Arenavirus Infection: Lymphocytic Choriomeningitis Virus Infection
Mus musculus is the natural reservoir host for lymphocytic choriomeningitis virus (LCMV), and mice have carried this virus throughout the world from their original Old World niche.
LCMV is a significant pathogen of humans, and was initially discovered during investigation of St. Louis encephalitis in 1933, when human brain material was injected into the brains of monkeys and mice, which developed lymphocytic choriomeningitis. The lesion, lymphocytic choriomeningitis, is not a feature of natural infection in mice. LCMV belongs to the family Arenaviridae, which includes a single genus Arenavirus, named because of the granular-appearing (Latin arenosus, “sandy”) ribosomes within virions. If for no other reason, LCMV is important because of its significant zoonotic potential. LCMV has been studied extensively as a model system of immune-mediated disease, virus persistence, and immune tolerance, resulting in emphasis on aspects of infection that are not necessarily relevant to natural infection. LCMV has also been used as a model of noncytolytic viral disruption of differentiated cell function, resulting in disease without lesions (a claim that has not involved pathologists). LCMV is an unacceptable agent in laboratory animal facilities, and its eradication should be aggressively effected. The polytropic nature of LCMV and its wide host range allow this virus to readily infect transplantable tumors and cell lines, which can serve as a source of contamination for mouse colonies.Epizootiology and Pathogenesis
LCMV is not ubiquitous, in that isolated mouse populations may or may not be enzootically infected. It is fortunately rare among contemporary laboratory mouse
populations, but may be a contaminant of biological products derived from mice. LCMV has also been found in pet mice and colonies of mice raised for feeding other species, including non-human primates.
LCMV can naturally infect a variety of other mammals, including hamsters, guinea pigs, cotton rats, chinchillas, canids, and primates, including humans. Newborn rats can be infected experimentally, but this species seems to be refractory to natural infection. Among mice, the highly labile virus can be transmitted by direct contact and aerosol through nasal secretions as well as urine and saliva.Enzootic infection in a mouse population is maintained by vertical transmission from dam to fetus and neonates. Infection of the fetus occurs during early pregnancy, and ova can be infected prior to implantation. There is no evidence for transmission by coitus. Nearly every cell in the fetus may become infected non- cytolytically, with no significant adverse effect, although reduced litter sizes and runted pups may occur. The widely disseminated fetal infection involves the immature thymus, resulting in selective immune tolerance, with negative selection and depletion of LCMV- responsive CD8 T cells. This state of tolerance is highly LCMV selective, with otherwise normal immune responsiveness to other antigens. A similar scenario can occur if pups are infected as neonates. The state of LCMV immune tolerance allows multisystemic, persistent, sub- clinical infection, with the mice growing to reproductive maturity and perpetuating the cycle to the next generation. Immune-tolerant adult females that were infected as neonates can not only infect their own young in utero but also do not confer passive immunity to neonates, which further facilitates spread of the virus due to communal nursing behavior. Tolerance is not absolute, as mice develop LCMV-specific antibody, but antibody is non-neutralizing and complexed with excess viral antigen, which tends to be deposited in tissues, including arterial walls, choroid plexus, and glomeruli. Eventually, tolerance breaks down further, resulting in chronic lymphocytic infiltrates in multiple tissues and immune complex glomerulonephritis (late disease).
The onset of late disease varies with genetic strain of mice. This phenomenon is irrelevant in wild mice, since mice become ill at an age in which their reproductive contributions to the population are no longer essential. LCMV immune-tolerant mice with disseminated infection can also develop a number of endocrine disorders due to diminished secretion of growth hormone with hypoglycemia, diabetes with hyperglycemia and abnormal glucose tolerance, and decreased thyroxine and thyroglobulin. These phenomenas are due to noncytolytic infection of endocrine organs, resulting in disturbance of cell function.In contrast, natural or experimental infection of adult, immunocompetent mice follows a distinctly different course. Experimental infection results in a wide variety of disease manifestations, depending upon host and virus factors. Following natural exposure, or natural routes of experimental inoculation (intranasal or oral), immunocompetent adult mice typically develop shortterm, acute infections from which they recover and seroconvert. However, parenteral inoculation with “aggressive” strains of virus results in disseminated infection of multiple organs, followed by a host immune response with CD8 T-cell-mediated disease. When virus is inoculated intracerebrally, disease is characterized by immune-mediated lymphocytic choriomeningitis (especially with “neurotropic” strains), whereas intraperitoneal inoculation results in immune-mediated hepatitis. In contrast, inoculation of mice with high doses of virus that are “docile” and “viscerotropic” results in immune exhaustion (in contrast to immune tolerance) and therefore mice develop no clinical, T-cell- mediated disease (thus, the term “docile”). The mechanism for immune exhaustion is selective targeting and high affinity for alpha-dystroglycan receptors on dendritic cells. The virus thus initially targets dendritic cells in the marginal zones of the spleen and lymph nodes, and then spreads to T-cell regions, with subsequent T-cell-mediated immune destruction of infected lymphoid tissue.
This cycle of infection and destruction results in massive depletion of lymphoid tissues, including thymus, spleen, and T-cell regions of lymph nodes. This immune exhaustion results in global immunodeficiency, in contrast to the selective immune tolerance that takes place in fetally or neonatally infected mice. In both scenarios, virus infection is persistent.LCMV strains and isolates cannot be differentiated serologically, as LCMV is a monotypic quasispecies. Nevertheless, there are a number of clonal laboratory strains with differing experimental tissue tropism and biological behavior, including Armstrong, Traub, WE, Pasteur, and others. Experimental disease is dependent upon virus strain, dose, route of inoculation, and host factors, including age, strain, and immunocompetence. A significant determinant of susceptibility to the adult, immune-mediated form of experimental disease is linked to the H-2 locus. Mice with H-2q∕q (e.g., SWR) or H-2q∕k (e.g., C3H.Q) haplotypes are disease susceptible, whereas mice of the H-2k∕k haplotype (e.g., C3H/ He) are disease resistant. H-2 haplotype is associated with CD8 T-cell responsiveness, but CD4 T cells, B cells, NK cells, and interferons, among other factors, contribute to LCMV immunity. The adult form of infection following parenteral inoculation is largely an experimental phenomenon, but it provides insight into the outcome of inoculation of mice with contaminated biological material. LCMV is a frequent contaminant of transplantable tumors. Considering the wide variety of immune-deficient mice being utilized today, and the many immune factors that are determinants of controlling outcome of infection or disease, awareness of the full spectrum of LCMV biology is useful.
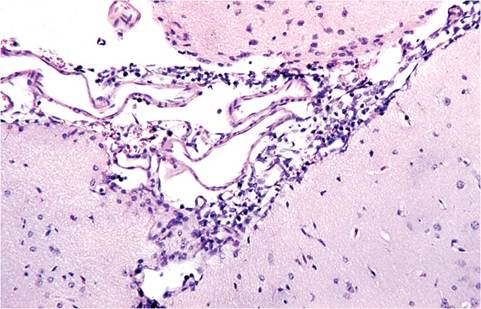
FIG. 1.24. Lymphocytic infiltration of meninges of a mouse inoculated intracerebrally with lymphocytic choriomeningitis virus (LCMV).
Pathology
Clinical signs of natural LCMV infection are minimal, but can include runting in infant mice, reduced litter size, and chronic wasting in older mice if infection is occurring in utero within the colony. Microscopic lesions are likewise nonspecific and most likely to be found in persistently infected aged mice, which may develop vasculitis, glomerulonephritis, and lymphocytic infiltration in multiple tissues, including brain, liver, adrenal, kidney, and lung. Acute disease is largely an experimental phenomenon arising from parenteral inoculation with high doses of virus. Infection with “aggressive” strains of virus results in generalized lymphadenopathy with lymphoid hyperplasia, lymphocytic infiltrates in multiple tissues, necrotizing hepatitis, and lymphocytic choriomeningitis (especially if inoculated intracerebrally) (Fig. 1.24). Infection with “docile” strains of virus results in severe generalized lymphoid depletion in T-cell regions of the thymus, spleen, and lymph nodes. Although natural infection of nude mice has been documented, pathology was not described.
Diagnosis
Definitive diagnosis of LCMV infection cannot be based on pathology. Serology with recombinant nucleoprotein antigen has circumvented the hazards of growing the virus for antigen. However, serology may be problematic, since horizontal infection among adult mice is inefficient and likely to cause seroconversion among a very few mice within a population. Mice infected in utero or as neonates are immune-tolerant to LCMV and do not efficiently seroconvert, or circulating antibody is complexed with antigen. Adult mice infected with docile types of virus may have immune exhaustion, and not seroconvert. Thus, serological testing must be applied to a large sample size and can be enhanced by cohousing adult sentinel mice with mice suspected to be persistently infected. LCMV can be confirmed in suspect tissues with a variety of bioassay approaches, such as
MAP testing, but this has been supplanted by PCR. Differential diagnoses for runting in infant mice include a number of other viral infections. Chronic illness in older mice must be differentiated from generalized lymphoproliferative disorders, amyloidosis, glomerulonephritis, and chronic renal disease of aging mice.
More on the topic Arenavirus Infection: Lymphocytic Choriomeningitis Virus Infection:
- LYMPHOCYTIC CHORIOMENINGITIS VIRUS INFECTION
- Lymphocytic Choriomeningitis Virus Infection
- Picornavirus Infection: Mouse Encephalomyelitis Virus Infection
- Arterivirus Infection: Lactate Dehydrogenase-Elevating Virus Infection
- Coronavirus Infection: Mouse Hepatitis Virus Infection
- WEST NILE VIRUS INFECTION
- Cowpox Virus Infection
- Sendai Virus Infection
- Pneumonia Virus of Mice (PVM) Infection
- LJUNGAN virus infection
- Pneumonia Virus of Mice Infection
- USUTU VIRUS INFECTION
- Hamster Polyoma Virus Infection: Transmissible Lymphoma
- Neurological complications of HIV-1 infection, due either to the immunosuppression (opportunistic infections and neoplasms) or the neurotropism of the virus, are common and add considerably to the morbidity and mortality of the infection
- Streptococcus pneumoniae Infection: Pneumococcal or Diplococcal Infection
- Parvovirus Infection
- Pasteurella pneumotropica Infection
- NEUROLOGICAL DYSFUNCTION ASSOCIATED WITH HIV-1 INFECTION
- INFLAMMATION IN HIV-1 INFECTION
- CD8+ T LYMPHOCYTES IN HIV-1 INFECTION