Coronavirus Infection: Mouse Hepatitis Virus Infection
Mouse hepatitis virus (MHV) belongs to the family Coronaviridae. MHV is very common among wild and conventionally housed mouse populations, and is represented by numerous antigenically and genetically related strains that vary considerably in their virulence and organotropism.
This is due to the high propensity of coronaviruses to mutate and recombine. Despite “hepatitis” in its name, MHV is not always hepatotropic. MHV shares antigenic cross-reactivity with other (Group 2) coronaviruses, which includes human coronavirus OC43, bovine coronavirus, rat coronavirus, and hemagglutinating encephalomyelitis virus of swine. Although the role of these related viruses is often questioned as sources of unexplained MHV outbreaks, there is no evidence that MHV comes from anywhere but mice and mouse products. The polytropism of some MHV strains contributes to their propensity to infect a variety of mouse-derived biological products, including transplantable tumors, cell lines, ES cells, and hybridomas. MHV has protean effects upon a wide variety of research variables, particularly immune response.Epizootiology and Pathogenesis
MHV was initially recognized in mice with neurologic disease in 1949 and subsequently isolated from a variety of cell cultures, tumors, clinically ill mice with a variety of disease manifestations, and mice with abnormal responses to research variables. Despite its prevalence and potential pathogenicity, clinical MHV disease is not common. The outcome of infection depends upon the interaction of different MHV strains on host variables, which include age, genotype, and immune status, including maternally derived antibody.
MHV strains can be divided into two biologically distinct but overlapping groups. Respiratory strains possess primary tropism for upper respiratory mucosa and enterotropic strains have primary tropism for entero- cytes.
Those strains with respiratory tropism initially replicate in nasal mucosa and disseminate to a variety of other organs because of their polytropic nature. Representatives of this type of MHV include the prototype strains MHV-JHM, MHV-A59, MHV-S, and MHV-3, among others. Disseminated infection to multiple organs is favored by virulent strains of virus, mice less than 2 weeks of age, genetically susceptible strains of mice, or immunocompromised mice. Dissemination of MHV from the nose occurs via the blood and lymphatics to pulmonary vascular endothelium and draining lymph nodes, respectively. Secondary viremia disseminates virus to multiple organs, with virus replication and cytolytic lesions in central nervous system, liver, lymphoid tissues, bone marrow, and other sites. Infection of central nervous system by viremic dissemination occurs primarily in neonatal or immunodeficient mice but not older, immunocompetent mice. Direct infection of adult mouse brain can also occur by extension of the virus along olfactory neural pathways, even in the absence of dissemination to other organs. After approximately 5-7 days, immune-mediated clearance of the virus begins, with no persistence or carrier state beyond 3-4 weeks. Infection of mice after weaning age is usually subclinical, particularly in natural infections caused by generally nonvirulent strains of virus. The obvious exception is immunodeficient mice, which cannot clear the virus and develop progressively severe disease. They can die acutely or develop chronic wasting disease if infected with relatively avirulent strains of MHV.Mouse genetic susceptibility and resistance to polytropic MHV has been studied extensively. BALB/c mice are generally quite susceptible to MHV, whereas SJL mice are remarkably resistant. Susceptibility to MHV-A59 and MHV-JHM has been linked to allelic variation of carci- noembryonic antigen-related cell adhesion molecule 1 (CEACAM1). MHV does not necessarily respect such clear explanations, since these differences are quite virus strain-specific, and other MHV strains utilize alternative cellular receptors.
MHV-3 is widely known to be highly virulent in genetically susceptible BALB/c and DBA/2 mice, but is less virulent in semisusceptible C3H and disease-resistant A/J mice. MHV-3 disease severity is determined by thrombosis and coagulation necrosis due to induction of procoagulant activity by macrophages in susceptible, but not resistant, mice. Most polytropic MHV strains display neurotropism following intranasal inoculation of various mouse genotypes, including A, BALB/c, CBA, C3H/He, and C3H/Rv but not SJL mice. These experimental examples emphasize the point that the biologic behavior of wild-type MHV is likely to be unpredictable.Enterotropic MHV strains tend to selectively infect intestinal mucosal epithelium, with minimal or no dissemination to other organs, even in immunodeficient mice. Enterotropic strains that have been described include MHV-S/CDC, MHV-Y, MHV-RI, and MHV-D, among others. Early descriptions of lethal intestinal virus of infant mice (LIVIM) are consistent with enterotropic MHV, but the LIVIM agent was lost before it could be fully characterized. All ages and strains of mice are susceptible to enterotropic MHV infection, including SJL mice, which are resistant to polytropic MHV. Disease occurs only in infant mice due to their intestinal mucosal proliferative kinetics. Infection of neonatal mice results in severe necrotizing enterocolitis with high mortality within 48 hours after inoculation. Mortality and lesion severity diminish rapidly with advancing age at inoculation. Adult mice develop minimal lesions, but replication of equal or higher titers of virus occurs, compared with neonates. The severity of intestinal disease is associated with age-related intestinal mucosal proliferative kinetics, rather than immune-related susceptibility. To underscore this point, disease is minimal in nude or SCID mice infected as adults. Recovery from enterotropic MHV is immune-mediated and requires functional T cells. No persistent carrier state seems to occur in recovered, immunocompetent mice, but persistent infection and virus shedding in feces can occur in T- and B-cell-immunodeficient mice, as well as transgenic mice without known immune dysfunction.
Thus, biologic behavior, in particular duration of infection, is unpredictable in GEMs. Enterotropic MHV infections are often complicated by other opportunistic pathogens, including Escherichia coli and Spironucleus muris.Host immunity to MHV is strongly MHV strain specific. Recovery from one MHV strain provides strong resistance to re-exposure with the homotypic strain, but only partial or no resistance to infection with an antigenically heterotypic strain. Innate immunity and both cellular and humoral arms of the acquired immune response are important in controlling infection, but T cells are critical for virus clearance and recovery from infection. Maternally derived passive immunity is critically important in MHV epizootiology. The highly contagious nature of some MHV strains, particularly enterotropic MHV, within naive mouse populations can result in high mortality among neonatal mice. Pups suckling immune dams, on the other hand, are completely protected from infection, and subsequently acquire infection at an age in which they are not apt to develop severe clinical disease. Maternal protection against polytropic MHV strains is mediated through serum IgG that is passed in utero to the fetus or post- natally through intestinal IgG receptors. In contrast, maternal protection against enterotropic MHV is mediated through luminal whey containing IgA and IgG. Maternally derived passive immunity, like active immunity, is also MHV strain specific, providing protection against homotypic, but only partial or no protection against heterotypic strains of MHV. In spite of the fact that most MHV infections at the level of a single mouse are acute with recovery, the high rate of MHV mutation within an infected population and the strain-specific nature of active and passive immunity contribute to and actually select for persistence of virus within a population.
Vertical transmission from infected dams to fetuses has been documented experimentally, but is highly unlikely to occur naturally.
This would require infection of a virus-naive genetically susceptible mouse with a relatively virulent, polytropic strain of MHV during pregnancy. Even if such events take place, fetal infection would be fatal, so live offspring would not be an issue. The zona pellucida is an effective barrier to MHV infection. However, introduction of MHV through ES cells or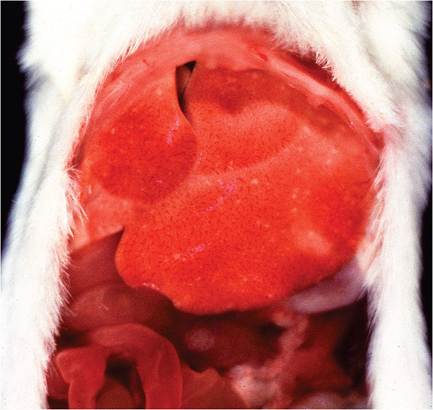
FIG. 1.25. Multifocal hepatitis in a BALB/c mouse infected with mouse hepatitis virus (MHV).
germplasm into naive mouse colonies represents a greater concern. MHV has a well-documented ability to persistently infect cell lines without cytopathic effect, including ES cell lines.
Pathology
The majority of natural polytropic MHV infections are subclinical, with mild or no gross lesions. Immunocompetent mice may have grossly discernable white foci on the surface of the liver (Fig. 1.25), which can be profoundly obvious and often hemorrhagic in immunodeficient mice (Fig. 1.26). In immunodeficient mice infected with relatively low virulence MHV strains, mice may survive long enough to develop hepatic nodular hyperplasia with parenchymal collapse and fibrosis. Foci
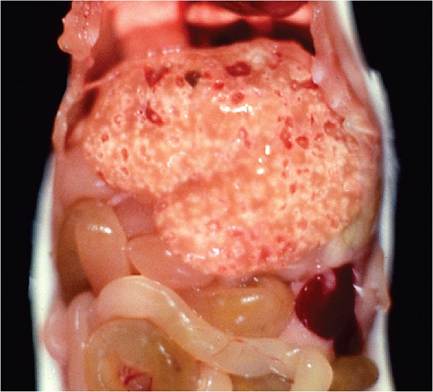
FIG. 1.26. Severe multifocal coalescing hepatic necrosis in a SCID mouse infected with MHV.
of splenic necrosis, with splenomegaly due to extramedullary hematopoiesis, are often present in immunodeficient mice. Neurologic manifestations may be observed in immunodeficient mice, and rarely in immunocompetent mice, including vestibular signs and posterior paresis.
Microscopic findings in mice infected with polytropic MHV strains are characterized by focal acute necrosis and syncytia of parenchymal cells and vascular endothelium in multiple organs, including liver, splenic red and white pulp, lymph nodes, gut-associated lymphoid tissue, thymus, and bone marrow.
Focal peritonitis may also be present. Hallmark syncytia are not as obvious in immunocompetent mice, but residua of syncytia may be found in liver, characterized by large degenerating cells at the periphery of necrotic foci with dense basophilic apoptotic nuclear bodies. Syncytia are common in lesions of immunodeficient mice, which allow lesions to reach their fullest potential (Figs. 1.27A and B). Syncytia of pulmonary vascular endothelium are common in immunodeficient mice (Fig. 1.28). Pancytopenia has been documented in mice experimentally infected with polytropic MHV, and probably occurs in severe natural infections. Bone marrow necrosis, syncytia,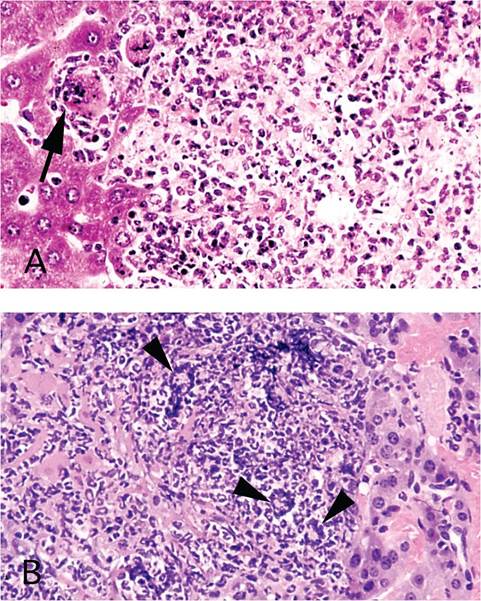
FIG. 1.27. (A and B) Focal hepatitis in mice infected with MHV. Note the degenerated syncytium (arrow) at the periphery of a necrotic focus in an immunocompetent mouse (A), in contrast to the prominent syncytia (arrowheads) that are obvious in the liver of an immunodeficient mouse (B).
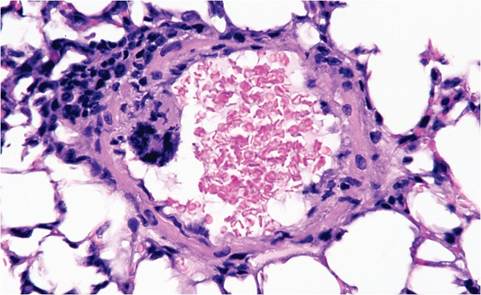
FIG. 1.28. Endothelial syncytium in a pulmonary vessel of an immunodeficient mouse infected with MHV.
and compensatory hematopoietic hyperplasia can be found in naturally infected mice, accounting for the splenomegaly that occurs in immunodeficient mice. Neonatally infected mice and immunodeficient mice can have vascular-oriented necrotizing encephalitis (Fig. 1.29). Mice may also develop nasoencephalitis due to localized infection of olfactory mucosa, olfactory nerves, olfactory bulbs, and olfactory tracts of the brain, with meningoencephalitis and demyelination. This pattern of infection occurs regularly after intranasal inoculation of many MHV strains, but is a relatively rare event by natural exposure.
Lesions due to enterotropic MHV depend primarily upon age of the host. Neonatal mice in naive mouse colonies can experience massive outbreaks of high mortality, presenting as dehydrated, runted pups. These mice have segmentally distributed areas of villus attenuation, enterocytic syncytia (balloon cells), and mucosal necrosis (Fig. 1.30). Eosinophilic intracytoplasmic inclusions may be present, but are not as diagnostic as syncytia. Mesenteric lymph nodes usually contain lymphocytic syncytia, and mesenteric vessels may contain endothelial syncytia.
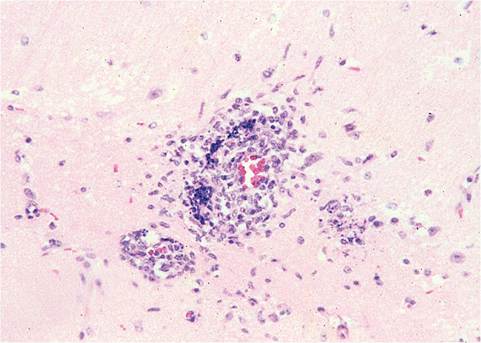
FIG. 1.29. Vascular-oriented encephalitis from an immunodeficient mouse naturally infected with MHV.
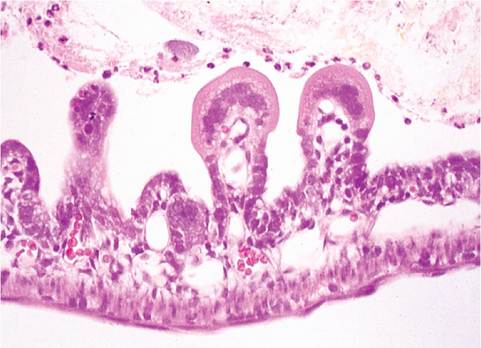
FIG. 1.30. Small intestine of a neonatal mouse infected with enterotropic MHV. Villi are markedly attenuated, with prominent syncytia of villus enterocytes.
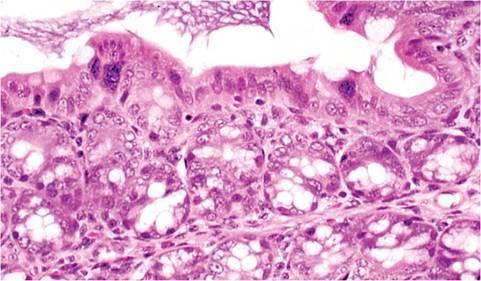
FIG. 1.31. Ascending colon from a mouse with MHV infection. Syncytia are present on the mucosal surface. The ascending colon is the most common site to find MHV syncytia in adult mice.
Surviving mice develop compensatory intestinal mucosal hyperplasia. Lesions are most likely to be found in the terminal small intestine, cecum, and ascending colon. Lesions are progressively milder with increasing age at the time of exposure. Adult mice have minimal lesions, except for enterocytic syncytia in surface mucosa, particularly in cecum and ascending colon (Fig. 1.31). Immunodeficient mice develop similar but progressive lesions, depending upon age at inoculation. Natural enterotropic MHV infection has been described in adult nude mice with chronic hyperplastic typhlocolitis and mesenteric lymphadenopathy. More typically, intestinal lesions in immunodeficient mice, including naturally infected nude mice, are remarkably mild with minimal hyperplasia. Enterotropic MHV strains do not generally disseminate, but hepatitis and encephalitis can occur with some virus strains in certain mouse genotypes.
Residual lesions in mice that have recovered from infection may include reactive hyperplasia of lymph nodes and spleen, hematopoietic hyperplasia, perivascular lymphocytic infiltrates in the lung, and foci of inflammation in liver. Residual brain lesions are usually found in the brain stem and characterized by perivascular cuffing of lymphocytes and vacuolization. Demyelination is a much emphasized feature of experimental brain infections, but is rare under natural circumstances. Unusual presentations may occur in MHV-infected GEMs with specific gene defects. Granulomatous seros- itis (Fig. 1.32), with or without hepatitis or intestinal lesions, has been found in interferon-gamma and inter- feron-gamma receptor null mice, but this pattern of disease has also been observed in mice with other targeted mutations. The serosal exudates contain numerous syncytia (Fig. 1.33) and MHV antigen. It is unknown if this syndrome is related to polytropic or enterotropic MHV.
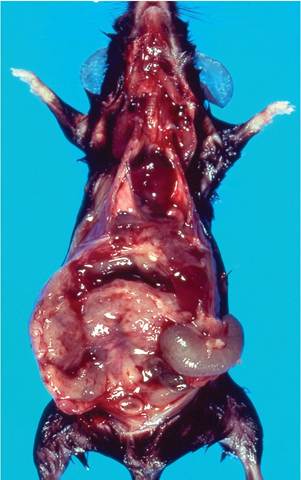
FIG. 1.32. Granulomatous serositis in an interferon-gamma null mouse naturally infected with MHV. (Source: France et al. 1999. Reproduced with permission from John Wiley & Sons Limited.)
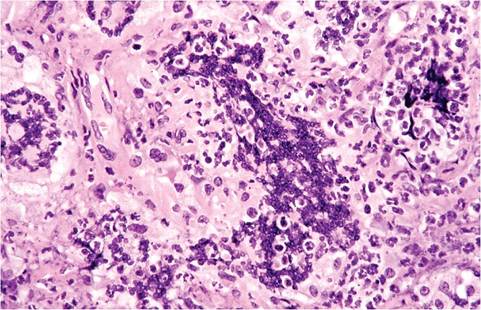
FIG. 1.33. Syncytia in the serosal exudate of an immunodeficient mouse with granulomatous serositis due to MHV infection.
Diagnosis
Diagnosis during the acute stage of MHV infection can be made by visualization of characteristic lesions with syncytia in target tissues, but clinical signs and lesions can be highly variable for reasons discussed above. Active infection can be confirmed by immunohistochemistry, by virus isolation, or PCR. Recovered mice may have perivascular lymphocytic infiltrates in the lung and microgranulomas in the liver. In general, respiratory strains of MHV are polytropic and grow in a number of established cell lines in vitro, but enterotropic MHV strains are far more restrictive and fastidious in vitro. Virus in suspect tissue can be confirmed by a variety of bioassay methods, such as MAP testing or infant or nude mouse inoculation. Amplification by passage in immunodeficient mice will increase the likelihood of in vitro isolation from infected tissue. PCR can be utilized to detect MHV in feces or tissue of infected mice. Serology is the most useful means of surveillance for retrospective infection in a colony. Seropositive mice are poor candidates for pathology workup, since they are likely to have recovered, but on occasion they can be actively infected with a second strain of virus. Nude mice can develop antibody, although their antibody response is unpredictable. There is little merit in attempting to identify MHV strains serologically, since all strains are broadly cross-reactive and antigenic relatedness does not predict virulence or organotropism. Differential diagnoses include salmonellosis, Tyzzer's disease, and mousepox in adult mice, as well as reovirus, cytomegalovirus, and adenovirus in infant mice. Mice with enteritis must be differentiated from epizootic diarrhea of infant mice (EDIM), salmonellosis, Tyzzer's disease, and reovirus infection. Demyelinating lesions must be differentiated from those caused by mouse encephalomyelitis virus, LDV in immunosuppressed AKR or C58 mice, or polyoma virus in immunodeficient mice.
More on the topic Coronavirus Infection: Mouse Hepatitis Virus Infection:
- Picornavirus Infection: Mouse Encephalomyelitis Virus Infection
- Arterivirus Infection: Lactate Dehydrogenase-Elevating Virus Infection
- Arenavirus Infection: Lymphocytic Choriomeningitis Virus Infection
- Rat Coronavirus Infection: Sialodacryoadenitis
- Sendai Virus Infection
- WEST NILE VIRUS INFECTION
- LJUNGAN virus infection
- Cowpox Virus Infection
- Pneumonia Virus of Mice (PVM) Infection
- Lymphocytic Choriomeningitis Virus Infection
- Hamster Polyoma Virus Infection: Transmissible Lymphoma
- LYMPHOCYTIC CHORIOMENINGITIS VIRUS INFECTION
- Pneumonia Virus of Mice Infection