Cellular biochemistry
The cell theory, namely the idea that cells only arise from other cells, was first proposed in the late 1800s. This now seems elementary, but in the 1600s, Robert Hooke based it on initial observations of plant samples, which showed that tissues were composed of cells.
This was followed by studies by the German scientists Schleiden and Schwann, who insisted that all living things were composed of cells. This idea flew in the face of the notion of spontaneous generation, which suggested that organisms arose from debris or other inert nonliving materials. Four ideas inherent in cell theory are as follows:NHj
Phosphate bonds, for example, with the C5 rarhrtn n÷ rihneo nr
tute life and living. Understanding physiology requires an appreciation of these other scientific disciplines. Be advised that our discussions of specific chemical reactions, biochemical events, or physical properties are no substitute for specific classes in physics, chemistry, or biochemistry. However, we do believe that a sincere, thoughtful study of physiology provides an opportunity to integrate the fundamentals gained from the study of these disciplines. What could be more interesting than learning how our bodies and those of our animals function?
Chemical bonds
Cellular activity is tied to the creation of complex macromolecules, which requires combining other simpler molecules and creation of new bonds, and catabolic activity to break the bonds in nutrient molecules to supply the building blocks for these new creations. Some of the energy derived from breaking chemical bonds is also captured for future use by the cells. Enzymes catalyze nearly all cellular chemistry. As you learned in general chemistry, attractive forces maintain orientations between atoms in molecules to produce chemical bonds.
Three major types of bonds are (1) hydrogen bonds, (2) ionic bonds, and (3) covalent bonds. In addition, Van der Waals forces, similar to hydrogen bonds, are important in maintenance of the structure of complex biological molecules where charge differentials aid maintenance of structural relationships. We have already discussed hydrogen bonds in our description of how water behaves (see Fig. 2.1). Hydrogen bonds are produced when a hydrogen atom is covalently bound to an electronegative atom. The most common examples are oxygen and nitrogen. When these groups are positioned close to other strongly electronegative atoms, this produces a kind of tug of war as the two electronegative atoms "fight" for dominion of the hydrogen atom. These reactions create attraction forces that link or bridge the molecules. Hydrogen bonds are particularly important as intramolecular forces that act to mold the three- dimensional shape of many macromolecules. Much of the folding of proteins that defines their tertiary structure depends on hydrogen bonds. Another example is the interaction between complementary strands of the DNA molecule. Despite the relative weakness of individual hydrogen bonds, they are collectively critical for normal cellular functioning.Ionic bonds are produced when the transfer of electrons from one atom to another generates attractive forces between atoms. This happens because the usual balance between + and - charges is lost and ions are formed. The atom that accepts electrons acquires a net negative charge and is called an anion. The electron donor, now called a cation, has a net positive charge. Atoms with opposite charges attract. This is the basis of the ionic bond. A commonly used example of ionic bonds is table salt or sodium chloride. As you may recall from general chemistry, sodium has an atomic number of 11 and therefore has only one electron in its third or outer valence shell. To achieve stability, the atom would have to acquire an additional seven electrons (the third orbit around the nucleus can accept eight electrons).
Stabilizing this outer orbit is more likely to happen by shedding the single electron so that the second orbit (filled) becomes the valance shell around the nucleus of the atom. Chlorine has an atomic number of 17. The outer valance shell needs only one electron to fill its valance orbit. When it accepts an electron, stability is increased, but the atom acquires a net negative charge and becomes an anion. As you would suspect, ionic bonds are common between atoms with one or two valance electrons (metallic elements, sodium, calcium, and potassium, for example) and elements with seven valance electrons (chlorine, fluorine, and iodine). The majority of ionic compounds exist as salts. When dry, they form highly organized crystals as a consequence of their ionic bonds. In aqueous environments, salts dissociate to produce ions. Many more common ions (Ca+2, Na+, K+, CE) are critical to normal cellular physiology because of their roles in regulation of activity of many enzymes (Ca+2) and significance in maintenance of polarity across cell membranes. Calcium, which is the most abundant essential mineral in the body, is especially important. It not only is critical for the activation of various cytoplasmic kinases, it is essential for muscle function, and in a more stable form (hydroxyapatite—Ca10(PO4)6(OH2)), it is a fundamental part of the inorganic, extracellular structure of bone. This likely is the reason why shed antlers or skeletal remains of animals are not found in rural areas. Other animals, especially rodents, use these materials to supply their mineral needs.About one-third of basal energy (ATP hydrolysis) is used to maintain an ionic gradient across the plasma membranes through the activity of Na-K ATPase protein transporters in the plasma membrane. Specifically, these proteins move Na+ and K+ against their concentration gradients (energy requiring) so that concentrations of intracellular Na are about 10-fold lower inside than outside the cell (~15 vs.
150mEq∕L) with the opposite for K+ ions (~14 vs. 140mEq∕L). Since these ions passively move down their concentration gradients, the ATPase pumps must be constantly active, somewhat like the bilge pumps of a boat that act to constantly remove water that leaks or splashes into the boat. The transporter couples sodium and potassium transport so that each action of the protein ejects 3Na+ out of the cell and carries 2K+ ions into thecell. Coupled with the fact that the membrane is slightly more permeable to K+ than Na+ under basal conditions, the action of the ATPase protein maintains the ionic and electrical gradient across the cell membrane. These actions combined with the accumulation of cellular proteins cause the production of an electrical gradient across most cells of about -40 mV (inside relative to outside). Many cells, especially nerve cells, take advantage of changes in ion concentrations or polarity for cell signaling. The activities of chemical and voltage-regulated gates for sodium and potassium (transporter proteins) are altered to explain the abrupt changes in membrane potential that occurs during nerve transmissions.
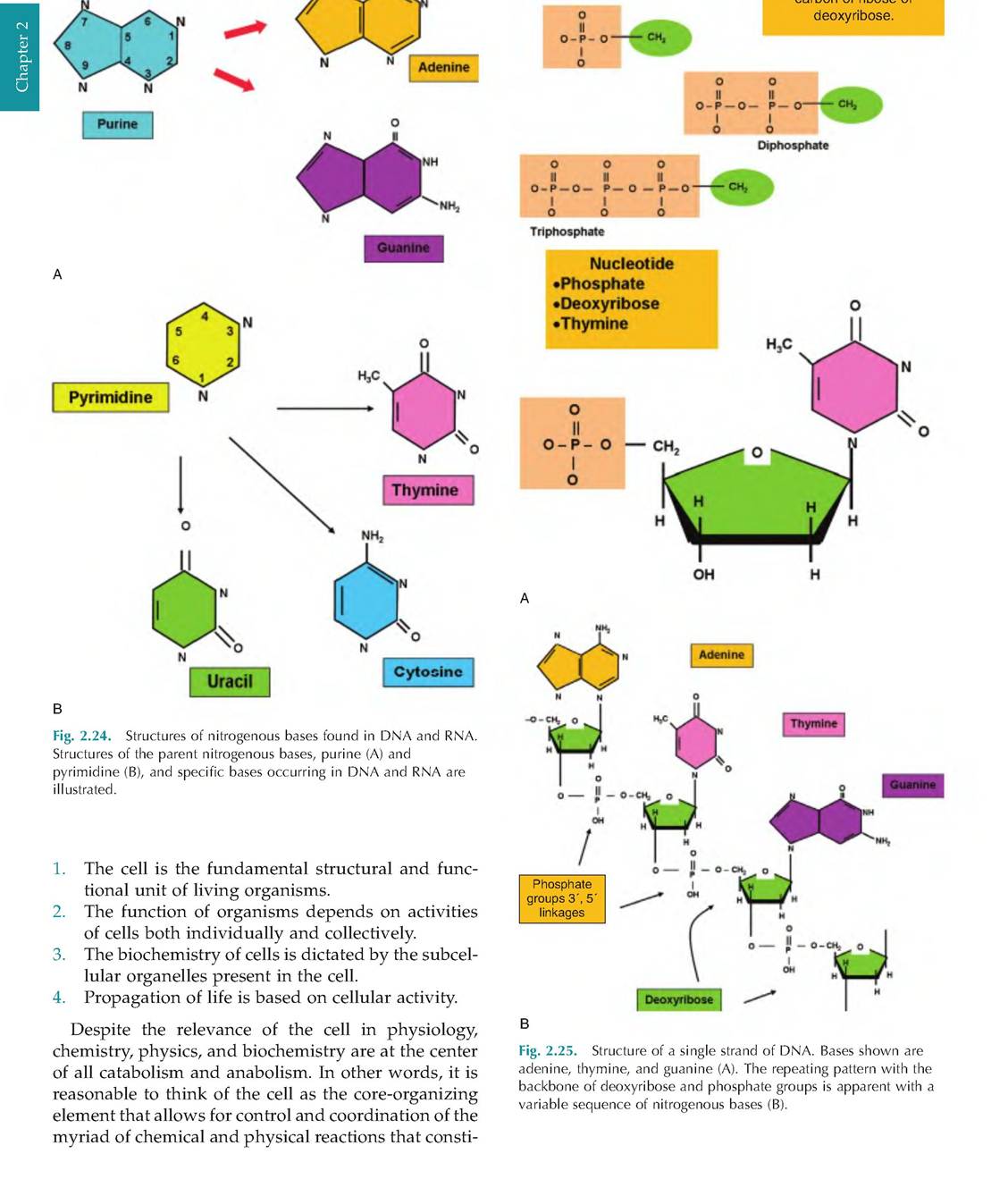
Aside from transferring electrons, atoms can also be stabilized by the sharing of electrons between nuclei. In these cases, the valence orbits of the sharing atoms are also shared so that stability of both atoms is achieved. This is the essence of covalent bonds. If a single pair of electrons is shared, this creates a single Covalentbond. For illustrative purposes, this is usually shown as a single line connecting two atoms. In other cases, atoms can share two or three electron pairs; this produces double or triple covalent bonds that are illustrated by double or triple lines between atoms, as (O=O) in O2 gas or (N?N) in N2 gas (see Fig. 2.18). Some atoms are considered relatively reactive and others relatively inert. As Figure 2.26 shows, a look at the atomic structure of some example atoms explains the reason for these differences.
Carbon is an especially abundant cellular atom. It has four electrons in its outer or valence orbit, but stability is achieved when the orbit is filled to capacity (eight electrons). This means that there are numerous possibilities of sharing electrons to achieve stability. One possibility is to create sharing of four electrons from four distinct neighbors. This would be the case in the creation of methane gas (CH4). Since hydrogen needs either to lose an electron or gain another to complete its outer valence orbital, sharing between the single carbon atom and four hydrogen atoms satisfies both atoms. Numerous examples of covalent bonds involving carbon atoms are shown in the structures of neutral lipids (essentially hydrocarbon chains), carbohydrates, and proteins (see Fig. 2.2, Fig. 2.18, and Fig. 2.20). By convention in drawings, carbon atoms are not explicitly shown but are understood to be positioned at line intersections. A simple example of double bonds is the sharing of electrons between a carbon atom and two oxygen atoms. In carbon dioxide (CO2 or O=C=O), sharing of electrons is equal between the atoms. Because of this, there is no separation of charge, so this covalent bond is also called a nonpolar Covalentbond. Nonpolar covalent bonds are common. For example, consider the fatty acid tails of the phospholipids in cellular membranes (see Fig. 2.3). Our
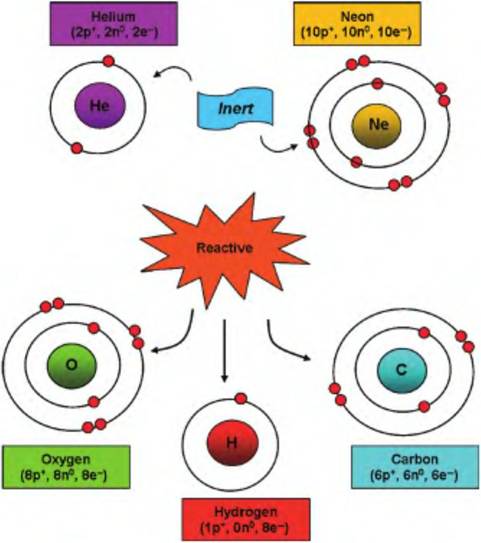
Fig. 2.26. Examples of chemically reactive and inert atoms. The red balls depict electrons in orbit around the nucleus of each atom. Helium and neon with completely filled valence orbits have little incentive to interact with neighboring molecules. In contrast, oxygen, carbon, and hydrogen need a change in the number of valence electrons to complete their outer shell of electrons and maximize stability.
prior discussion of water (see Fig. 2.1) illustrates an example of a polar covalent bond.
In these instances, like spoiled children, one or more atoms of the bond unit have a greater capacity to attract the shared electrons. Since the electrons spend more of their time in orbit near the stronger partner, these covalent bonds produce a separation of charge. In other words, on average, there is a separation of charge: one area of the new molecule has a net negative charge (shared electron(s) more often in this region) and a net positive charge (shared electrons more often "playing next door"). As a general rule, small atoms with six or seven valence electrons (oxygen, nitrogen, chlorine) are better able to attract electrons so are they are described as being strongly electronegative. These atoms favor "hogging" shared electrons to complete their valence orbital shells. Atoms with only one or two valence electrons are more likely to relinquish control of shared electrons and are called electropositive atoms. Examples of these include hydrogen, potassium, and sodium. If you ponder the difference between polar and nonpolar covalent bonds, it should be easy to imagine why molecules with an abundance of polar covalent bonds easily dissolve in (associate with) water. Conversely, molecules with few or any polar covalent bonds—lipids, for example—have little capacity to interact with the abundant polar water molecules in our cells and bodies.Chemical reactions
It is apparent that living processes depend on an almost bewildering array of chemical reactions. Molecules in our food or in the rations of our animals are destroyed (chemical bonds are broken) to supply intermediate molecules for building blocks, for example, consider hydrolysis of plant starches to supply glucose for production of glycogen in liver cells. Some of the glucose is oxidized to produce ATP needed to supply cellular energy. DNA and RNA are synthesized as our cells flourish and grow. How do we organize and make sense of all of these reactions? Fortunately, despite the number of individual molecules involved, patterns begin to evolve. For example, the breaking down of bonds between the glucose and fructose in the disaccharide sucrose or of the peptide bonds between alanine and lysine in a protein molecule are both hydrolysis reactions. Chemical reactions occur when chemical bonds are created between atoms, when chemical bonds between atoms are broken, or when chemical bonds are rearranged. A common illustration is to denote the reactions in simple symbolic expressions as chemical equations. For example, the combination of a carbon and oxygen to create carbon dioxide could be written in the following manner:
2O + C→CO2.
(reactants) (product)
This is an example of a synthesis reaction that requires the formation of new chemical bonds. This could be generally indicated by the following expression: A+ B → AB. Simplistically, such reactions explain the anabolic side of metabolism. Figure 2.18 shows an example of a synthesis reaction between two amino acids to create a dipeptide. The opposite of this is a decomposition reaction. These are often hydrolysis reactions because of the addition of water to complete the reaction. The expression AB → A + B gives a generic example of such a reaction. Specific examples would include the cleavage of glucose monomers from glycogen, the cleavage of peptides from a dietary protein by enzymes in the intestinal lumen, or the initial reactions of beta-oxidation—the process that acts to Catabolize two carbons at a time from fatty acid chains. These are all examples of catabolic reactions.
Another type of reaction is the exchange reaction. As the name suggests, in this situation, a functional group is moved from a donating molecule to a receiving molecule. This is indicated by these expressions AB + C → AC + B or AB + CD → AD + CB. Such reactions are common. For example, the nucleotides ATP or GTP are frequent donors of a phosphate group to another molecule. A very large family of enzymes called kinases catalyzes these phosphorylation reactions. As we will see in our discussion of cell signaling, many important regulatory enzymes are themselves controlled by their phosphorylation state. Some of these proteins are activated when a phosphate group is added and others are inhibited. Other examples also abound. For example, the first step in the catabolism of glucose in the biochemical pathway, glycolysis, involves the introduction of phosphate to produce glucose-6-phosphate catalyzed by the enzyme hexose kinase. Other exchange reactions transfer amino groups to allow generation of nonessential amino acids. For example, the enzyme alanine-pyruvate transaminase functions to promote the transfer of amino groups from most amino acids to generate alanine by adding the amino group to pyruvate.
A final class of reactions to consider is the oxidationreduction reactions. These reactions are central to catabolism of nutrients to manufacture ATP. Failure of these reactions, as occurs in the absence of oxygen, results in death. The essence of the reactions is the exchange of electrons between reactants. The molecule or atom that loses the electron (electron donor) is characterized as having become oxidized. The reactant that accepts the electron is said to become reduced. Many of these reactions involve the transfer of hydrogen (which of course includes the electron). These reactions will be considered in greater detail in our discussion of aerobic respiration.
Significance of enzymes
It is difficult to overstate the importance of enzymes in cellular metabolism. Without the presence of enzymes to catalyze the myriad of biochemical reactions required for anabolic and catabolic activities, life would cease. All enzymes are globular proteins that act as promoters of various but specific chemical reactions. Reactants must be present in the cell, but rates of reaction for virtually all biologically relevant processes are nonexistent if the required enzymes are absent or defective. As catalysts, enzymes accelerate reaction rates but do not become altered in the process. The enzyme can be thought of as a kind of biochemical matchmaker. Binding sites for substrates allow for close association of the substrates, a lowering of activation energy, and an enhanced opportunity that the reaction will take place. Because of the critical nature of the substrate-binding sites on the enzyme, even small changes in conditions can perturb the arrangement of amino acids around the binding site, rendering the enzyme ineffective (see Fig. 2.19). Because of
the unique nature of the binding site, enzymes are highly specific. For example, the enzyme hexose kinase, which adds a phosphate group to glucose, has no capacity to add a phosphate group to fructose, despite the fact that both molecules are similar hexose sugars.
Some enzymes are functional just as they are synthesized, that is, the protein alone. Other enzymes require the presence of a particular ion (often a trace mineral, e.g., Mg, Co, Cu, Mn) in their environment. These ions are thought to maintain the active-binding site of the enzyme in the appropriate conformation for reactants to be able to bind. These aids to enzyme function are called cofactors. Still other enzymes operate in combination with organic molecules called coenzymes that are essential for the enzyme to carry out its function. Coenzymes assist some groups of enzymes by acting as carriers of various specific functional groups or of electrons (hydrogen atoms). Many coenzymes are derived from vitamins, which explains the need for these substances to be present in the diet. For example, the B vitamins riboflavin (B2) and niacin give rise to flavin adenine dinucleotide (FAD) and nicotinamide adenine dinucleotide (NAD), respectively. These cofactors become reduced as a consequence of carbohydrate catabolism. When membrane bond enzymes in the mitochondria subsequently oxidize them, a portion of the energy associated with these electron transfers is utilized to drive the synthesis of ATP. Finally, some enzymes are composed of more than one protein. Some enzymes are first synthesized as pro- or precursor enzymes that only become active when they reach the area where they are to be used. For example, digestive enzymes synthesized by pancreatic cells are secreted into ducts where the enzymes are mixed with alkaline secretions and dumped into the lumen of the small intestine. The enzymes only become functional when they enter the lumen of the intestine. This is an important safety mechanism. Since these enzymes are capable of hydrolyzing a wide array of cellular proteins, inappropriate activation of the enzymes at the time of synthesis and secretion could cause degradation of your own pancreatic tissue. This is clearly undesirable.
Most enzymes are named based on the reactions they catalyze or the substrates upon which they act. For example, enzymes that cleave starch molecules are called amylases, those that cleave proteins are called proteases, and lipases break down lipids. Others are designated as dehydrogenases because they act to remove hydrogen atoms or decarboxylases for removal of carboxyl groups. Unfortunately, there are often multiple names applied to the same enzymes, and traditional names remain in usage. For example, the small intestine enzyme chymotrypsin is described as a hydrolase but more specifically as a proteinase and
Table 2.7. Enzyme classifications.
| Enzyme Class | Action | Example |
| Oxicloreductases | Oxidationreduction reactions (electronhydrogen transfer) | Lactate dehydrogenase |
| Transferases | Transfer of groups (other than hydrogen between substrates) | Hexose-6- phosphotransferase (hexose kinase) |
| Hydrolyases | Cleaves ester, peptide, and other types of bonds | Amylase |
| Lyases | Removes groups leaving behind double bonds | Aldolase |
| Isomerases | Interconversions of optical or geometric isomers | Phosphohexose isomerase |
| Ligases | New bonds to link together two substrates | Glutamine synthase |
further a serine proteinase because this defines the catalytic mechanism by which the enzyme cleaves proteins. The International Union of Biochemistry promotes a very systematic nomenclature for the naming of enzymes. However, in much of the physiological literature, common and general names for enzymes abound. Table 2.7 provides a six-level classification of enzymes along with examples.
Three steps occur in enzyme action. Step one is binding of substrates to the active site of the enzyme (formation of the enzyme-substrate complex). This binding induces a conformation change in the protein that allows rearrangement of the substrates and formation of the product (step two). The third step is release of the products from the enzyme. This happens because the newly created product no longer has a shape that is compatible with the active site of the enzyme. Enzymes function by reducing activation energy. Some chemical reactions can be promoted in the test tube by heating the reactants. This increases kinetic and thermal energy and makes collisions between substrates more frequent, thereby reducing the required activation energy. Lower activation energy in this manner in cells is not feasible. For example, one of the consequences of a high fever is the degradation of some proteins. By specific mechanisms that are poorly understood, binding of a substrate(s) to its specific enzyme lowers activation energy. The significance is profound. Compared with noncatalyzed conditions, rates of reaction are increased a million- or more fold. Just as remarkable, once the
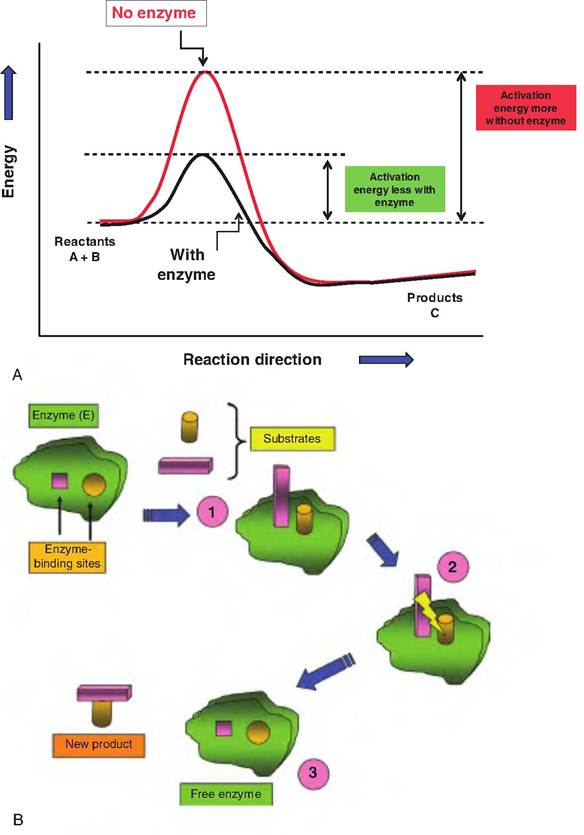
Fig. 2.27. Significance of activation energy. Panel A demonstrates the effect of lowering activation energy on the chemical reactions. Enzyme-catalyzed reactions are markedly enhanced. In panel B, the mechanism of enzyme action is depicted. Each enzyme is highly specific; only substrates capable of binding to the active sites will be affected. In step 1, the enzyme-substrate complex is formed. In step 2, internal rearrangements occur, and substrates are covalently linked. In step 3, the new product is released, and the enzyme is free for another round of activity.
product is released, the enzyme is free to repeat the process thousands of times. Indeed, some cellular enzymes remain in the cell for extended periods before they must be replaced. Figure 2.27 illustrates the idea of lowered activation energy and mechanisms of enzyme action.