DNA VIRAL INFECTIONS
Adenovirus Infections
Mice are host to 2 distinct adenoviruses, murine adenovirus-1 (MAdV-1) and murine adenovirus-2 (MAdV-2), which should be more accurately termed mouse adenovirus 1 and 2.
MAdV-1 and MAV-2 can be differentiated from each other genetically, serologically, and by pathology. MAdV-1 and MAdV-2 differ significantly at the DNA level, with MAdV-2 having a distinctly larger genome. Adenoviruses are nonenveloped DNA viruses that replicate in the nucleus. Infection results in pathognomonic intranuclear inclusions.Epizootiology and Pathogenesis
MAdV-1 (previously FL) was initially discovered as a contaminating cytopathic agent during attempts to establish Friend leukemia (FL) virus in tissue culture. Infection is transmitted by direct contact through urine, feces, and nasal secretions. Serologic surveys have indicated that MAdV-1 was at one time common in laboratory mouse colonies, but it is now rare or nonexistent in laboratory mice from North America and Europe. Naturally occurring clinical disease or lesions due to MAdV-1 have not been described, but experimental intraperitoneal inoculation of neonatal, suckling, and immunodeficient mice with MAdV-1 results in viremia and a fatal, multisystemic infection within 10 days. MAdV-1 infects cells of the monocyte-macrophage lineage, microvascular endothelial cells, respiratory epithelium, adrenal cortical cells, and renal distal tubular cells. Experimental inoculation of weanling or adult mice results in multisystemic infection with leukocyte-associated viremia and prolonged viruria. There are marked mouse strain differences in susceptibility to experimental infection. Mice less than 3 weeks of age are universally susceptible to experimental disease, and adult C57BL/6, DBA/2, SJL, SWR, and outbred CD-1 mice tend to be susceptible to lethal experimental disease, whereas BALB/c, C3H/HeJ, and most other inbred strains tested are disease resistant.
Susceptible strains develop hemorrhagic encephalitis, which does not occur in resistant strains. The nature of infection of immunodeficient strains of mice depends upon strain genetic background. BALB-scid and BALB-scid/beige mice develop fatal disseminated infection with focal hemorrhagic enteritis and microvesicular fatty change in the liver, consistent with Reye's-like syndrome, but no neurologic involvement. B6-Rag1 mice develop disseminated disease with hemorrhagic encephalomyelitis.Athymic C3H/HeN-nude mice develop a progressive wasting disease with disseminated infection and duodenal hemorrhage, but no central nervous system involvement. Thus, genetic background is a major determinant of susceptibility to lesions in the central nervous system. Lymphoid B cells are critical for controlling disseminated infection, and T cells are required for recovery from infection but also contribute to pathology.
MAdV-2 (K87) was initially isolated from feces of an otherwise healthy mouse. In contrast to MAdV-1, MAdV- 2 is principally enterotropic, regardless of route of inoculation, mouse strain, or immunosufficiency. Following oral inoculation of mice less than 4 weeks of age, MAdV-2 is excreted in feces for 3 or more weeks with peak infection between 7 and 14 days. Immunocompetent mice apparently recover. Seroconversion of rats to MAdV-2 has been noted, but they are not susceptible to experimental inoculation with the mouse virus, suggesting that they are host to a related, but different, adenovirus (see Rat Chapter 2, Rat Adenovirus Infection).
Pathology
Natural infection of immmunocompetent adult mice with MAdV-1 is typically subclinical, but the growing number and use of immunodeficient strains of mice warrant awareness of experimental findings. Mice experimentally inoculated with MAdV-1 develop runting, dehydration, thymic involution, and grossly evident foci of necrosis in liver, spleen, and other organs. Intranuclear inclusions can be found within foci of necrosis and hemorrhage in multiple organs, including brown fat, myocardium, cardiac valves, adrenal gland (Fig.
1.16), spleen, brain, pancreas, liver, intestine,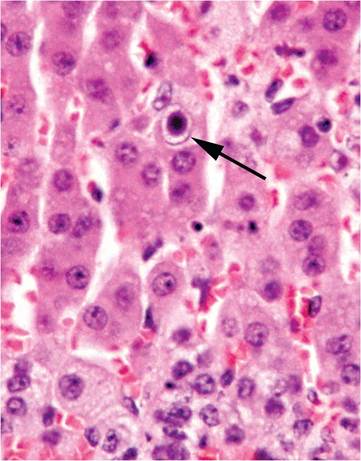
FIG. 1.16. Adrenal cortex from a mouse experimentally infected with mouse adenovirus MAdV-1. A single intranuclear inclusion (arrow) is present in the cortical epithelium.
salivary glands, and renal distal tubular epithelium. Peribronchiolar and pulmonary interstitial infiltration with mononuclear leukocytes have been found in adult B6 mice following intranasal inoculation. Focal hemorrhagic enteritis has also been noted in some genotypes of mice following experimental inoculation. Gastrointestinal tracts can be empty, with segmental inflammation and hemorrhage in the distal duodenum and jejunum. Inclusions in intestinal lesions tend to be difficult to visualize. Hemorrhagic foci may occur throughout the central nervous system but especially in the white matter of susceptible mouse strains. Endothelial cell necrosis is prominent in these foci, but inclusions are rare except in Purkinje cells. Central nervous system lesions can be clinically manifest as rigid tails, hypermetria, paraphimosis, ataxia, and urinary bladder distention.
Clinical signs are usually absent in MAdV-2 infected mice, but runting of suckling mice has been observed in natural infections. Naturally infected athymic nude mice are clinically normal. Gross lesions of MAdV-2 infection are not evident, except that juvenile mice may be bloated and runted. Mice naturally or experimentally infected with MAdV-2 develop intranuclear inclusions in mucosal epithelial cells of the small intestine, especially in the distal segments, and the cecum. Inclusions are most plentiful in infant mice but can also be found in smaller numbers in the mucosa of adult mice, especially dams that are suckling infected infants. Similar inclusions have been noted in adult nude mice without other detectable lesions. Typically, inclusionbearing nuclei are located in the apical portions of cells, rather than in their normal basal location (Fig.
1.17).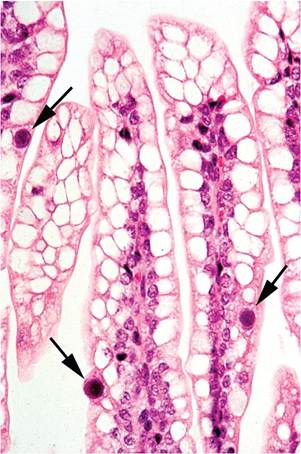
FIG. 1.17. Small intestine from a juvenile mouse infected with mouse adenovirus MAdV-2. Note the distinct intranuclear inclusion bodies (arrows) in a few enterocytes lining the villi.
Diagnosis
Serology is the most effective means of screening mouse populations for MAdV infection. These agents partially cross-react serologically, depending on method, in a one-way relationship with antiserum to MAdV-2 crossreacting with MAdV-1, but not conversely. Since MAdV- 1 is quite rare in laboratory mice, MAdV-2 antigen should be used. Differential diagnoses for MAdV-1 include multisystemic infections that produce intranuclear inclusions, such as polyoma virus and cytomegalovirus. Intestinal epithelial MAdV-2 inclusions are pathognomonic, but not always apparent. They are not induced by any other known agent, although K virus can form inclusions in intestinal endothelial cells. Inclusions in the apical region of enterocytes must be differentiated from mitotic cells and intraepithelial lymphocytes. Success at finding MAdV-2 inclusions is maximized in infant mice. MAdV strain-specific PCR can also be used.
Herpesvirus Infections
Mice are host to two members of the family Herpesvir- idae, subfamily Betaherpesvirinae and genus Muromega- lovirus, including mouse cytomegalovirus (MCMV) and mouse thymic virus (MTV). Neither is common among contemporary laboratory mouse populations, but may contaminate archival biological products.
Mouse Cytomegalovirus Infection
MCMV is a mouse-specific virus that was originally isolated by M.G. Smith from salivary glands of naturally infected laboratory mice. Cytomegaloviruses replicate in the nucleus and cause cytomegalic inclusion disease, characterized by enlarged cells bearing both intranuclear and intracytoplasmic inclusions, particularly in salivary glands. MCMV has been studied extensively as an animal model of human CMV (HCMV) infection, but significant biological differences exist between MCMV and HCMV.
Most laboratory studies have utilized the original Smith strain of MCMV, or derivatives thereof, and may not accurately reflect the natural biology of other MCMV strains. Nevertheless, the intense scrutiny of MCMV as a prototype model system has shed considerable light on its pathogenesis.Epizootiology and Pathogenesis
Wild mice are commonly infected with MCMV. There are multiple genetically diverse MCMV strains within wild mouse populations, and mixed infections of single mice are common. Laboratory studies have found that infection-induced immunity to one strain does not preclude infection with a second strain. MCMV is transmitted oronasally by direct contact and is excreted in saliva, tears, urine, and semen. Experimental infection is significantly influenced by virus strain, dose, route of inoculation, and host factors (age, genotype). Neonates of all mouse strains are universally susceptible to severe disease, and resistance to lethal disease evolves after weaning and increases until about 8 weeks of age. Genetically resistant mouse strains include B6, B10, CBA, and C3H mice, and susceptible strains include BALB/c and A strain mice. Resistance is associated with H-2k haplotype, but non-H-2 associated factors also exist, including a resistance factor that is linked to loci on chromosome 6 within the natural killer (NK) cell complex. This region encodes a receptor expressed on NK cells that binds to a glycoprotein of MCMV, but MCMV isolates from wild mice have naturally occurring mutations that fail to activate NK cells through this receptor. NK cells interact with MCMV in other ways that are linked to the H-2k haplotype.
Following experimental inoculation of infant mice, viremia and multisystemic dissemination occur within 1 week, with infection of lung, heart, liver, spleen, salivary glands, and gonads. Macrophages are major targets of the virus, and blood monocytes are important for the viremic phase of infection. Following dissemination, virus is rapidly cleared from tissues, except from salivary gland.
Infection of NK-deficient beige mice, or mice depleted of NK cells, significantly prevents virus clearance. Adaptive immune responses, including CD4 and CD8 cells, are also important in clearing infection from most sites. Athymic or SCID mice fail to control active infection, but B-cell-deficient mice can recover from acute infection. Curiously, despite functional innate and acquired immune responses in fully immunocompetent mice, including NK, CD4, CD8, and B cells, MCMV continues to persist and replicate in salivary gland tissues. A number of MCMV genes function to control the innate and acquired immune responses, in addition to determining cell tropism and inhibiting apoptosis. Most of these genes are not essential for virus replication in vitro, and therefore provide selective advantages for virus persistence in vivo. An important feature of MCMV (and other herpesviruses) is latent infection, in which virus persists in a nonreplicative state, but can be reactivated by immunosuppression or stress. Based upon tissue explanting and PCR, latency of MCMV has been documented in various organs, including salivary glands, lung, spleen, liver, heart, kidney, adrenal glands, and myeloid cells. This state of latency can persist for the life of the mouse.Unlike HCMV, MCMV does not readily cross the placenta, and in utero transmission does not usually take place in naturally and experimentally infected mice. Infection of pregnant mice may cause fetal death and resorption, delayed birth, and runted pups, but these are nonspecific events. Nevertheless, latently infected dams have been documented to transmit low- level or latent infection to fetuses in utero. MCMV infects cells in the epididymis, seminal vesicles, testes, including Leydig cells and spermatozoa, as well as ovarian stromal cells. Experimental transmission by artificial insemination has been reported. Thus, sexual transmission is likely.
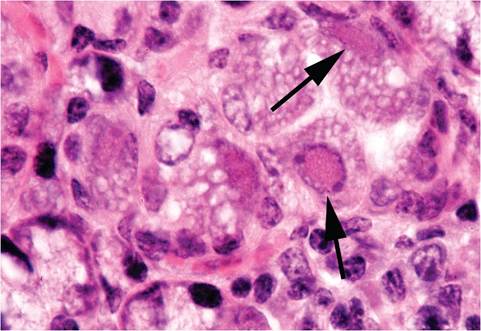
FIG. 1.18. Submaxillary salivary gland from a wild mouse infected with mouse cytomegalovirus. Intranuclear inclusions (arrows) are present in acinar epithelial cells.
Pathology
Overt disease and disseminated lesions do not usually occur in naturally infected mice. The most frequently encountered lesions occur in the submandibular salivary glands and, rarely, in the parotid glands. Eosinophilic intranuclear and intracytoplasmic inclusions are present in acinar epithelial cells with cytomegaly (Fig. 1.18) and lymphoplasmacytic infiltration of interstitium. During the acute disseminated phase in experimentally inoculated infant mice or T-cell-deficient mice, focal necrosis, cytomegaly, inclusions, and inflammation occur in many tissues, including salivary glands, lacrimal glands, brain, liver, spleen, thymus, lymph nodes, peritoneum, lung, skin, kidney, bowel, pancreas, adrenal, skeletal and cardiac muscle, cartilage, and brown fat. Diffuse interstitial pneumonitis has been described in BALB/c mice that were immunosuppressed by a variety of methods, and athymic mice develop progressive multifocal nodular pulmonary inflammation. Athymic mice also develop progressive destruction of adrenals. Arteritis of the pulmonary artery and aorta (at the base of the heart) has been documented during experimental MCMV infection in B6 and BALB/c mice, but no virus was confirmed within the lesions. A single case of naturally occurring MCMV disseminated infection has been reported in an aged laboratory mouse. MCMV has been shown to have a synergistic effect with Pseudomonas aeruginosa.
Diagnosis
Lesions in salivary glands are typical of cytomegalovirus, but are not always present in infected animals. Serology is the method of choice for colony surveillance, but detection of infection in immunodeficient mice that do not seroconvert must be accomplished through nucleic acid detection, including in situ hybridization and PCR. These methods can also be used to detect latent infection. Differential diagnosis for sialoadenitis with inclusion bodies must include polyoma virus. Other viruses that infect salivary glands include reovirus 3, mouse thymic virus, and mammary tumor virus, but these viruses do not induce inclusions.
Mouse Thymic Virus Infection
Detailed information about mouse thymic virus (MTV) is generally lacking, since in vitro methods of propagation have not been identified and little experimental work has been performed. Virtually nothing is known about its genome. Synonyms include thymic necrosis virus, thymic agent, and mouse T-lymphotropic virus. The prevalence of MTV in laboratory mouse populations is extremely rare, but it is very common among wild mice. MTV can be a contaminant of MCMV stocks prepared from salivary glands. Because of its lymphocytotropism, MTV infection may result in significantly altered immune responses.
Epizootiology and Pathogenesis
MTV was first discovered when inoculated newborn Swiss mice developed thymic necrosis following serial passage of mammary tumor homogenates. This feature of MTV has been emphasized in subsequent studies, but MTV infects salivary glands as its primary target. Outcome of experimental infection is strikingly age dependent and also influenced by mouse genotype. Intraperitoneal as well as oronasal inoculation of newborn mice results in acute thymic necrosis, which is visible grossly as diminished thymic mass, within 1-2 weeks. Subsequently, the thymus returns to normal, but mice remain persistently infected. Age-related susceptibility to thymic necrosis decreases progressively until 6 days of age, at which point mice are no longer susceptible to thymic necrosis. Although largely an experimental phenomenon, thymic necrosis has been encountered in infant mice from naturally infected mouse colonies. CD4+8+ and CD4+8- T cells are selectively targeted by MTV, although virus replication also occurs in thymic epithelial cells and macrophages. Newborn mice develop viremia, with MTV detectable in multiple organs. Mice of all ages develop infection of salivary glands, with persistent virus shedding in saliva for several months or more. Athymic mice, which lack the T-cell substrate for virus replication, tend to shed virus less consistently. The mode of MTV transmission is presumed to be primarily through the saliva, and MTV is readily transmitted by direct contact. MTV has also been isolated from mammary tissue of an infected mother and from mammary tumor extracts, suggesting another possible route of transmission. Vertical (in utero) transmission has not been documented.
Pathology
MTV infection of infant mice results in the formation of intranuclear inclusion bodies and necrosis of thymocytes and, to a lesser extent, cells in lymph nodes and spleens. During recovery, there is granuloma formation. Lesions in salivary glands have not been noted. BALB/c and A strain mice, but not B6, C3H, or DBA/2 mice, when inoculated as neonates develop gastritis. Mice of other strains develop oophoritis and antibodies to thyroglobulin. These phenomena are believed to be autoimmune in origin, via nonspecific activation and expansion of self-reactive T cells, and not related to MTV in these tissues.
Diagnosis
Seroconversion can be detected using infected salivary tissue as antigen. Mice infected as neonates may not seroconvert, probably due to immune tolerance. The mouse antibody production (MAP) test can be useful, but PCR is now available for testing mouse tissue and biologic products. Differential diagnoses include coronavirus or stress, which may cause thymic necrosis, but not inclusions. A bioassay has been used to detect MTV, in which inoculated infant mice develop thymic necrosis.
Parvovirus Infections
Laboratory mice are subject to infection with two different autonomously replicating types of viruses in the family Parvoviridae: minute virus of mice (MVM) and mouse parvovirus (MPV). The official and generally unaccepted name for MVM is mice minute virus (MMV), which will not be used in this text. MVM and MPV, including dual infections, are among the most prevalent viruses in contemporary laboratory mouse populations. MPV strains represent the predominant type (75%) in parvovirus-positive populations. Clinical disease is seldom present in immunocompetent mice, but these viruses have significant immunomodulatory effects, and they are remarkably refractory to effective eradication from contaminated mouse colonies.
MVM and MPV share considerable homology among genes that encode 2 antigenically cross-reactive (among all mouse parvoviruses) nonstructural proteins, NS1 and NS2, but display variation among genes that encode structural capsid proteins, VP1, VP2, and VP3. VP2, in particular, contributes to significant antigenic and biologic differences among mouse parvoviruses. Sequence analysis, differential PCR, and restriction fragment length polymorphism analysis have lent clarity to the rodent parvovirus interrelationships. The MVM group contains MVMp, MVMi, MVMm, and MVMc, and the MPV group contains a cluster of closely related MPV-1a, MPV-1b, MPV-1c, a somewhat disparate cluster containing MPV-2, and another cluster containing MPV-3 and a closely related hamster parvovirus that is closely related to MPV-3 and likely to be of mouse origin. The mouse parvoviruses are distinctly different from the parvoviruses of rats. More isolates and strains are likely to be discovered, and therefore it is most expedient to discuss the two major phylogenetic groups.
Epizootiology and Pathogenesis
Mouse parvoviruses are transmitted through feces and urine by oronasal exposure with a slow rate of cage-to- cage spread. In general, parvoviruses of all host species are dependent upon the S phase of the cell cycle for virus replication, and therefore induce cytolytic disease only in dividing tissues (including lymphoid tissues undergoing antigenic stimulation). However, virus replication, and therefore patterns of disease, is limited to certain cell types that bear the appropriate viral receptors. For example, many parvoviruses replicate in intestinal crypt epithelium, but rodent parvoviruses do not target that cell population, and therefore do not induce intestinal disease.
Following oral inoculation, mouse parvoviruses initially replicate in intraepithelial lymphocytes, lamina propria, and endothelium of the small intestine, and then disseminate to multiple organs, including kidney, intestine, lymphoid tissues, liver, and, to a much lesser extent, lung, with tropism for endothelial cells, hematopoietic cells, and lymphoreticular cells. Viremic dissemination is likely related to the high degree of lymphocytotropism of these viruses, but MVM viremia is also erythrocyte associated. Both types of virus target small intestine and lymphoid tissue, and MVM also replicates in kidney. MVM infection of both infant and adult immunocompetent mice is limited in duration, with recovery. In contrast, MPV infection is typically persistent following infection of mice of all ages, but juvenile mice may transmit virus more efficiently. Neonatal mice are protected from infection by maternal antibody in enzootically infected colonies. Mice resist reinfection with the homotypic virus, but are fully susceptible to infection with the heterotypic serotype, thus explaining the naturally high frequency of dual MVM and MPV infections.
Pathology
Natural infection of immunocompetent mice is clinically silent, regardless of age or strain of mice. Experimental infection of neonatal BALB/c, SWR, SJL, CBA, and C3H mice with MVMi has been shown to cause mortality due to hemorrhage, hematopoietic involution, and renal papillary infarction. DBA/2 neonates also developed intestinal hemorrhage and more rapid hepatic hematopoietic involution, whereas B6 neonates are resistant to vascular and hematopoietic diseases. MVM is more pathogenic for hematopoietic tissues than MPV. Infection of neonatal BALB/c mice with MVMi revealed replication and a significant decrease in bone marrow and splenic cellularity, with depressed myelopoiesis. MVM infection of SCID and neonatal mice has been found to induce lethal leukopenia, due to virus replication in primordial hematopoietic cells, with severe depletion of granulomacrophagic cells and compensatory erythropoiesis in bone marrow. Natural MVM-related disease has been observed in
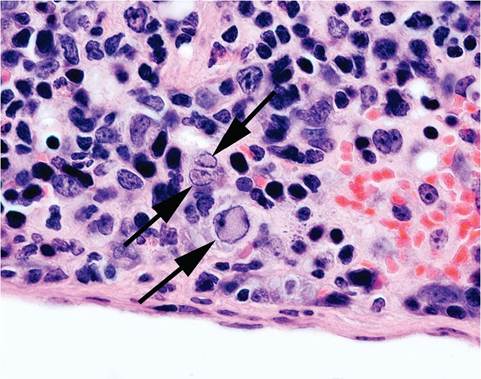
FIG. 1.19. Intranuclear inclusions (arrows) in mononuclear cells of the spleen in an immunodeficient mouse naturally infected with minute virus of mice (MVM). (Source: Franklin 2006. Reproduced with permission from Oxford University Press.)
NOD.Cg-H2H4-Igh-6 null mice with leukopenia and anemia. Intranuclear inclusions were present in mononuclear cells of spleens (Fig. 1.19) and bone marrow. Experimental exposure of infant mice with MVM, including intranasal inoculation and contact exposure, results in virus replication in the cells of the subventric- ular zone, subependymal zone of the olfactory bulb, and the dentate gyrus of the hippocampus. These are the 3 main germinal centers in postpartum neurogenesis of the mouse. MVM also targets the outer granular layer of the cerebellum, with cytolysis of cells that have migrated to the internal granular layer, resulting in granuloprival cerebellar hypoplasia. In experimentally infected pregnant mice, MVM replication may occur in various tissues, including placenta and fetus, without histological evidence of lesions.
Diagnosis
MVM and MPV infection is usually diagnosed by seroconversion, but colony surveillance can be challenging due to the inefficient cage-to-cage transmission of these viruses, thereby requiring large sample sizes for surveillance. Indeed, the inefficient cage-to-cage transmission has made it possible to test and cull as a means of eliminating infection from a population. Furthermore, experimental inoculation of adult ICR, BALB/c, C3H/ HeN, C57BL/6, and DBA/2 mice with different doses of MPV has revealed marked differences in serologic response, with antibody detection in all C3H mice, some but not all ICR, BALB, and DBA mice, and none of the B6 mice, except when inoculated with very high doses of virus. ELISAs utilize the extensive cross-reactive recombinant NS1 antigen to detect antibody to both MVM and MPV, and recombinant VP2 antigen or empty capsids can be used to differentiate antibodies between
MVM and MPV. Caution is advised, since antibodies to new isolates or strains that are more distantly related to MVM or MPV may not be detected. PCR primers for detection of conserved and group-specific regions of the genome are available for detection of virus in tissues and feces. The mesenteric lymph node is an optimal tissue for PCR-based detection of mouse parvoviruses. A comparison of sensitivity of testing procedures resulted in the following percentage of positives: mesenteric lymph node PCR (93%), serologic immunofluorescence assay (68%), direct fecal PCR (10%), and cage fecal PCR (5%). The MAP test and virus isolation, including tissue explant cultures, are more labor-intensive methods that can also be utilized.
Papillomavirus Infections
A papillomavirus (MusPV) was discovered in an inbred NMRImude mouse colony, in which the mice developed florid papillomas at the mucocutaneous junctions of their nose and mouth. The agent induced papillomas in T-cell-deficient (athymic nude and SCID) strains of mice. MusPV is infectious, but not oncogenic, in a variety of immunocompetent strains of mice, which display varying degrees of susceptibility. MusPV is genetically closely related to the rat papillomavirus virus. Another M. musculus papillomavirus has since been reported from normal ear skin tissue of wild mice in Europe. Although these viruses are rare in laboratory mouse populations, they are likely to attain popularity as research models, with subsequent iatrogenic introduction to laboratory mouse populations.
Polyomavirus Infections
Mice are hosts to two genetically distinct polyomaviruses: polyoma virus (PyV) and K virus. They belong to the family Polyomaviridae, which contains a number of related viruses (macaque SV-40 virus, human BK and JC viruses, hamster polyomavirus, rat polyomavirus, and rabbit kidney vaculolating virus). These viruses once belonged to the family Papovaviridae, but that family has been disbanded and split into Polyomaviridae and Papillomaviridae. PyV encodes a middle T antigen that is important in oncogenesis. The K virus genome is similar to that of PyV, but lacks the middle T antigen of PyV. Both viruses are rare or nonexistent in contemporary laboratory mouse populations, but PyV continues to be used as an experimental oncogenic virus that on occasion results in iatrogenic introductions to susceptible mouse colonies.
Polyoma Virus Infection
PyV was originally termed the “Stewart-Eddy (SE) polyoma virus” and the “parotid tumor virus.” The virus has been extensively studied as an oncogenic virus that induces many (poly) types of tumor (-oma). The name is well deserved, as tumors arise from more than a dozen different cell types. Under experimental conditions, it is oncogenic in several different species. The oncogenic activity for which this virus is so well known is largely an experimental phenomenon, requiring parenteral inoculation of genetically susceptible strains of mice within the first 24 hours of life with high titers of selected isolates virus with high oncogenic activity. The relevance of PyV to the laboratory mouse has risen with the use of PyV middle T (PyV-MT) gene as a component of transgenic constructs. PyV is known to contaminate transplantable tumors and cell lines, which in turn have served as inadvertent sources of contamination of mouse stocks.
Epizootiology and Pathogenesis
PyV was initially discovered by Ludwig Gross, when newborn mice unexpectedly developed salivary gland tumors following inoculation with filtered extracts of mouse leukemia tissue. PyV is an environmentally stable virus that is shed primarily in saliva, urine, and feces. Infection is most efficiently acquired intranasally. Infection of a mouse population requires a continuous source of exposure, which is provided by repeatedly utilized nesting sites of wild mice. The virus fails to survive under laboratory mouse husbandry conditions and is, therefore, quite rare in contemporary mouse colonies. Oro- nasal inoculation of neonatal mice results in virus replication in the nasal mucosa, submaxillary salivary glands, and lungs, followed by viremic dissemination to multiple organs, including kidneys. Mortality can be high at this stage. By day 12, the virus is cleared by the host immune response from most sites but persists in the lung and especially kidney for months, where virus replicates in renal tubular epithelial cells. Infection of older mice is more rapidly cleared, with inefficient virus excretion for shorter periods. Transplacental transmission does not seem to occur naturally, but virus can be reactivated in the kidneys of adult mice during pregnancy if they were infected as neonates. Since PyV is a widely used experimental virus, iatrogenic contamination of laboratory mice can take place, but consequences are limited for the reasons just cited. Thus, under natural conditions, maternal antibody from immune dams, coupled with the low level of environmental contamination in a laboratory mouse facility, precludes successful infection of neonatal mice and attenuates survival of the virus in the population.
The oncogenic characteristics of PyV have been extensively studied and are herein reviewed because pathologists may encounter mice that are either experimentally infected with the virus, inadvertently exposed to the virus as immunodeficient strains, or derived from PyV transgenic constructs. Not all strains or isolates of the virus are oncogenic, including many “wild-type” isolates. Virus strains that produce “large plaques” in cell culture can induce tumors in 100% of susceptible mice, whereas other strains, which produce “small plaques,” induce few or no tumors. If genetically susceptible mice are experimentally inoculated parenterally with oncogenic virus at less than 24 hours of age, microscopic foci of cellular transformation arise in multiple tissues. Most of these foci remain microscopic, but others grow rapidly into large tumors within 3 months. Tumors arising in mice inoculated with less oncogenic strains may not arise until 6-12 months, and the tumors that arise are usually mesenchymal, rather than epithelial in origin.
The genetic basis of susceptibility to PyV oncogenesis has been extensively studied in more than 40 inbred strains and F1 hybrids, with susceptibility ranging from 100% to complete resistance, with many intermediate phenotypes. Resistance can be determined by both immunological and nonimmunological factors. For example, C57BL mice are highly resistant due to effective antiviral and antitumor immunity, which can be abrogated by neonatal thymectomy, irradiation, or immunosuppression. C57BR mice are susceptible to infection as neonates but do not develop tumors. Other strains of mice are resistant to tumor induction, even when immunosuppressed. Susceptibility to tumor induction has been linked to H-2k haplotypes. C3H/ BiDa (H-2k) are fully susceptible, whereas DBA/2 and BALB/c mice (H-2d) are resistant. In addition, susceptibility is conferred in many mouse strains by an endogenous mouse mammary tumor provirus, Mtv-7. Mtv-7 encodes a superantigen (Sag) that, when expressed, results in deletion of Vβ6+ T cells. This abrogates the ability of these mice to mount an effective antitumor cytotoxic T-cell response. Another, non-Sag mechanism for susceptibility has recently been discovered in wild- derived inbred mice.
The biological characteristics of PyV are uniquely suited for disseminated, polytropic infection and neoplasia. The virus protein VP1 nonselectively binds universally to sialic acid of cell surfaces, contributing to polytropism. The virus also possesses multivalent enhancer regions that enable it to be transcribed and replicate in many cell types. The virus encodes three T (tumor) antigens that interact with various cell factors and growth signaling pathways. PyV-MT antigen is the major transforming protein that binds and activates protein kinase pp60 (c-Src) and other members of the c-Src family. PyV-MT antigen can transform cells by itself and is therefore a popular component of transgenic constructs for neoplasia research. Many transgenic lines of mice possess PyV-MT contributions to their genome.
Pathology
Under natural conditions, lesions are not likely to be encountered, except in immunodeficient mice. Nude mice have been shown to develop multifocal necrosis and inflammation, followed by tumor formation in multiple tissues reminiscent of experimentally inoculated neonatal mice. Microscopic examination of tissues from neonatally inoculated mice has revealed foci of
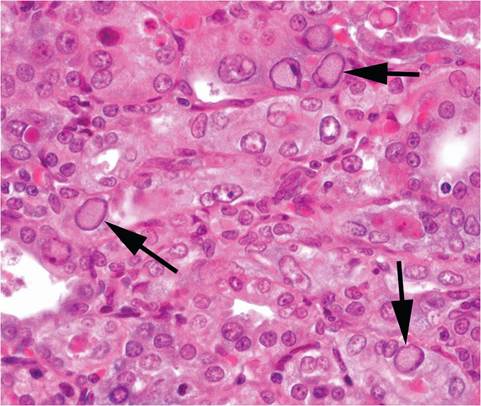
FIG. 1.20. Intranuclear inclusions in the renal tubular epithelium (arrows) of a mouse experimentally infected with polyoma virus.
virus replication in over 40 different cell types, underscoring the virus polytropism. Intranuclear inclusions can be observed with difficulty in cytolytic lesions and are most apparent in renal tubular epithelium (Fig. 1.20). Many of these foci give rise to transformed cells without virus replication. Cytopathic and proliferative changes are especially apparent in bronchiolar, renal pelvic, and ureteral epithelium. When genetically susceptible neonatal mice are experimentally infected with oncogenic strains of the virus, tumors arise most commonly from mammary gland, salivary gland, and thymus. Multiple skin tumors of hair follicle origin (with notable similarity to the skin tumors that occur in hamsters under natural conditions when exposed to hamster polyomavirus) are frequently observed. Tumors of mesenchymal origin are also common, including renal sarcomas, osteosarcomas, hemangiomas, and fibrosarcomas. Experimentally infected mice develop runting syndrome, polyarteritis, and enhanced autoimmune disease.
Accidental infection of nude mice has been shown to result in multisystemic wasting disease, with paralysis and development of multiple tumors, particularly of uterus and bone. Experimental infections of nude mice yielded a very high prevalence of mammary adenocarcinomas among females and osteosarcomas among males. Nude mice can also develop infection of oligodendroglia with demyelination, similar to progressive multifocal leukoencephalopathy (PML) in immmuno- compromised humans infected with BK and JC viruses, and macaques infected with SV-40 virus. Paralysis in nude mice is due to vertebral tumors as well as demyelination. A single report examined the effects of intraperitoneal inoculation of C.B-17-scid and B6-scid mice. Mice of both types became acutely ill and died within 2 weeks, with cutaneous hemorrhages and splenomegaly. The mice were thrombocytopenic, with depletion of megakaryocytes in spleen and bone marrow, and developed marked extramedullary myelopoiesis in the spleen, which was misinterpreted as “myeloproliferative disease.” If PyV gains access to other types of immunodeficient mice, it is likely to behave in a manner that is dictated by the properties of the virus, genetic background of the mouse strain, and nature of the immunodeficiency, but the rarity and inefficient transmission of PyV have limited the chances of natural exposure.
Diagnosis
The presence of PyV in immunocompetent mouse populations is best detected serologically, but PCR has been developed for detecting virus in tissues and other biological products. Differential diagnoses of nude mice with wasting disease include primarily mouse hepatitis virus, Pneumocystis murina, Sendai virus, and pneumonia virus of mice (PVM). Microscopic lesions containing intranuclear inclusion bodies must be differentiated from lesions caused by K virus, MAdV, and MCMV.
K Virus Infection
For all practical purposes, K virus is of historical interest and occurs rarely, if at all, in contemporary laboratory mouse colonies. Unlike PyV, K virus has no oncogenic action, either naturally or experimentally, in keeping with the absence of middle T antigen.
Epizootiology and Pathogenesis
K virus was initially discovered by Lawrence Kilham (thus the K) following intracerebral inoculation of infant mice with tissue extracts from an adult mouse during experiments on the mammary tumor virus. K virus is spread by the oronasal route. When orally inoculated into neonatal mice, the virus initially replicates in intestinal capillary endothelium and then disseminates hem- atogenously to other organs, including lung, liver, spleen, kidney, and brain, where it replicates in vascular endothelium. There appears to be strong tropism for pulmonary endothelium. At 6-15 days after inoculation of neonatal mice, there is a sudden onset of dyspnea due to pulmonary vascular edema and hemorrhage, resulting in rapid death. Pulmonary disease does not occur when older mice are inoculated, with complete resistance evolving between 12 and 18 days of age. Older mice mount an early and effective immune response that prevents the viremic phase of infection. Infection of nude mice results in disease similar to that seen in suckling mice. Regardless of age, mice remain persistently infected, and the site for virus persistence is renal tubular epithelium, which is typical of other members of the polyomavirus group. In naturally infected colonies, clinical signs are absent, with dams conferring passive immunity to litters during the disease-susceptible neonatal period.
Pathology
Gross lesions are restricted to lungs of neonatal or immunodeficient mice. Microscopically, intranuclear inclusions are present in vascular endothelium of jejunum, ileum, lung, liver, and occasionally brain. Inclusions are poorly discernible and require optimal fixation, especially in the intestine. Pulmonary lesions consist of congestion, edema, hemorrhage, atelectasis, and septal thickening. Livers of neonatal mice can have sinusoidal leukocytic infiltration and nuclear ballooning of cells lining sinusoids. Lymphocytic infiltrates (including interstitial pneumonia) arise in recovering mice. Inclusions may be found in renal tubular epithelial cells, frequently in groups of 2 or more, associated with foci of interstitial inflammation.
Diagnosis
Recognition of diagnostic lesions is difficult at best and is most likely in neonatally infected mice. Serological surveillance can be carried out by a variety of methods. PCR has been utilized to detect virus in infected mouse tissues. Differential diagnosis of multisystemic infection with intranuclear inclusions should include PyV, MAdV- 1, and MCMV.
Poxvirus Infection: Ectromelia Virus
Infection; Mousepox
No virus of laboratory mice conjures up an image of ruin like ectromelia virus (ECTV). Some of this reputation is justified, but most is human in origin. ECTV is a large DNA virus of the family Poxviridae and genus Orthopox, to which vaccinia, variola, monkeypox, cowpox, and others also belong. Orthopoxviruses share extensive antigenic cross-reactivity. Each is a distinct species, but the host range can be broad. Marchal reported an epizootic disease with high mortality in adult mice and termed it “infectious ectromelia,” because of the frequency of limb amputation (ectromelia) in surviving mice. Frank Fenner performed seminal work on pathogenesis of the agent “ectromelia virus,” which causes the disease “mousepox,” although the terms are often erroneously interchanged. Outbreaks in the United States have stimulated renewed interest in the pathogenesis of mousepox. Mousepox was originally studied as a model for human smallpox, and that interest re-emerged with bioterrorism research.
Epizootiology and Pathogenesis
The origin of ectromelia virus remains an enigma, since it has never been found in wild populations of M. musculus. Unsubstantiated evidence has suggested infection of wild non-Mus rodents in Europe, but it has not been confirmed by appropriate sequence analysis and may actually reflect infection with cowpox virus. Ectromelia virus was at one time common among mouse colonies in Europe and may be enzootic in laboratory mice in China. In the past, outbreaks in the United States have been due to introduction of infected mice or mouse products from Europe. In more recent outbreaks of mousepox, the source of the virus was traced to commercial mouse sera either collected from mice in the United States or imported from China. Strains of ECTV, including Hampstead, Moscow, NIH-79, Washington University, St. Louis 69, Beijing 70, Ishibashi I-III, and NAV, vary in virulence, but are serologically and genetically homogeneous. The NAV strain, which was isolated from mice infected with commercial mouse serum from China, is essentially the same virus as the original Hampstead virus. These findings strongly suggest that ECTV has not had a long or widespread enzootic history among commensal or domestic mice. ECTV is not highly contagious. It can be experimentally transmitted via a number of routes, but the primary means of natural transmission is believed to be through cutaneous trauma, which requires direct contact, and transmission has been shown to be facilitated by handling. Young mice suckling immune dams are protected by maternal antibody from disease but not from infection. ECTV readily infects the placenta and fetuses, but infected fetuses die and are not a source for vertical transmission within a population.
The hypothetical model of infection involves invasion through skin or mucous membranes, local replication, spread to regional lymph nodes, primary viremia, and then replication in spleen and liver. Between 3 and 4 days after exposure, a secondary viremia ensues, inducing replication of virus in skin, kidney, lung, intestine, and other organs. There is increasing evolution of lesions (disease) on days 7-11, including cutaneous rash. This scenario differs markedly between mouse genotypes. Susceptible mouse strains, such as C3H, A, DBA, SWR, CBA, and BALB/c, die acutely with minimal opportunity for virus excretion. Several other mouse strains develop illness but survive long enough to develop cutaneous lesions with maximal opportunity for virus shedding. Others, such as B6 and AKR mice, are remarkably resistant to disease and, therefore, allow inefficient virus replication and excretion. Thus, a textbook mousepox epizootic requires a select combination of introduction, suitable mouse strains for transmission, and the presence of susceptible strains for disease expression. Immunosuppression will exacerbate disease in mildly or subclinically infected mice. For these reasons, classic outbreaks of high mortality are often not seen in genetically homogeneous colonies of mice. Immunologically competent mice recover completely from infection and do not generally serve as carriers. Therefore, rederivation of virus-free mouse populations can be achieved from immmunocompetent mice. Immunodeficient mice cannot clear virus and are likely to be highly susceptible to fatal disease. Susceptibility to ECTV is dependent upon age, sex, strain, and immune status of the host and the virus strain. Interferons, NK cells, T cells, and B cells are all important. The genetics of resistance are complex and polygenic and not linked to H-2 haplotype. Resistance
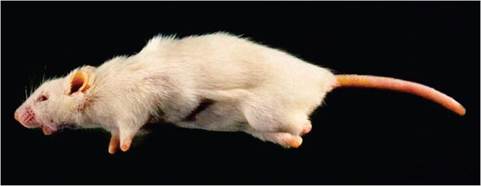
FIG. 1.21. Healed amputating lesions of the distal extremities (ectromelia) from a mouse that survived natural infection with ectromelia virus. (Source: R. Feinstein, The National Veterinary Institute, Sweden. Reproduced with permission from R. Feinstein.)
factors have been mapped to loci on chromosome 6 that includes the NK cell complex, chromosome 2 that includes the gene for C5, chromosome 17, and chromosome 1.
Pathology
Expression of lesions is dependent upon factors discussed above. Clinical signs range from subclinical infection to sudden death. External lesions during the acute phase of infection in susceptible surviving mice include conjunctivitis, alopecia, cutaneous erythema and erosions (rash), and swelling and dry gangrene of extremities, which result in “ectromelia” in surviving mice (Fig. 1.21). Internally, livers may be swollen, friable, and mottled with multiple pinpoint white to coalescing hemorrhagic foci (Fig. 1.22). Spleens, lymph nodes, and Peyer's patches may be enlarged, with patchy pale or
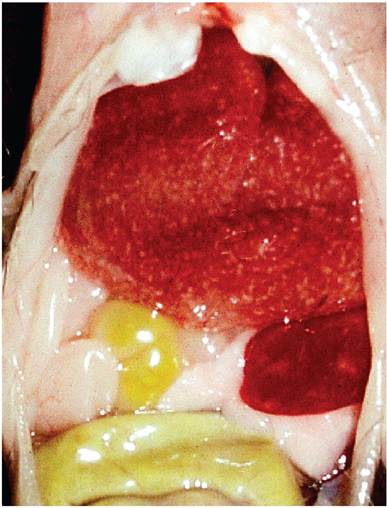
FIG. 1.22. Multifocal necrotizing hepatitis and splenitis in a mouse during the acute phase of mousepox. (Source: Labelle et al. 2009. Reproduced with permission from American Association for Laboratory Animal Science.)
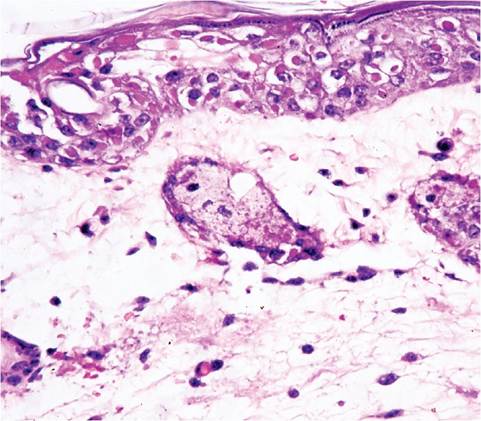
FIG. 1.23. Skin from a mouse infected with ectromelia virus. Note ballooning degeneration and intracytoplasmic inclusions in the epidermis, with underlying dermal edema.
hemorrhagic areas. Intestinal hemorrhage, particularly in the upper small intestine, is common. Microscopic lesions consist of focal coagulative necrosis in the liver, spleen, lymph nodes, Peyer's patches, and thymus, as well as other organs. Multiple basophilic to eosinophilic intracytoplasmic inclusion bodies (1.5-6 μm) are evident in infected cells, especially hepatocytes at the periphery of necrotic foci. These inclusions are poorly discernible with routine staining, but can be enhanced by doubling hematoxylin-staining time. Lymphoid tissue can be hyperplastic and/or focally necrotic, with occasional eosinophilic cytoplasmic inclusion bodies (type A pox inclusions or Marchal bodies). Erosive enteritis, often in association with Peyer's patches, is common, with type A inclusions in enterocytes. Skin lesions consist of focal epidermal hyperplasia, with hypertrophy and ballooning of epithelial cells and formation of numerous prominent large type A inclusions (Fig. 1.23). Later, skin lesions become erosive and inflammatory in character. Inclusions, inflammation, and erosion are also found in conjunctiva, vagina, and nasal mucosa. The conjunctival mucosa is a preferred area to search for inclusion bodies. Recovered mice often have fibrosis of the spleen and can have amputated tails and digits.
Diagnosis
The variable clinical signs and lesions can be problematic, but careful selection of clinically ill mice will enhance an accurate diagnosis. The complex of liver, spleen, and epithelial lesions bearing typical inclusions is pathognomonic. Splenic fibrosis in recovered mice is also a unique feature of this disease. Confirmation can be achieved by electron microscopic identification of the strikingly large poxvirus particles, immunohistochemistry, PCR, or virus isolation. Serology is a useful diagnostic adjunct in recovered mice and is an important surveillance tool for monitoring mouse populations. Serologic testing is likely to be of little value during the early stages of the infection. Vaccination is variably practiced and can interfere with interpretation of serology results. It should also be noted that vaccination protects mice from severe disease, but still allows active infection. Differential diagnoses must include agents that cause hepatitis in adult mice, such as MHV, Tyzzer's disease, salmonellosis, and others. Skin lesions must be differentiated from bite wounds, trichotillomania, hypersensitivity, and other forms of dermatitis. Gangrene and amputation of digits or tail can also occur due to trauma or “ringtail.” Draconian depopulation measures to eliminate ECTV from mouse populations are probably not necessary if a rational approach to quarantine, testing, and rederivation is taken based upon the biology of this virus.