RNA VIRAL INFECTIONS
Astrovirus Infection
Astroviruses are associated with gastroenteritis in children and a variety of mammals and birds. Astrovirus infection has been documented by PCR and gene sequencing in association with enteritis and mortality in young rabbits in the United States and Italy.
It remains to be determined if astrovirus can be a primary pathogen in rabbits, as it is usually associated with other pathogens of the rabbit enteritis complex.Borna Virus Infection: Borna Disease
Borna virus has a wide host range, and is particularly known to cause meningitis and encephalomyelitis in horses and sheep. Natural infection of rabbits has been documented in Europe, with demonstration of viral antigen in the brain. Experimental infection of laboratory rabbits results in encephalitic lesions with multifocal retinopathy preferentially involving the posterior pole of the eye.
Calicivirus Infections
Caliciviruses are highly significant pathogens of lago- morphs. Pathogenic caliciviruses encompass the Lagovi- rus genus (rabbit hemorrhagic disease virus and European Brown Hare syndrome virus) and the Vesivirus genus (rabbit vesivirus).
Rabbit Hemorrhagic Disease Virus Infection
RHD virus specifically infects Oryctolagus rabbits. Sylvi- lagus rabbits and hares are resistant to RHD virus infection, although European hares are afflicted with a closely related virus, European Brown Hare Syndrome (EBHS) virus. RHD was first recognized among rabbits imported from Germany into China in 1984. Within months, RHD virus killed over 140 million domestic rabbits in China, and subsequently spread to Korea. By the 1990s, RHD was reported in over 40 countries. RHD virus is endemic in wild rabbit populations throughout Europe, and RHD virus was iatrogenically introduced to Australia and New Zealand in the 1990s, where it is now endemic among wild rabbits.
Sporadic outbreaks of RHD, often of unknown origin, have occurred among domestic rabbits in the Americas and northern Africa. RHD virus is believed to have emerged from avirulent endemic viruses circulating subclinically among European rabbit populations. Although rabbits appear to be specifically susceptible to RHD virus, viral RNA has been documented in small mammal species sympatric to wild rabbits.RHD is a reportable disease in the United States. Aggressive depopulation of affected colonies is required. In Europe and other areas of the world where RHD virus is enzootic, vaccination is effective at protecting susceptible populations of rabbits, although new variants of the virus have been shown to cause RHD in vaccinated populations. Various types of vaccines are available, including heat-killed liver extracts, VP60 protein, and a recombinant myxoma-RHD live virus.
Epizootiology and Pathogenesis
RHD virus is highly contagious, and is spread through oro-fecal exposure, as well as environmental contamination and mechanical transmission by blood-feeding insects. RHD virus is highly resistant and stable in the environment. Rabbits that survive RHD may persistently shed virus, and subclinically infected carriers may exist. There are several genogroups of RHD virus, including many nonpathogenic strains that are endemic among wild rabbits. Serologic surveys of RHD-free domestic and wild rabbit populations in Europe have revealed a high frequency of RHD virus-seropositive rabbits, suggesting enzootic infection with nonpathogenic strains of the virus. Contact experiments with seropositive rabbits exposed to nonpathogenic RHD virus have also revealed seroconversion in the absence of clinical signs, whereas exposure of rabbits to pathogenic RHD virus-infected rabbits resulted in clinical disease. It is generally believed that pathogenic RHD virus arose from enzootically circulating nonpathogenic strains of the virus. New pathogenic variants of RHD virus, which are genetically closely related to nonpathogenic strains, have been documented.
For example, an outbreak of disease resembling RHD was reported in Michigan, and the agent was found by sequence analysis to be related to "nonpathogenic" RHD strains.RHD virus binds to carbohydrate moieties of host-cell histo-blood group antigens (HBGAs) that are expressed on the surface of upper respiratory and intestinal epithelium. Young rabbits weakly express one of these HBGAs, which is believed to be a possible mechanism by which rabbits less than 2 months of age are remarkably resistant to infection. Other susceptibility factors are involved, as hepatocytes are a major cellular target for RHD virus, but do not express HBGAs. The age-related resistance of young rabbits is also not absolute. Variant RHD virus strains have been associated with outbreaks in young rabbits on farms in Spain and Portugal where rabbits had been vaccinated, suggesting that these virus strains are antigenically distinct, but also have evolved to target alternate receptors. Furthermore, field strains of RHD virus in Australia have been shown to be more virulent than the original released strain, thereby overcoming host genetic resistance, and there is evidence that HBGA specificities have progressively shifted in virus strains that have evolved in France. Thus, RHD virus poses considerable challenges for control and prevention.
Pathology
The incubation period of RHD ranges between 1 and 3 days, and rabbits are typically moribund within 12-36 hours after onset of fever. Mortality may reach greater than 80% of adult rabbits. In peracute infections, no clinical signs may be apparent. Acute infections are manifest as anorexia, depression, conjunctival congestion, and neurologic signs. Tracheitis, dyspnea, and cyanosis, with foamy hemorrhagic epistaxis and ocular hemorrhage may be present. Subacute infections are milder, with some rabbits surviving, and a small percentage of rabbits may develop chronic disease that features anorexia, lethargy, and jaundice. Chronically infected rabbits usually succumb within 1-3 weeks, but may survive.
At necropsy, there may be blood-stained nasal discharge, pulmonary hemorrhage and edema (Fig. 6.21), hepatomegaly with accentuated lobular pattern (Fig. 6.22), splenomegaly, perirenal hemorrhage, and serosal ecchymoses on areas such as pericardium and intestine. The major histopathologic lesion of RHD is acute necrotizing hepatitis (Fig. 6.23). Viral antigen can be found in hepatocytes within hours of infection. Viral antigen can also be demonstrated in Kupffer cells, macrophages of the spleen, pulmonary macrophages, kidney, and small intestine. Segmental necrotizing enteritis of the small intestine is a primary lesion, but is relatively minor compared to pathology in other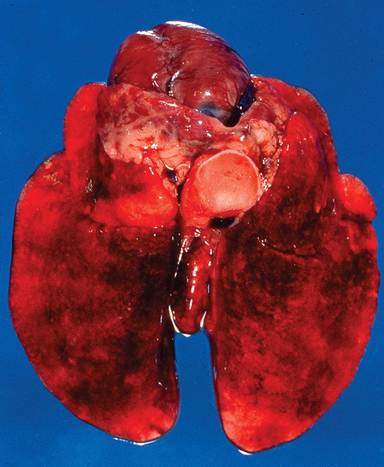
FIG. 6.21. Lungs from a wild Oryctolagus rabbit with acute rabbit hemorrhagic disease (RHD) virus infection. There is marked pulmonary edema with multifocal ecchymoses.
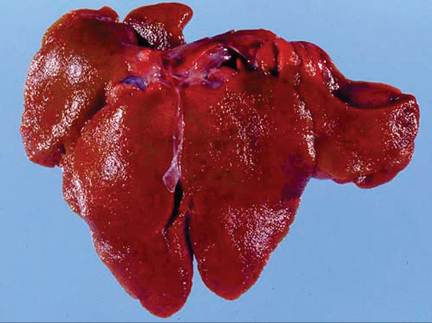
FIG. 6.22. Liver from a domestic rabbit naturally infected with RHD virus. Note the granular texture of the hepatic capsule and accentuated lobular pattern. (Source: Bergin, I.L., et al. 2009. Reproduced from Centers for Disease Control and Prevention, a U.S. Government agency. http://wwwnc.cdc.gov/eid/page/copyright-and- disclaimers)
organs. There is generalized lymphocytic depletion with immunosuppression. The major cause of death in RHD is massive disseminated intravascular coagulation (DIC). Fibrin thrombi are present in small vessels of multiple organs, including kidney, brain, adrenals, heart, testes, and lung (Fig. 6.24).
Diagnosis
Aside from the characteristic clinical signs and lesions, infection can be confirmed by PCR. RHD virus cannot be successfully grown in cell culture. The presence of cal- icivirus within rabbit populations can be evaluated by serology.
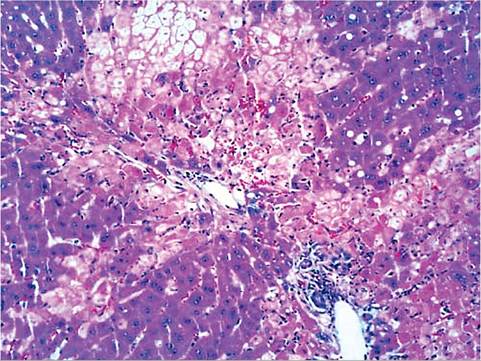
FIG.
6.23. Liver from a rabbit experimentally infected with rabbit hemorrhagic disease virus. Note the acute periportal and midzonal hepatocellular necrosis. (Source: Bergin, I.L., et al. 2009. Reproduced from Centers for Disease Control and Prevention, a U.S. Government agency. http://wwwnc.cdc.gov/eid/page/copyright-and-disclaimers)
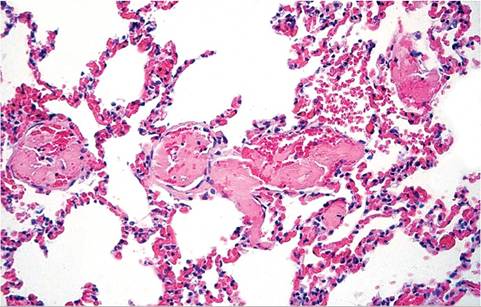
FIG. 6.24. Lung from a case of RHD in a feral Oryctolagus rabbit from
Australia. Note multiple intravascular thromboses (Source: M. Kabay.)
European Brown Hare Syndrome Virus Infection
European Brown Hare Syndrome (EBHS) is caused by a virus that is closely related to RHD virus. The first outbreak of disease occurred in 1980 among hares in Sweden, and is now prevalent throughout Europe. EBHS virus is not infectious to Oryctolagus or Sylvilagus rabbits. The virus has been associated with outbreaks of high morbidity and mortality among farmed hares in Europe, as well as among multiple species of free-ranging hares in Europe. Adult animals experience the highest mortality, and animals less than 40 days of age are unaffected. Disease is characterized as acute necrotizing hepatitis and pulmonary hemorrhages, with DIC, reminiscent of RHD in rabbits. There is no vaccine available for EBHS prevention.
Rabbit Vesivirus Infection
Rabbit vesivirus has been observed in a small Oregon rabbitry. Its etiology was confirmed by electron microscopy and cultivated from the intestine of juvenile domestic rabbits with severe diarrhea and enteritis. Genetic analysis of the virus grouped it within a clade that contained a number of marine caliciviruses and a nonhuman primate calicivirus. As with most cases of rabbit enteritis, other agents including E. coli and coc- cidia likely contributed to disease severity. Oral inoculation of susceptible rabbits with cell culture (Vero cells)- grown virus resulted in subclinical seroconversion.
Coronavirus Infections
Coronaviruses are now divided into 4 genera: Alpha-, Beta-, Gamma-, and Deltacoronavirus.
Based upon serologic evidence and recent genetic sequence analysis, rabbits have been shown to be infected with a member of the Alphacoronavirus genus (pleural effusion disease virus) and Betacoronavirus genus (rabbit enteric coronavirus). The family Coronaviridae also includes the subfamily Toro- virinae. Toroviruses infect a variety of mammals, and have been associated with gastroenteritis in horses and cattle. Neutralizing antibodies against the equine torovirus, Berne virus, have been detected in clinically normal laboratory rabbits. The role of toroviruses in rabbit enteric disease is unknown.Rabbit Enteric Coronavirus Infection
Enteritis has been reported in young rabbits from commercial rabbitries, as well as barrier-maintained laboratory rabbits in Europe and Canada. Serosurveys suggest that coronavirus infection is also present among rabbits in the United States, and recent studies have shown that infection is widespread among rabbits in China. Young rabbits 3-10 weeks of age appear to be most susceptible to enteritis, with mortality reaching 60% of affected rabbits. Experimental infection of susceptible rabbits resulted in transient and mild diarrhea. In natural outbreaks, animals are thin and dehydrated, with fecal staining of the perineal region. The cecum may be distended, containing watery, off-white to tan feces. Microscopic changes are confined to the small and large intestine. Villus blunting, vacuolation, and necrosis of enterocytes, mucosal edema, and polymorphonuclear and mononuclear leukocyte infiltration are characteristic findings. Coronaviral infection is likely to be complicated by other pathogens of the rabbit enteritis complex. The presence of typical coronaviral particles in the feces of young diarrheic rabbits is confirmatory. However, coronaviral particles have also been observed in the gut contents of subclinically infected animals. Although coronaviral enteritis may occur in the absence of identifiable copathogens, careful screening for concurrent infections with other pathogens is recommended.
Pleural Effusion Disease Virus: Coronaviral Cardiomyopathy
Pleural effusion disease and cardiomyopathy have been associated with coronaviral infections in laboratory rabbits in the United States and Europe. In the Scandinavian reports, pleural effusion was recognized in rabbits following inoculation with a coronavirus-contaminated stock of the Nichols strain of Treponema pallidum. The virus was related antigenically to human coronavirus strain 229E. Fatal infections were characterized by lymphoid depletion of the splenic follicles, focal degenerative changes in the thymus and lymph nodes, proliferative changes in glomerular tufts, and uveitis. In rabbits inoculated with material containing the same strain of T. pallidum in the United States, multifocal myocardial degeneration and necrosis were observed. Viral particles were demonstrated in the sera of infected rabbits postinoculation. Antibodies to 2 human strains of coronavirus were demonstrated in convalescent rabbit sera, and antigen was detected in myocardial lesions using antisera to human 229E coronavirus. There is no evidence that the causative agent occurs as a natural pathogen of rabbits.
Hepatitis E Virus Infection
Hepatitis E virus (HEV) is associated with acute hepatitis in humans. Five genotypes, consisting of a single serotype, have been identified in a variety of mammals: genotypes 1 and 2 are restricted to humans, and genotypes 3 and 4 are zoonotic, particularly in swine. HEVs closely related to genotype 3 have been isolated from commercially raised rabbits in China and the United States. The rabbit HEVs were infectious to other rabbits, swine, and macaques. HEV could be detected in serum, tissues and feces of inoculated rabbits, and acute necrotizing hepatitis was found in infected rabbits and macaques. In addition, rabbits are experimentally susceptible to infection with HEV genotypes 1 and 4. Although HEV is not likely to be associated with clinical disease in rabbits under natural conditions, these findings emphasize the potential zoonotic risk of HEV-infection in rabbits.
Paramyxovirus Infection
During the mid-1980s, a survey of laboratory rabbits in Japan revealed seroconversion to Sendai virus in over 50% of rabbits from a number of different colonies. Laboratory rabbits are susceptible to experimental Sendai virus infection, although viral replication appeared to be confined to the upper respiratory tract. Following intranasal inoculation with the MN strain of Sendai virus, there was transient viral shedding postinoculation, and animals seroconverted. Viral antigen was detected in the nasal epithelium for up to 10 days. Infected rabbits remained clinically normal throughout the study. Sendai virus is now rare or nonexistent among laboratory animals, but other paramyxoviruses may arise in the future. Acute fatal pneumonia was reported in a single dwarf rabbit from a small colony in Europe in which a paramyxovirus was incriminated, based upon ultrastructural findings. The rabbit developed acute interstitial pneumonia in which alveoli were filled with exfoliated lining cells, macrophages, and cellular debris. Rare multinucleate syncytia and eosinophilic inclusions were observed among detached alveolar lining cells.
Rabies Virus Infection
Rabies has been documented in several domestic rabbits in the United States, and both raccoon and skunk variants have been documented in infected rabbits. These cases serve to emphasize the importance of adequate protection for any pet animals with access to an outdoor environment.
Rotavirus Infection
Rotaviruses are divided into 5 species: A, B, C, D, and E. Group A rotaviruses are the most significant pathogens for both humans and animals, including rabbits. Within each rotavirus species, rotaviruses are classified based upon nucleotide sequences of 2 outer capsid proteins, VP7 (glycoprotein) and VP4 (protease-sensitive), which define the “G” and “P” genotypes. Rabbits rotaviruses are typically G3P[14] and G3P[22], although a bovine-like G6P[11] genotype rotavirus was recently found in a laboratory rabbit colony, but rabbits had no clinical signs of disease. Rotaviruses have been associated with diarrheal disease in domestic rabbits in Europe, Japan, and the United States.
Epizootiology and Pathogenesis
Rotavirus typically causes diarrhea among infant rabbits, since virus targets terminally differentiated enterocytes lining the tips of villi of the jejunum and ileum. Since neonatal bowels have a high number of terminally differentiated enterocytes with slow epithelial turnover kinetics, young rabbits are most susceptible to clinical disease. However, maternal antibody, if present, is protective at this early age. Serosurveys have confirmed that rotaviral infections are commonly endemic within domestic Oryc- tolagus rabbits, as well as among wild Sylvilagus and Lepus lagomorphs in Europe, Asia, and North America. Epidemiologic studies have indicated that protection may be afforded by transplacentally derived maternal antibodies, which subsequently decline to low levels in kits by 1 month of age. However, on exposure to the virus at 30-45 days of age, there may be sufficient residual antibody to protect kits from overt disease. Based on serological surveys, clinically healthy rabbits may have a subclinical infection at around 4 weeks, with subsequent rise in antibodies to rotavirus. In 1 epizootic of rotaviral enteritis in a specific-pathogen-free rabbitry, sucklings 1-3 weeks of age were affected. The disease was characterized by rapid spread, with high morbidity and mortality. This was consistent with the introduction of virus into a colony not previously exposed, thus with no maternal immunity to afford protection during the neonatal period. Virus shedding in diarrheic rabbits is frequently seen at 35-42 days of age. In addition to relatively mild lytic effects of virus infection, which may not be microscopically apparent, virus NSP4 induces a number of physiologic effects upon enterocytes, including chloride secretion, disruption of water reabsorption, disruption of disaccharidases, and activation of secretory reflexes of the enteric nervous system. Due to perturbation of disaccharidases, lactose and other disaccharides remain in the gut lumen, causing an osmotic drain and attracting fluid into the lumen. Viral infections of enterocytes may also facilitate bacterial adhesion to damaged cells. In weanling rabbits inoculated with an enteropathogenic strain of E. coli, superimposed rota- viral infections have been shown to have an additive effect, resulting in increased morbidity and mortality.
Pathology
On gross examination, animals may be dehydrated, and fluid contents are present in the cecum. Other organs are usually grossly normal. In the small intestine, lesions are generally mild, although there may be moderate to severe villus blunting, villus fusion, and vacuolation to flattening of apical enterocytes in the jejunum and
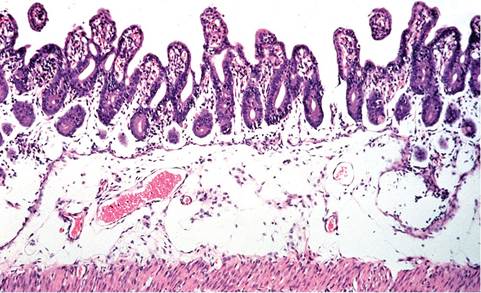
FIG. 6.25. Small intestine from a juvenile rabbit with a natural case of epizootic rotaviral enteritis. There is submucosal edema, with blunting and fusion of villi. Enterocytes within crypts and on villi are immature, indicative of regeneration. (Source: Schoeb et al. 1986. Reproduced with permission from American Association for Laboratory Animal Science.)
ileum, variable epithelial exfoliation, and edema of the submucosa (Fig. 6.25). There may be focal areas of exfoliation of surface epithelium in the cecum.
Diagnosis
Demonstration of the virus (usually by examination of intestinal contents for viral particles by electron microscopy), virus isolation, and serology enable incrimination of rotavirus as the causative viral agent. ELISA kits, marketed for detection of human rotavirus antigen in feces, are available from commercial suppliers and work well for testing fecal samples for group A rotaviral antigen. Fecal flotation and/or microscopic examination of the small and large intestines are essential steps to determine whether coccidia or copathogenic bacteria are playing a role in the disease. Differential diagnoses include coronaviral enteritis, coliform enteritis, salmonellosis, clostridial enteropathies, and coccidiosis. Demonstration of rotaviral particles or antigen in the feces does not constitute a definitive diagnosis but should be confirmed by microscopic examination of the gastrointestinal tract for characteristic lesions.
Endogenized Viruses
Genomic sequencing has demonstrated a number of viruses that have been integrated (endogenized) into the genome of lagomorphs. The best known of these viruses are endogenous retroviruses. Although rabbit endogenous retrovirus has been shown to be expressed in lymphoma cell cultures, its role as an etiologic agent of lymphoma has not been demonstrated. Notably, a rabbit endogenous retrovirus DNA has been found to contaminate human DNA samples from a number of sources, and was until recently believed to be a human retrovirus known as HRV-5. As with other placental mammals, lagomorphs have “domesticated” the envelope sequence of an ancient retrovirus, known as syncytin (syncytin-Ory1). Rabbit syncytin is genetically distinct from syncytin genes of rodents and primates, suggesting a separate origin but similar evolutionary advantage. Syncytins are selectively expressed in the placenta, and are essential for syncytiotrophoblast formation. Perhaps the most interesting rabbit endogen- ized retrovirus is rabbit endogenous lentivirus type K (RELIK). RELIK is defective, but it represents evidence of the ancient origins of lentiviruses, and is the first lenti- virus to be shown that integrated into a mammalian genome. Endogenized DNA parvovirus-like elements have also been found in the rabbit genome. These virus-like genetic sequences are incomplete (defective) and not replication-competent.
More on the topic RNA VIRAL INFECTIONS:
- RNA VIRAL INFECTIONS
- RNA VIRAL INFECTIONS
- RNA VIRAL INFECTIONS
- Test for the virus: antigen, viral DNA and RNA, subtypes, mutants
- HERPETIC VIRAL INFECTIONS
- BIBLIOGRAPHY FOR VIRAL INFECTIONS
- Bibliography for viral infections
- VIRAL INFECTIONS
- BIBLIOGRAPHY FOR VIRAL INFECTIONS
- Viral infections
- BIBLIOGRAPHY FOR VIRAL INFECTIONS
- DNA VIRAL INFECTIONS