagus rabbit naturally infected with cottontail rabbit papilloma virus (CRPV). (Source: Flicker © C Forry wd45/364229280.)
consist of a pedunculated, cornified surface overlying a fleshy central core. Papillomas in Sylvilagus rabbits vary in size and number, up to numerous large cutaneous horns that debilitate the host (Fig. 6.10). Papillomas due to CRPV, which arise in naturally infected Oryctolagus rabbits, tend to be small and few in number (Fig. 6.11). Histologic findings are consistent with squamous papilloma. In those tumors that progress to malignancy, squamous cell carcinomas are locally invasive as well as metastatic to regional lymph nodes and lung. Differential diagnosis must discriminate from spontaneous nonviral papillomas, which tend to arise in haired skin.
Rabbit Oral Papillomavirus (ROPV) Infection
ROPV is distinct from CRPV. The natural host of ROPV is the Oryctolagus rabbit. Like CRPV, ROPV is also used as an experimental model for papilloma virus pathogenesis
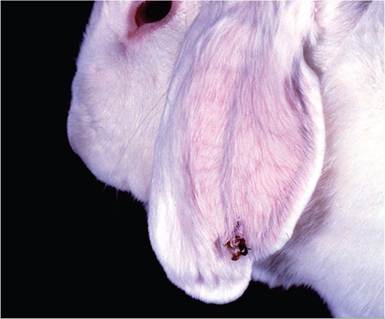
FIG. 6.11. Solitary papilloma on the ear of a laboratory rabbit naturally infected with cottontail rabbit papilloma virus (CRPV). Since CRPV papillomas arise in an abnormal host (domestic rabbit), these tumors do not support virus replication and are prone to malignant transformation.
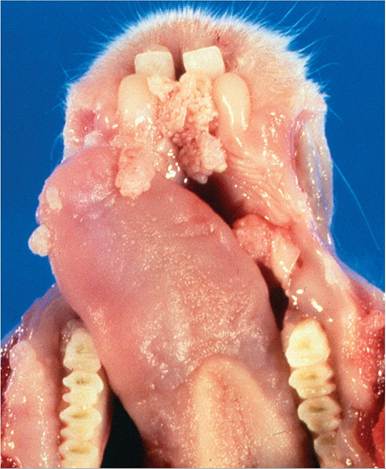
FIG. 6.12. Oral papillomas involving the tongue and gingiva of a
juvenile New Zealand White rabbit naturally infected with rabbit oral papilloma virus. These lesions undergo regression with no tendency toward malignancy.
and immunity. Natural infection is common among domestic rabbits, and infection is spread by direct contact, including from does to their kits.
Lesions most frequently occur in rabbits between 2 and 18 months of age. Pedunculated and sessile lesions are most commonly found along the ventral aspect of the tongue (Fig. 6.12), but may also involve other areas of oral and labial mucosa. These papillomas usually regress spontaneously within a few weeks. They are typical squamous papillomas on microscopic examination. Persistent papillomas of the conjunctiva have also been attributed to ROPV. Basophilic intranuclear inclusions and viral antigen may be present in the stratum spinosum. There are no reports of malignant transformation of oral papillomas in rabbits. Viral DNA can be detected in oral swabs in rabbits without lesions, latent virus remains in epithelium following papilloma regression, and latent virus can be activated by nonspecific injury.
Polyomavirus Infection
Cottontail rabbits are frequently subclinically infected with rabbit kidney vacuolating virus, a nononcogenic member of the Polyomaviridae family. The virus was isolated from CRPV papillomas inoculated into primary kidney cultures derived from Sylvilagus rabbits. This virus has no known pathogenic effect in cottontail or domestic rabbits but can be a contaminant of papilloma virus stocks. Seroconversion to the virus has been found in Sylvilagus rabbits, but antibodies have not been detected against the virus in domestic rabbits. Intranuclear inclusions in renal tubular epithelium, consistent with
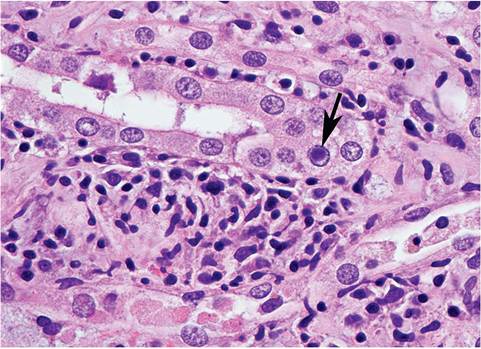
FIG. 6.13. Intranuclear inclusion body (arrow), found as an incidental finding in the renal tubular epithelium of an adult New Zealand White rabbit. These inclusions are suspected to be due to an uncharacterized polyomavirus.
polyomavirus-like inclusions, may be seen as an incidental finding in domestic rabbits (Fig. 6.13), but their etiology has not been defined.
Poxvirus Infections
Rabbits are host to several poxviruses with considerable clinical significance. The family Poxviridae is divided into 2 subfamilies, Entomopoxvirinae (which infect insects) and Chordopoxvirinae (which infect mammals). The latter contains the Leporipoxvirus genus, which includes the lagomorph pathogens known as myxoma, rabbit fibroma, and hare fibroma viruses. These viruses, particularly myxoma and rabbit fibroma viruses, are closely related and are represented by a number of strains with an overlapping spectrum of virulence. Another Leporipoxvirus is squirrel fibroma virus, which is antigenically and genetically closely related to, but distinct from myxoma and rabbit fibroma viruses. Finally, rabbitpox virus is closely related to vaccinia virus in the Orthopox genus. A comprehensive review of poxviral infections of rabbits is available in Brabb and DiGiacomo (2012).
Myxoma Virus Infection: Myxomatosis
Myxomatosis was first recognized in laboratory Orycto- lagus rabbits acquired for experiments in a South American laboratory in the late 19th century. The name “infectious myxomatosis” was used to denote the myxoid appearance of the subcutaneous masses associated with the disease. In the original cases described, the virus was believed to have been transmitted by insect vectors from the relatively resistant tropical forest rabbit (S. braziliensis). Myxomatosis was also recognized in North America in 1930, where outbreaks of the disease occurred in rabbitries in southern California. The brush rabbit (S. bachmani) has been implicated as the reservoir host in that area. Myxomatosis is enzootic in the western
United States, with sporadic cases spreading into domestic rabbits, and is also endemic in wild Sylvilagus and feral Oryctolagus rabbits in South and Central America. Around 1950, a highly virulent South American strain of myxoma virus was introduced into Australia in an effort to reduce (or eliminate) the overpopulation of feral Oryctolagus cuniculus, which had become a major economic problem in that country. Mortality rates of up to 99% subsequently dropped to around 25% within a few years. The dramatic reduction in mortality was related to natural selection for genetically resistant rabbits and the emergence of attenuated strains of the virus. In 1953, myxoma virus was released into France by a citizen who was disenchanted with the wild rabbit problem. The virus subsequently spread to other countries in western Europe, including England, where it is now a well-established enzootic among wild Oryctolagus rabbits. Transmission of the virus is usually mechanical and by arthropod vectors, primarily mosquitoes in the Americas and Australia and fleas in Europe, but also by direct contact and fomites. There is considerable variation in virulence among myxoma virus strains that have evolved from these events. Attenuated strains of myxoma virus and rabbit fibroma virus are used as live vaccines in Europe to protect commercially raised rabbits.
Pathology
Disease severity is highly variable, depending upon virus strain and host species. Sylvilagus rabbits (natural hosts) tend to be resistant, and develop localized cutaneous lesions similar to those induced by fibroma viruses. Following inoculation of Oryctolagus rabbits by an arthropod vector, viral replication results in the development of a primary subcutaneous myxoid mass, usually within 3-4 days. Within 6-8 days, mucopurulent conjunctivitis, facial edema, and multiple cutaneous skin masses (myxomas) are usually observed (Fig. 6.14), often involving the base of the ears. Swelling of the anogenital region is also common, with scrotal edema. In rabbits that die with a peracute form of the disease, the animal may be found dead, and other than redness of the conjunctiva, there may be no other
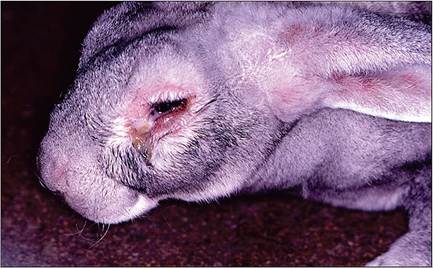
FIG. 6.14. Mucopurulent conjunctivitis, periocular swelling, and facial edema in a rabbit with myxomatosis.
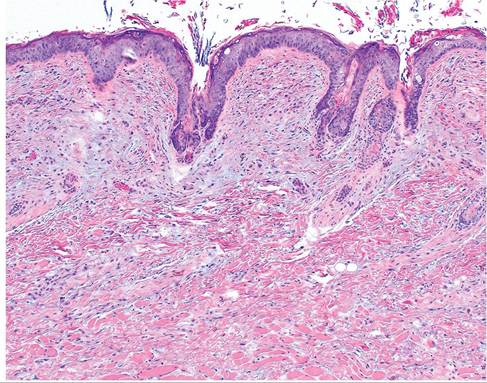
FIG. 6.15. Skin of a rabbit with myxomatosis. Note the bluestaining mucoid material in the superficial dermis.
evidence of disease. Microscopically, there is dermal and subcutaneous proliferation of large, stellate mesenchymal cells (myxoma cells) interspersed within a homogeneous mucinoid matrix (Fig. 6.15), with a sprinkling of inflammatory cells. Hypertrophy and proliferation of endothelial cells occur, and changes in the epithelium overlying the lesions may vary from hyperplasia to degeneration. Intracytoplasmic inclusions are often present in the affected epidermis and in epithelial cells of the conjunctiva (Fig. 6.16), and in some cases may be found in respiratory epithelium. Remarkably, systemic changes are minimal, except lymphoid tissue. These include proliferation of alveolar epithelium, pulmonary hemorrhage, and orchitis in bucks. Myxoma virus is T lymphocytotropic, and viremic dissemination involves lymphocytes and monocytes, as well as rapid local dissemination to draining lymph nodes. Lymph nodes are initially enlarged with formation of syncytia
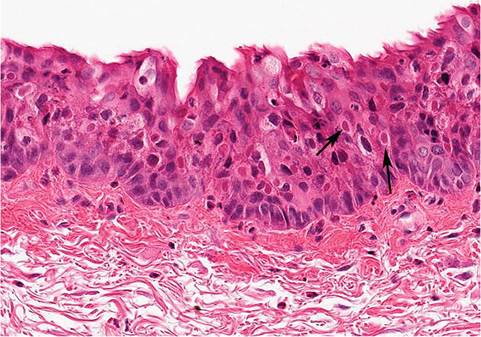
FIG. 6.16. Intracytoplasmic inclusion bodies (arrows) in the conjunctival epithelium of a rabbit with myxomatosis.
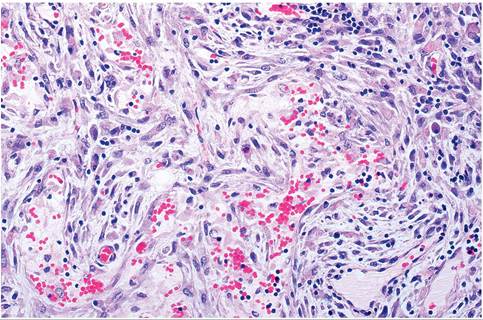
FIG. 6.17. Lymph node of a rabbit with myxomatosis, depicting hypertrophy and hyperplasia of stellate myxoma cells and severe lymphocytic depletion.
among apoptotic lymphoid cells, followed by marked lymphocytic depletion, particularly the T-cell zones. Affected lymph nodes, and to a lesser extent spleen, have hypertrophy and hyperplasia of stellate myxoma cells (Fig. 6.17), as well as focal necrosis, hemorrhage, and proliferative vasculitis. The cause of death in myxomatosis has never been fully defined, and suggested to be related to cytokine-mediated shock. Limited analysis of natural cases suggests that acute sepsis may contribute to mortality due to massive immunodeficiency. Mild disease may occur in rabbits inoculated with live attenuated vaccine strains, or partially immune-vaccinated rabbits exposed to myxoma virus. Under these circumstances, rabbits may develop nodular fibromatous masses of the conjunctiva.
"Amyxomatous" myxomatosis is also recognized, in which rabbits have minimal or small skin nodules, and present with respiratory signs. Rabbits develop pneumonia, and pox inclusions may be visualized in respiratory epithelium (Fig. 6.18). Presumably, this form of myxomatosis is spread through respiratory contact.
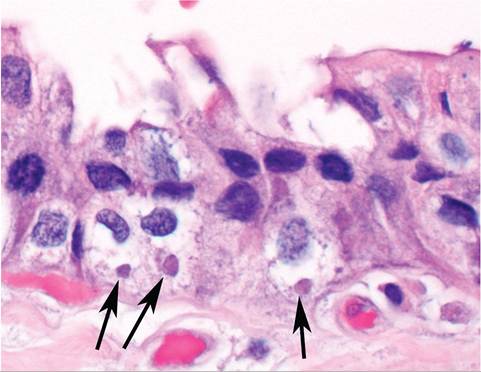
FIG. 6.18. Intracytoplasmic inclusion bodies (arrows) in the respiratory epithelium of a rabbit with “amyxomatous” myxomatosis. (Source: S. Diab, University of California, Davis, CA. Reproduced with permission from S. Diab.)
reservoir host, with spread by insect vectors. In naturally infected cottontail and European rabbits, firm, flattened tumors occur on the legs and feet (Fig. 6.19), sometimes with involvement of the muzzle, periorbital, and perineal regions. These cutaneous tumors may be up to 7 cm in diameter, are usually freely movable, and may persist for several months, and then undergo spontaneous resolution. In young rabbits, fibromatous lesions may arise in abdominal viscera and bone marrow. There is localized fibroblast proliferation, with mononuclear and polymorphonuclear cell infiltration. Affected fibroblasts are characteristically fusiform to polygonal. In European domestic rabbits, subcutaneous masses may vary from myxoid in type to typical fibromas. Large, intracytoplas- mic, eosinophilic inclusion bodies may be present in fibromatous cells (Fig. 6.20) and in epidermal cells overlying the tumors. The typical gross and histological appearance of the circumscribed masses should
Rabbit Fibroma Virus Infection
Rabbit fibroma virus, also known as Shope fibroma virus, is closely related antigenically to myxoma virus and to the hare and squirrel fibroma viruses. Shope fibroma virus was first isolated from a cottontail rabbit (S. flori- danus) in the United States in 1932. The virus is transmissible to Oryctolagus rabbits, producing localized fibromas. Shope fibroma virus infections are relatively widespread in wild cottontail rabbits in the eastern United States and Canada. Infection is usually a benign, self-limiting disease in the wildlife population. The virus may persist for several months within lesions, and mechanical transmission by arthropod vectors appears to be the primary means of spread. In rare occasions, fibromatosis has been diagnosed in commercial rabbit- ries. Wild cottontail rabbits in the area are the likely
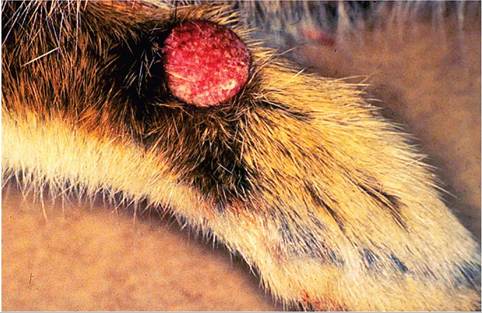
FIG. 6.19. Nodular fibroma on the forelimb of a Sylvilagus rabbit
naturally infected with Shope rabbit fibroma virus.
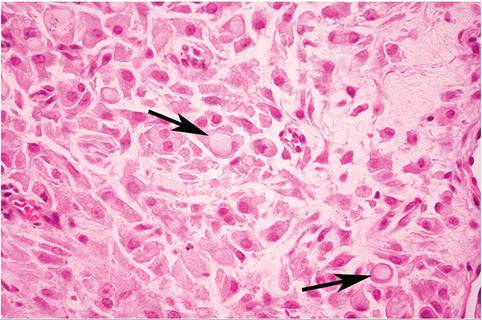
FIG. 6.20. Skin of a Sylvilagus rabbit infected with Shope fibroma virus. There is a dense network of fusiform to polyhedral fibroblasts in the dermis, many of which contain prominent intracytoplasmic inclusion bodies (arrows).
facilitate the differentiation of rabbit fibromatosis from myxomatosis and from the raised, horny, epidermal growths seen in papillomatosis. However, the myxoid forms sometimes seen histologically can be confused with myxomatosis. Considerable variation in microscopic appearance may occur.
Hare Fibroma Virus Infection
Hare fibroma virus is closely related to rabbit fibroma virus and myxoma virus and is an infection of European hares. Hare fibroma virus has been documented in European hares prior to the introduction of myxoma virus to Europe, and outbreaks have been documented in captive-bred European hares. Fibromatous lesions, 1-3 cm in diameter, typically arise on the ears and legs of affected hares, and spontaneously regress in adult hares. Lesions consist of large spindle and stellate cells with prominent intracytoplasmic inclusions. Virus is transmitted by insects and contact. Myxoma virus can also infect hares, resulting in formation of firm nodular cutaneous masses of the head, back, and limbs, which resemble hare fibroma virus-induced lesions.
Rabbitpox (Vaccinia) Virus Infection
Rabbit pox is relatively rare and has generally been associated with inadvertent exposure to vaccinia virus. Recent sequence analysis of rabbitpox virus has confirmed its close relationship to vaccinia virus. During outbreaks of rabbit pox, virus is highly contagious. It is readily transmitted by aerosol, with the respiratory tract being the primary site of replication, followed by viremic dissemination. Papular lesions may occur in the oropharynx, respiratory tract, spleen, and liver. In the "pockless" form, a few pocks were present in the oral cavity, and focal hepatic necrosis, pleuritis, and splenomegaly were observed. Histologic changes include disseminated focal necrosis with leukocytic infiltration in the skin and affected viscera, as well as necrosis of lymphoid tissue. Because of the similarity to smallpox, rabbitpox virus has been used as a model of variola virus infection, thereby emphasizing the possibility of future outbreaks due to the marked ability of natural spread among susceptible rabbits.
More on the topic DNA VIRAL INFECTIONS:
- DNA VIRAL INFECTIONS
- DNA VIRAL INFECTIONS
- DNA VIRAL INFECTIONS
- Test for the virus: antigen, viral DNA and RNA, subtypes, mutants
- BIBLIOGRAPHY FOR VIRAL INFECTIONS
- Bibliography for viral infections
- VIRAL INFECTIONS
- HERPETIC VIRAL INFECTIONS
- BIBLIOGRAPHY FOR VIRAL INFECTIONS
- BIBLIOGRAPHY FOR VIRAL INFECTIONS
- Viral infections
- RNA VIRAL INFECTIONS
- VIRAL INFECTIONS
- VIRAL INFECTIONS
- BIBLIOGRAPHY FOR VIRAL INFECTIONS
- RNA VIRAL INFECTIONS
- Bibliography for viral infections
- RNA VIRAL INFECTIONS