Cellular Anatomy and Morphology
OBJECTIVES
• define, understand, and differentiate between a cellular organelle and an inclusion body
• identify the cellular organelles in a cell model and in a diagram
• understand the structure and function of the cellular organelles
• identify the major cellular organelles using prepared slides
MATERIALS
• three-dimensional model of an animal cell, complete with cellular organelles
• compound microscope
• prepared slide of a giant multipolar neuron
• prepared slide of a Golgi complex
Introduction
The cell is the basic unit of organization of an animal's body.
The cell may also be a singlecelled life form, or cells can combine to make up organized tissue within a more complex animal. Cells are extremely complex and are both the structural and functional unit of the tissues they form. Cells have the ability to maintain their boundaries, metabolize and digest nutrients, dispose of wastes, grow and reproduce, move, and respond to stimuli. Each cell must perform certain functions to sustain its life. Cells are highly diverse; differences in size, shape, and internal composition reflect their specific functions in the body.19
Copyright 2010 Cengage Learning. All Rights Reserved. May not be copied, scanned, or duplicated, in whole or in part. Due to electronic rights, some third party content may be suppressed from the eBook and/or eChapter(s).
Editorial review has deemed that any suppressed content does not materially affect the overall learning experience. Cengage Learning reserves the right to remove additional content at any time if subsequent rights restrictions require it.
The Anatomy of the Cell
Generally, a cell is composed of three major parts: the plasma membrane, the nucleus, and the cytoplasm (Figure 3.1). All of these are readily observable with a light microscope. Within the cytoplasm are the organelles, which are either too small or require a special stain to be seen with ordinary light microscopy.
However, since the advent of the electron microscope, even the smallest organelles have been identified. Organelles, by definition, are highly organized, living, subcellular structures, each of which has a characteristic shape and function. They are, in essence, the internal working parts of the cell. They differ from inclusion bodies, which are non-active masses within a cell.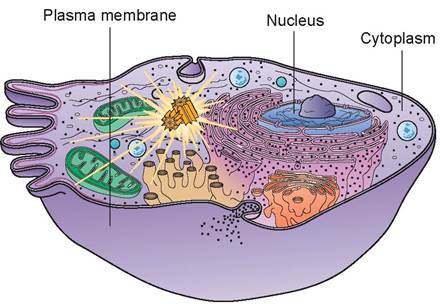
FIGURE 3.1: Structural components of a cell.
Identification of the Parts of a Cell
Using a cell model and Figure 3.2, identify the following parts of a cell.
1. plasma membrane: The plasma membrane is composed of bilayer phospholipids and globular protein molecules. This arrangement of compounds is called the fluid mosaic model of the plasma membrane, and the proteins are said to be floating in a double layer of phospholipids (see Figure 3.2). Some of the externally facing proteins and lipids have sugar (carbohydrate) side chains attached to them that are important in cellular interactions. A few cholesterol molecules are also dispersed in the fluid phospholipid bilayer to help stabilize it. The proteins in the plasma membrane are also responsible for its antigenicity.
The plasma membrane is a flexible, elastic, protective barrier that separates the cell’s internal components from the external environment. The plasma membrane has selective permeability and thus plays an active role in determining what enters and leaves the cell, and in what quantity. Transport through the plasma membrane can occur in two basic ways: actively and passively. In active transport, the cell must provide energy in the form
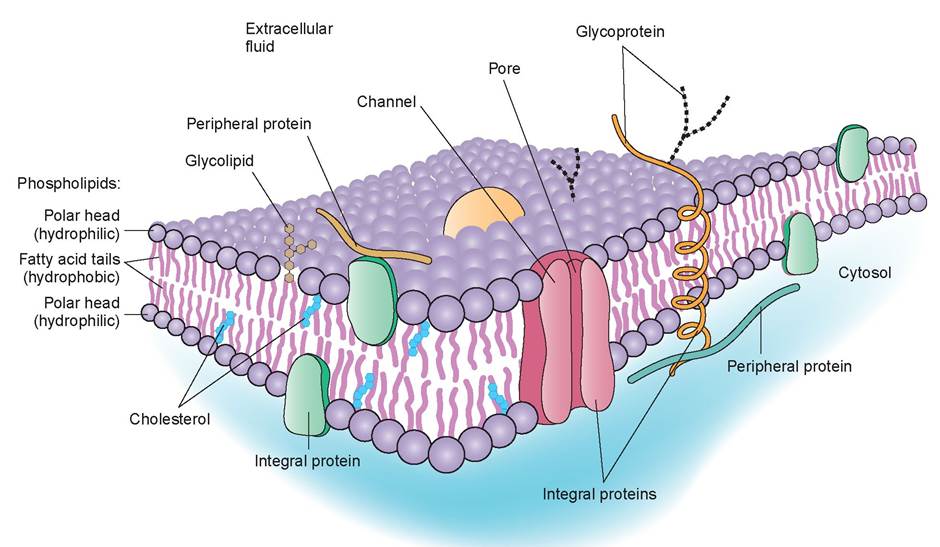
FIGURE 3.2: Structure of the plasma membrane.
of adenosine triphosphate (ATP) to power the movement of substances through the membrane. In passive transport, the movement of substances is driven by concentration or pressure differences.
The plasma membrane is also able to maintain a resting potential, which is absolutely necessary for cells that are excitable, such as nerve or heart cells. The resting potential of a membrane is associated with the type and quantity of ions inside the cell and on the outside. These ions impart a potential difference across the membrane; for example, a nerve cell membrane’s resting potential is about -85 mV. These characteristics play a vital role in cell signaling and cell-to-cell interactions.In addition, cells that are tightly joined together to form functional units of tissue connect to each other at cell junctions. These contact points between plasma membranes may serve one or more of the following three functions: (1) to form fluid-tight seals between cells, (2) to anchor cells together or to anchor them to extracellular material, or (3) to act as channels that allow ions and molecules to pass from cell to cell within a tissue.
2. nucleus: The nucleus is a spherical or oval structure and is the most prominent, visible feature inside the cell when viewed with a light microscope because of its contrasting, dark appearance. Nuclei usually stain a dark blue or purple (Figure 3.3). Most cells have one or more nuclei. For example, a red blood cell has a nucleus during its developmental stage, but the nucleus becomes pyknotic and disappears when the cell becomes mature. In contrast, skeletal muscle cells have numerous nuclei; in other words, they are multinucleated.
The nucleus is the control center of the cell and is necessary for cell reproduction. It is technically an organelle, but it is considered separately because of its numerous and diverse functions. On the exterior of the nucleus is the nuclear envelope. It is similar to the plasma membrane in that it is a bilayered (has two layers) lipid structure. The outer membrane of the nuclear envelope is continuous with the rough endoplasmic reticulum (ER), a membranous cytoplasmic structure that is the site of protein synthesis, and is very similar in structure.
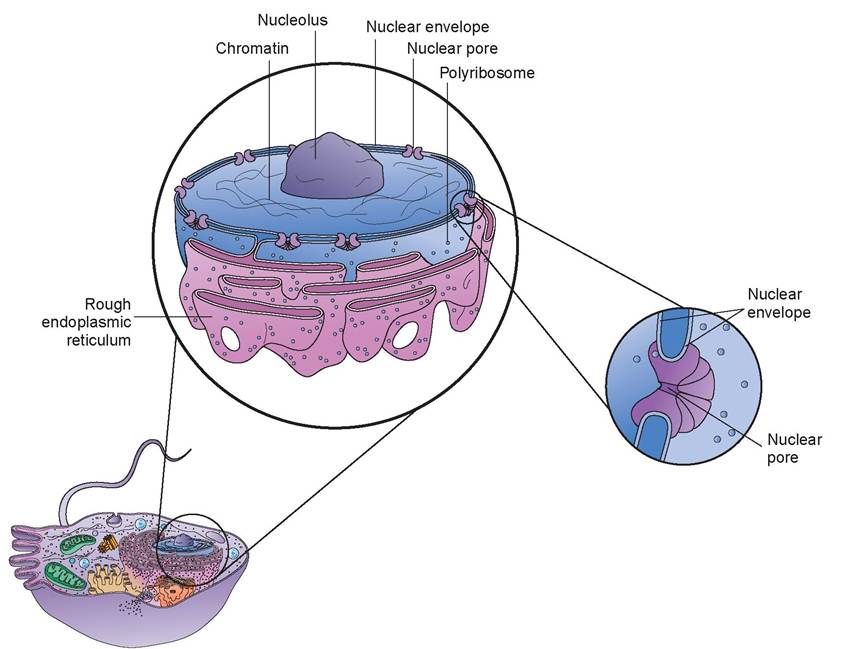
FIGURE 3.3: Nucleus.
The nuclear envelope is also perforated by numerous channels called nuclear pores. Each pore is made up of proteins arranged in a circle, forming a central channel. This channel is approximately 10 times larger than the pore within a protein in the plasma membrane. Nuclear pores control the movement of substances between the nucleus and the cytoplasm. Small molecules can pass via diffusion, but larger molecules, such as RNA and other proteins, must use active transport for passage. This permits the selective transport of proteins from the cytoplasm into the nucleus and of ribonucleic acid (RNA) molecules produced in the nucleus into the cytoplasm.
Another visible feature of the cell is the nucleolus, which is located within the nucleus. There may be one or more nucleoli per nucleus. It is spherical and is composed of a cluster of proteins, deoxyribonucleic acid (DNA), and RNA. It is not enclosed by a membrane. The nucleolus is where ribosomes, which play an important role in protein synthesis, are assembled. Thus, nucleoli are quite prominent in cells that produce large amounts of proteins, such as muscle and liver cells. The nucleoli disperse during cell division and reorganize when the new cells are formed.
The bulk of the nucleus is made up of chromatin, which may appear either diffuse or granular when viewed through a light microscope. Chromatin is the cell’s genetic material, or chromosomes, and is found in a dispersed, unorganized arrangement in the non-dividing cell. The chromosomes and their component genes control cellular structure and most cellular activities. The genes, of course, are the hereditary material, and they ultimately control, among other things, the structure and appearance of the animal.
3. cytoplasm: Cytoplasm is composed of two components: (1) cytosol and (2) organelles. The cytosol is the fluid portion of the cytoplasm and makes up 55% of the total cell volume.
It varies in consistency and composition from one part of a cell to another and is composed of 75% to 90% water, with the balance consisting of various ions, glucose, amino acids, fatty acids, proteins, lipids, ATP, and waste products. Also located in the cytosol are various organic molecules that aggregate into masses that are stored and used as required for metabolism. The cytosol is the site of many chemical reactions and is the place where energy is produced and captured to drive the cellular activities necessary for life. The bulk of the cytoplasm stains pink under a microscope because of the basic (not acidic) proteins that compose the cytosol.4. organelles: The word organelles literally means “small organs.” These structures are the machinery of the cell and are highly specialized to carry out specific functions (Figure 3.4).
a. ribosomes: Ribosomes are dense- staining, spherical bodies composed of ribosomal RNA (rRNA) and protein. They are the actual sites of protein synthesis, or the places where a new amino acid is added to the chain of amino acids in the protein being created. Structurally, a ribosome consists of two subunits, one about half the size of the other. These are formed separately
in the nucleolus; then they exit the nucleus to unite within the cytosol. They may unite on the nuclear membrane or on the endoplasmic reticulum. These are called membrane-bound ribosomes. These ribosomes synthesize proteins destined for insertion into the plasma membrane or for export from the cell. Others may be free in the cytosol, unattached to any structure in the cytoplasm; these are called free ribosomes. Primarily, free ribosomes synthesize proteins used inside the cell. Ribosomes are also located within the mitochondria, where they synthesize mitochondrial proteins. Sometimes 10 to 20 ribosomes join together in string-like arrangements called polyribosomes.
b. endoplasmic reticulum (ER): The ER is a network of membranes that form flattened sacs or tubules called cisterns that extend from the nuclear envelope (as mentioned previously).
The ER is so extensive throughout the cytosol that it constitutes more than half of the membranous surfaces found within the cytoplasm of most cells.There are two forms of endoplasmic reticulum in cells: rough ER and smooth ER. They differ in structure
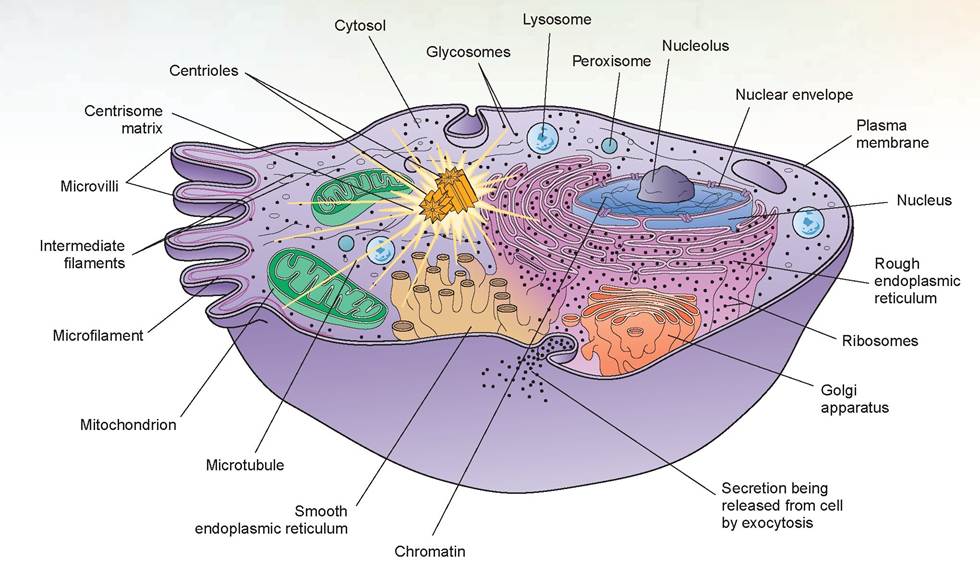
FIGURE 3.4: The cell and its organelles.
and function. The membranous rough ER is continuous with the nuclear envelope and is studded with ribosomes. Proteins synthesized enter the rough ER’s cisterns for processing and sorting (Figure 3.5). As stated previously, these proteins will be exported from the cell or be used in the plasma membrane. In certain cells, enzymes within the cisterns attach the proteins to carbohydrates to form glycoproteins. In other cells, the enzymes attach proteins to phospholipids to form plasma membranes or the membranes of other organelles. Thus, the rough ER is responsible for synthesizing secretory proteins and membrane molecules.
The smooth ER extends off of the rough ER at multiple sites to form another network of membranous tubules. It lacks the ribosomes of the rough ER, but it does contain unique enzymes that make it more functionally diverse than the rough ER. Smooth ER is able to synthesize phospholipids for membrane surfaces, and also synthesizes fats and steroids, such as estrogen and testosterone. In the hepatic cells of the liver, the enzymes of the smooth ER help release glucose into the bloodstream, and they inactivate or detoxify drugs and other
potentially harmful substances, such as alcohol. In muscle cells, calcium ions are released from a form of smooth ER, called the sarcoplasmic reticulum, which triggers the contraction process.
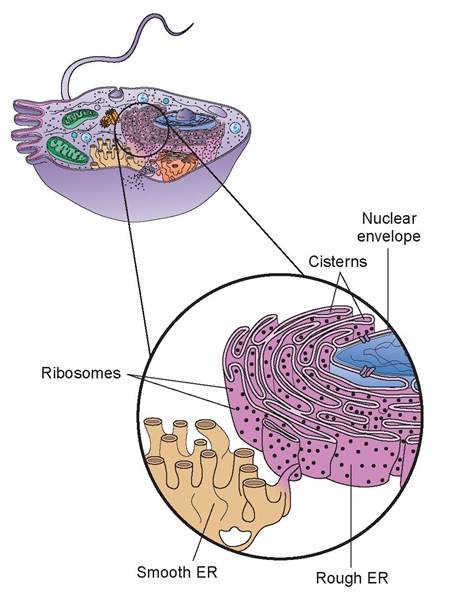
FIGURE 3.5: Endoplasmic reticulum (ER).
c. Golgi complex: The Golgi complex consists of 3 to 20 flattened, membranous sacs stacked on top of one another, with both ends dilated (Figure 3.6). The areas within these sacs are also called cisterns (like the similar areas found in the ER). The overall shape of the Golgi complex is cup-like because of the curvature of the cisterns. Most cells only have one Golgi complex, although some may have several.
In cells that secrete many proteins into the extracellular fluid, the Golgi complex is large and extensive. Its function is to modify, sort, package, and transport products received from the rough ER and form secretory vesicles that discharge processed proteins, via a process called exocytosis, into the extracellular fluid. The Golgi complex also replaces or modifies the existing plasma membrane and forms lysosomes and peroxisomes.
d. lysosomes: Lysosomes are membrane- enclosed vesicles that form in the Golgi complex (Figure 3.7). Inside the lysosomes are as many as 40 different kinds of digestive or hydrolytic enzymes capable of breaking down a variety of molecules. The enzymes tend to work best in an acidic pH, and thus the lysosomal membrane actively transports hydrogen ions (H+) into the lysosomes. The pH inside a lysosome is about 5.0, compared to the pH of the cytosol, which is neutral or 7.0. The lysosomal membrane also allows the final products of digestion, such as sugars and amino acids, to be transported into the cytosol. The functions of the lysosomes are to (1) digest substances that enter the cell via endocytosis, (2) digest worn-out organelles, a process called autophagy, (3) digest the entire cell if it dies, known as autolysis, and (4) occasionally conduct extracellular digestion, such as when spermatozoa release enzymes to help them penetrate the surface of an ovum.
e. peroxisomes: Peroxisome vesicles are smaller than lysosomes and contain one or more enzymes that can oxidize (by removal of a hydrogen atom) various organic substances. Peroxisomes detoxify harmful substances and are therefore abundant in kidney and liver cells. In addition to being produced by the Golgi complex, these organelles can self-replicate.
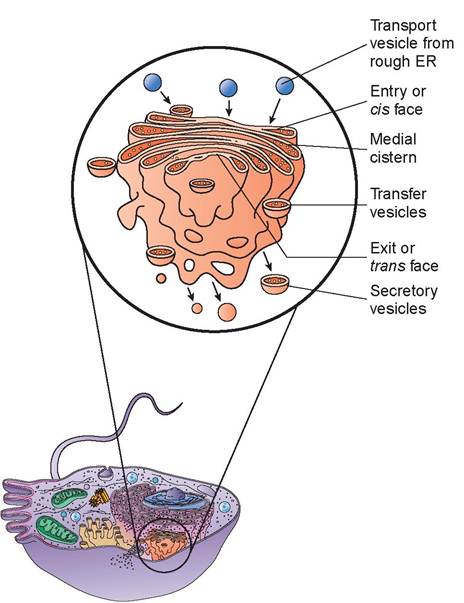
FIGURE 3.6: Golgi complex.
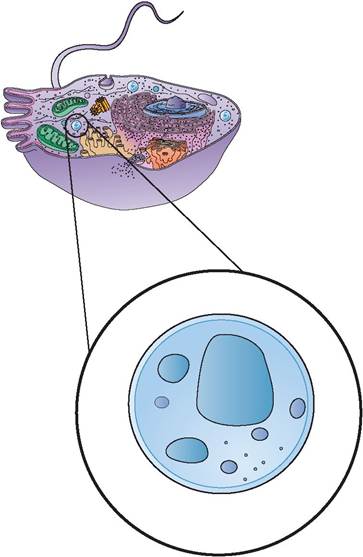
FIGURE 3.7: Lysosome.
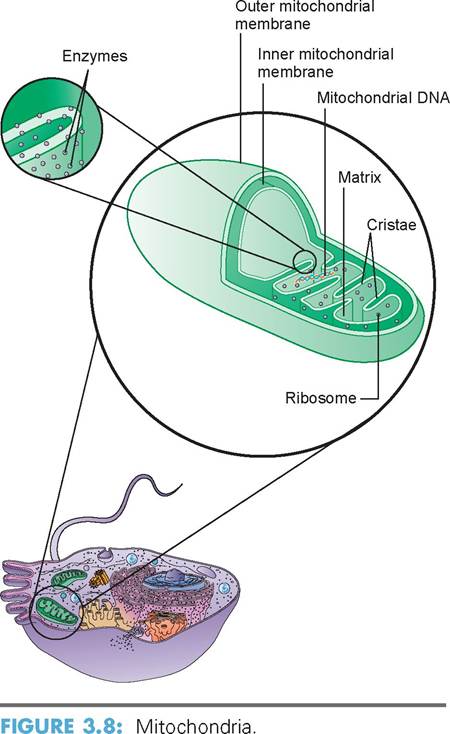
f. mitochondria: The mitochondria are the powerhouses of the cell because of their ability to produce ATP. A cell may have as few as 100 of them or as many as several thousand. Cells that have high physiologic activity, such as those in the muscles, liver, and kidneys, have many mitochondria because of their need for energy in the form of ATP. Mitochondria are found within the cytoplasm, where the need for energy is greatest, such as between the contractile proteins of muscle cells. Structurally, a mitochondrion is oval, like a small sausage. It is bounded by two membranes, similar to the plasma membrane. The outer mitochondrion layer is smooth, but the inner membrane is arranged in a series of folds called cristae. The center of the mitochondrion, bounded by the inner membrane and cristae, is fluid-filled and called the matrix. The cristae’s structure provides a large inner surface area in the mitochondria for the chemical reactions in the aerobic phase of cellular respiration (Figure 3.8). The enzymes that catalyze these reactions are located on the cristae and in the matrix. To reiterate, the main product of these reactions is ATP.
Mitochondria also can selfreplicate, as often occurs during times of increased cellular demand. Each mitochondrion has multiple identical copies of circular DNA. The genes formed by this DNA, along with the genes of the nucleus, control the production of proteins that build the mitochondrial components. Because there are also ribosomes inside the mitochondrial matrix, some protein synthesis also occurs within the inner membrane. In most cells the genes of the nucleus contain two sets of DNA, one each from the male and female parent. However, mitochondrial genes are inherited only from the female parent. This is because the head of the sperm (the only part that penetrates and fertilizes the ovum) normally lacks most organelles, such as mitochondria, ribosomes, endoplasmic reticulum, and Golgi complexes.
g. centrioles: The paired bodies of centrioles are cylindrical and are located close to the nucleus in all animal cells capable of reproducing themselves. They lie at right angles to each other. Internally, each cylinder is composed of nine triplets of microtubules, called a
9 + 0 array, because there are nine sets of microtubules in a circle without any in the center. The centrioles direct the formation of the mitotic spindle during cell division. They also form the basis for cell projections called cilia and flagella. A centrosome is the pericentriolar area plus the centrioles (Figure 3.9).
h. cytoskeletal elements: Inside the cell, within the cytosol, an internal scaffolding is formed that supports and moves substances within the cell. This structure is composed of several elements, including microtubules, which are slender tubules formed of proteins called tubulins. These tubulins have the ability to aggregate and disintegrate spontaneously. The centrosome serves as the
initiation site for the assembly of the microtubules. They grow outward from the centrosome toward the periphery of the cell. The microtubules aid in intracellular transport of organelles, such as secretory vesicles, and form the spindle for the migration of chromosomes during cell division. They also act in the process of transporting molecules down the length of an elongated cell, such as a neuron, and they help determine and maintain cell shape by providing rigidity to the cytosol. The intermediate filaments, another component of the cell’s internal scaffolding, are stable proteinaceous filaments that act as internal guy wires to resist mechanical (pulling) forces that act on the cells. Yet another component, microfilaments, are ribbon or cordlike elements within a cell formed from contractile proteins. Because they have the ability to shorten and lengthen (by relaxation), they are critical elements of cells that are mobile or cells that have the ability to contract, such as muscle cells. A cross-linked network of microfilaments braces and strengthens the internal face of the plasma membrane.
5. inclusions (inclusion bodies): Within the cytoplasm, various other substances and structures exist; these are stored foods (glycogen granules and lipid droplets), pigment granules, crystals of various types, water vacuoles, and ingested foreign materials. These are not active parts of the machinery of the cell; rather, they are passive masses and are therefore referred to as inclusions.
6. cilia and flagella: Cilia and flagella are motile, cell-surface projections composed of a 9 + 2 array of microtubule doublets (nine in a circle with two in the center) (Figure 3.10). Cilia are short and move fluids and debris over a cell’s surface, whereas flagella are long and can propel the entire cell.
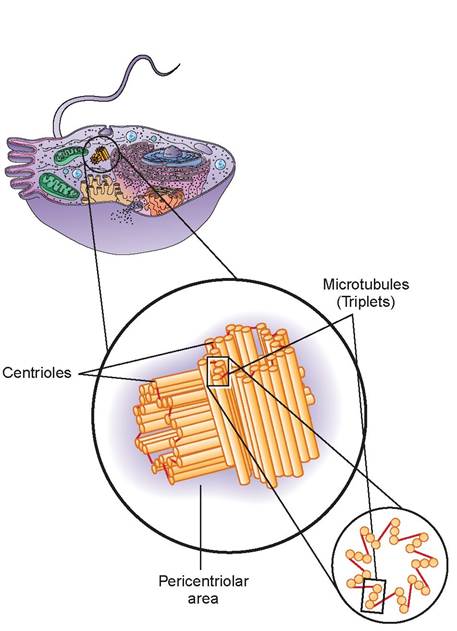
FIGURE 3.9: Centrosome.
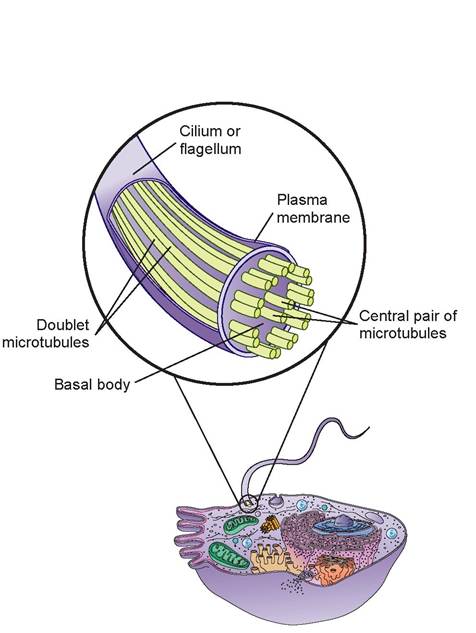
FIGURE 3.10: Cilia and flagella.
EXERCISE 3.1 IDENTIFICATION OF THE PARTS OF THE CELL
Using a model of a cell plus Figures 3.1 through 3.10, locate the following parts of a cell:
1. plasma membrane
2. nucleus
3. nuclear membrane
4. nuclear pores
5. nucleolus
6. chromatin
7. cytoplasm
8. cytoskeletal elements
9. ribosomes
10. rough endoplasmic reticulum
11. smooth endoplasmic reticulum
12. Golgi complex
13. lysosome
14. peroxisome
15. mitochondria
16. cristae
17. centrosome/centrioles
18. cilia
19. flagella
20. inclusion body
EXERCISE 3.2 IDENTIFICATION OF CELL PARTS USING A LIGHT MICROSCOPE
Follow the steps to complete the following procedure:
1. First, examine the giant multipolar neuron smear slide using the 10X objective lens on your light microscope. Locate a dark-blue, large motor neuron. Center the neuron in the field of view and change to the 40X objective.
Note that various parts of the slide take on different colors. We use various stains to help identify different cells or different parts of a cell according to color. A common stain is the hematoxylin and eosin combination stain. The hematoxylin tends to stain acidic portions of the cell dark blue or purple. These parts of the cell are called basophilic because they like or bond with basic stains. Because acid and base chemicals tend to bond together, cell parts high in RNA and DNA, such as the nucleus and nucleoli, attract basic stains and stain dark blue or purple. Using the same reasoning, eosin tends to stain basic portions of the cell pink or red. These stained parts are called acidophilic areas and include the more basic proteins within the cell, which are located within the cytoplasm. Other stains are more specific and may stain blue only the RNA, for example, as is the case with the cell you have on your microscope.
Note that at the cell boundary there is a distinct separation between the blue and the pink background. This, of course, is where the plasma membrane is located, but the details of this membrane are not visible using the light microscope. There are multiple cytoplasmic extensions, called cell processes, branching off the neuron’s cell body. It is for this reason it is called a multipolar neuron. Some of these processes were cut off during the sectioning of the tissue. Each cell, if sliced perfectly down the middle, would have visible one very long process, called an axon, and numerous shorter ones, called dendrites. At the center of the cell body is where the nucleus is located. Because the stain used for this slide stains the RNA darker, the nucleus (with mostly DNA) has a halo appearance around the RNA-rich nucleolus. Therefore, the dark dot in the center of the nucleus is the nucleolus. The nuclear membrane is either not visible or barely visible (Figure 3.11).
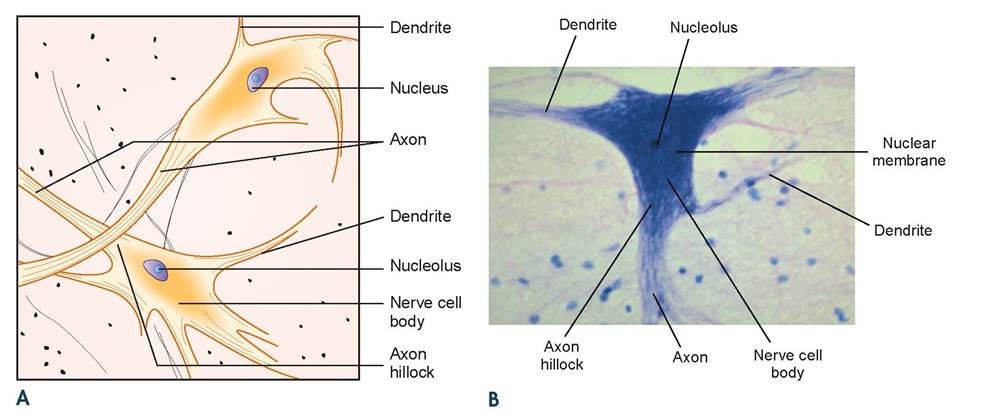
FIGURE 3.11 A and B: Illustration and photomicrograph of nerve cell body with processess.
In the space below, draw and label the parts of the neuron.
Think of the boundary of the circle below as the boundary of the microscope’s field of view.


Clinical Significance
There are numerous diseases specifically attributed to faulty cellular structure and function. Glycogen storage disease is a congenital absence of lysosomes, which permits accumulation of glycogen in cells. Vitamin A intoxication occurs when lysosomal enzymes are released to the exterior of cells, resulting in breakdown of the material between the cells. Cystic fibrosis is an inherited disorder that results in the production of a defective protein that fails to reach the plasma membrane. Because of its absence, the plasma membrane is unable to excrete chloride ions from the cell. This results in a thick mucus buildup outside the cells. In the lungs this causes difficulty in breathing, and elsewhere it results in improper secretion of pancreatic enzymes for digestion.
Summary
Because all pathological processes are ultimately the reflection of cellular dysfunction, understanding the inner workings of the cell is crucial to understanding how and why cells and tissues break down and cause disease. To understand pathology, it is first necessary to understand normal physiology. Hence, we study the structure and function of the cell and its organelles. This chapter began by explaining the structure of the cell's plasma membrane; then it discussed the nucleus and its internal structures. Finally, it addressed the structure and function of each individual organelle.
REVIEW QUESTIONS
1. What is the difference between an organelle and an inclusion body?
2. What is responsible for the plasma membrane’s antigenicity?
3. Name the two basic ways molecules move through the plasma membrane.
4. What structures control the movement of substance between the nucleus and the cytoplasm?
5. Where are the ribosomes assembled in the cell?
6. What structures synthesize proteins that are mainly used inside the cell?
7. What makes smooth endoplasmic reticulum so functionally diverse even though it lacks ribosomes?
8. What are the dilated ends of the sacs of the Golgi complex called?
9. What ion is actively transported into a lysosome, and why is this transport needed?
10. In what two organs are peroxisomes found in abundance, and why?
11. What molecule does the mitochondria produce in abundance?
12. Why is the DNA in the mitochondria different from the DNA of the nucleus?
13. Where is the mitochondrial DNA located in this organelle?
14. What is the difference between a centrosome and a centriole?
15. Name the three cytoskeletal elements.
16. Which of the three cytoskeletal elements contains contractile proteins?
17. Fill in the blank: Foreign material ingested into a cell is known as an.
18. What is the meaning of a 9 + 2 array of microtubules of cilia and flagella?
19. What is the chemical composition of the microtubules?
20. Why is the nucleolus so rich in RNA?