Extracellular environment and cell function
Animal cells are continually bathed in extracellular liquid called interstitial fluid that is mostly derived from blood. In many ways, it is similar to blood plasma but contains few, if any, blood cells (some wandering leucocytes are sometimes present), and blood clotting factors are absent.
Other electrolytes, nutrients, and water are abundant. The movement of these compounds into and out of cells is controlled by both passive and active mechanisms. Health of the cells depends on the fact that the plasma membrane is Semipermeable and selective. This means that some substances are allowed to pass across the membrane but others are not. As you might guess, nutrients are readily captured, waste products are secreted, and many potentially harmful substances are excluded. The purpose of this section is to characterize some fundamental properties related to cellular transport and maintenance of the cellular environment withinappropriate boundaries. As noted previously, maintenance of normal concentrations of nutrients and ions in body fluids is an important part of homeostasis.
Osmosis
The diffusion of solvent—water, for example—through a selective or Semipermeable membrane is called osmosis. Water transport occurs when there is a difference in the relative concentration of water molecules across the membrane. If both sides of a water-permeable membrane are exposed to distilled water, there is no net move of water molecules. Just as many molecules would randomly move in either direction across the barrier. On the other hand, if a substance (solute) were dissolved in the water on one side of the membrane but not the other, the relative number of water molecules per unit volume would be decreased on the side of the membrane with the solute. If the membrane were impermeable to the dissolved substance, but permeable to the water, then water molecules would begin to leave the area with the higher concentration of water molecules and pass across to the opposite side.
Movement of solvent in this manner is called osmosis. Figure 2.28 illustrates the effects of membrane permeability on diffusion and osmosis. In the open system illustrated in the figure, differences in osmolality drive the movement of water molecules until hydrostatic pressure becomes sufficiently high to resist the further movement. Movement of water molecules to a region of greater solute concentration can be prevented by applying pressure. The pressure required to prevent migration of solvent molecules is the effective osmotic pressure of the solution.If we consider the cytoplasm of the cell (or the aqueous environments of many cellular organelles), there are multiple opportunities for varying concentrations of solutes and corresponding differences in osmolarity across these membranes as well as between the cytoplasm and the outside interstitial fluid. Since these membrane-bound compartments are permeable to water, osmosis has dramatic effects. For these reasons, it is important that the osmolarity of interstitial fluids and, consequently, blood, be maintained within relatively narrow boundaries. Changes in the osmolarity of the solutions bathing cells can cause cells to either shrink or swell. The capacity of a solution to affect the tone or shape of cells by modifying the internal water volume of the cell is called tonicity. A typical value for blood plasma or interstitial fluid is about 300mOsm∕L. Cells exposed to solutions with the same osmolarity or tonicity as blood plasma will retain their normal shape. Such solutions are described as isotonic. Examples include 0.9% saline (NaCl) or a
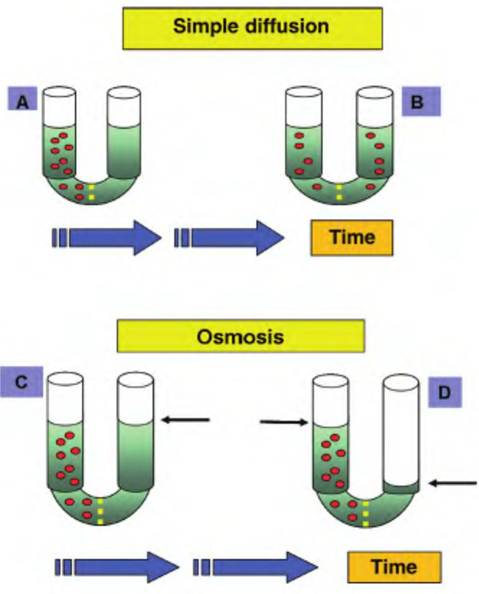
Fig. 2.28. Diffusion and osmosis. The upper panels illustrate the process of simple diffusion. At the beginning (A), two sides of a u-tube are separated by a membrane that is permeable to the solute (red particles) in the left arm of the tube. Over time (B), the solute molecules diffuse down their concentration gradient to areas of lower concentration.
With sufficient time, the solute molecules become equally dispersed throughout the entire solution. The lower panels demonstrate osmosis. Two sides of the u-tube are separated by a Semipermeable membrane that is permeable to the solvent (indicated by the green coloring) but not the solute (red) molecules (C). Since the right side of the tube has a greater concentration of solvent molecules, they move from the right side through the membrane. Migration continues until the hydrostatic pressure equals the osmotic pressure (D). Note the difference in the levels of liquid (arrows) after the experiment has continued for a period of time.5% glucose solution. These solutions are routinely used for preparation of intravenous treatments because the solutions have no impact on normal cell volume. Solutions with a greater effective osmotic pressure than blood are hypertonic and those with lesser effective osmotic pressures are hypotonic. Cells that are placed in a hypertonic solution lose water and shrink or crenate. Cells that are placed in dilute (hypotonic) solutions swell. As an extreme example, if cells are placed in distilled water, there are no dissolved substances, so water molecules enter the cells until the cells rupture. Figure 2.29 illustrates the responses of red blood cells incubated in isotonic, hypertonic, or hypotonic solutions.
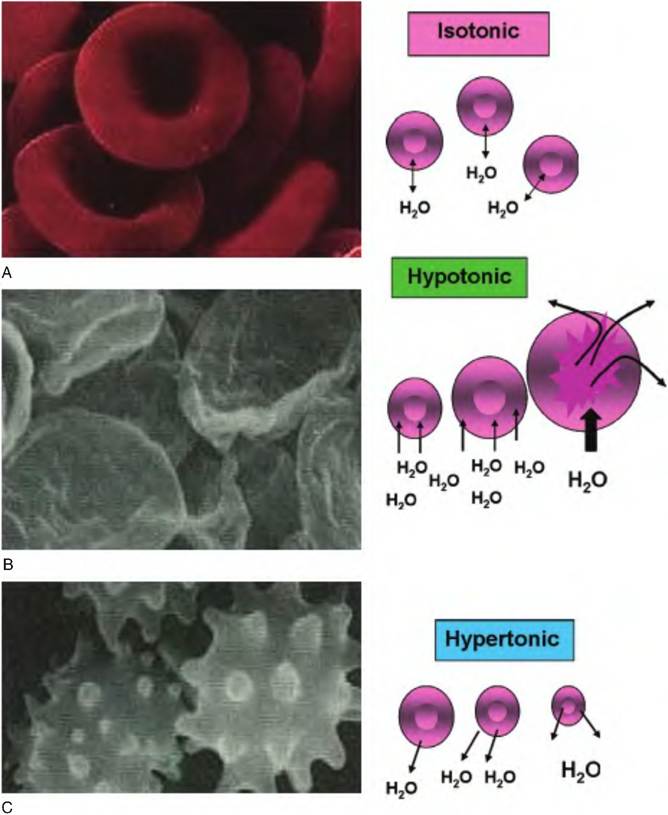
Fig. 2.29. Significance of tonicity. The consequence of placing red blood cells in solutions with differing tonicity values is illustrated. In panel A, cells are in an isotonic solution. Since the relative number of water molecules is the same inside and outside the cell, there is no net movement of water molecules and no change in cell volume. In panel B, cells are placed in a hypotonic solution. Here the number of water molecules outside is much greater than on the inside, so water enters and rapidly swells the cells. In fact, if the solution is sufficiently hypotonic, the cells will rupture.
This is shown on the left. These are remnants of red blood cell membranes. Panel C shows the effect of suspending cells in a hypertonic solution. Since the relative numbers of water molecules are greater inside, the cells lose water and they shrink. This is called Crenation and is characterized by the spikes of cell membrane shown on the left.Osmotic pressure (P) is related to temperature and volume, similar to the pressure of a gas as described in the following equation:
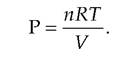
In this expression, n is the number of particles, R is the gas constant, T is the absolute temperature, and V is the volume. In usual physiological circumstances, R, T, and V are essentially constants. This means that the primary factor affecting osmotic pressure is the number of particles or solute molecules per unit of solvent. With ideal solutions, the osmotic pressure can be easily predicted. For example, if the solute under consideration ionizes completely, each molecule or particle is osmotically active. For example, Imol of NaCl per liter of water (58 g) would yield a 2 Osm or 2000 mθsm solution. This is because NaCl dissociates completely so that each Na and Cl ion exerts an osmotic effect. By contrast, 1 mol of glucose (180 g) per liter would produce a solution with an osmotic pressure of only IOsm or IOOOmOsm. For solutes that do not ionize, the molarity and osmolarity of the solution are the same. However, it is important to consider if the solute under consideration is penetrating or nonpenetrating relative to the cell membrane. If cells are
exposed to a 300m0sm solution of a penetrating solute, the solute will enter the cells, the relative water concentration outside the cell increases proportionally, and water then enters the cells. The solution effectively becomes more hypotonic over time. In addition, biological fluids are not ideal solutions. Interactions between molecules or partial dissolution of ionizing substances means that simply knowing the concentrations of various substances in blood, lymph, or urine, for example, cannot be used to accurately calculate osmolarity.
Fortunately, it is possible to measure osmolarity by indirect means. The temperature at which a fluid freezes is directly related to the osmolarity of the solution. Thus, the effective osmotic pressure of even complex solutions can be determined by measuring freezing point depression.Transport mechanisms
Passage of molecules into and out of cells and across organelle membranes occurs by either active or passive transport. For passive mechanisms, there is no direct energy expenditure by the cell. Active mechanisms, in contrast, require metabolic energy. Table 2.8 provides a listing of the membrane transport processes.
As illustrated in Figure 2.28, simple diffusion is the tendency for molecules to disperse evenly throughout a solution. Substances that are small, nonpolar, and
lipid soluble readily pass across cell membranes. Examples are oxygen and carbon dioxide. Since oxygen concentrations are virtually always higher in the blood and interstitial fluid than in the cytoplasm, oxygen continually diffuses into cells. Similarly, carbon dioxide is nearly always higher inside the cells, as a consequence of metabolic activity, so carbon dioxide is continually diffusing out of the cells. Most watersoluble molecules cannot simply diffuse through the cell membrane. This is because these polar molecules cannot "dissolve" in the fatty acid tails of the phospholipids of the lipid bilayer. However, if the substance is small enough, it can pass through the plasma membrane by passing through water-filled pores. Integral membrane proteins that span the entire width of the cell membrane create these pores. This means that substances sufficiently small and polar can essentially diffuse across the membrane by interacting with the water associated with these membrane-spanning proteins. Some of the pores are always available but others can be regulated. For example, in the cells of the kidney tubules, specifically the collecting ducts, permeability of the cells to water uptake by osmosis is variable.
This reflects changes in the conformation of these pore-forming proteins. When antidiuretic hormone (ADH) concentrations are elevated, the pores are widely available so that much of the water in kidney filtrate is recovered. When secretion of ADH is inhibited, more of the pore-forming proteins acquire a conformation that impairs water movement so urineTable 2.8. Summary of transport processes used by cells.
| Transport Type | Energy Source | Description | Example |
| Passive | |||
| Simple diffusion | Kinetic energy | Movement of molecules down a concentration gradient | Transfer of oxygen, some ions |
| Facilitated diffusion | Kinetic energy | Like simple but molecule bound to a carrier | Transfer of glucose into cells |
| Osmosis | Kinetic energy | Basically simple diffusion of water via Semipermeable membrane | Water transport |
| Filtration | Hydrostatic pressure | Movement of water and solutes through a Semipermeable membrane by a pressure gradient | Transfer of water and solutes from capillaries |
| Active | |||
| Active Transport | ATP | Move a solute against a concentration gradient | |
| Exocytosis | ATP | Secretion of cellular products via fusion of membranebound secretory vesicles | Secretion of milk proteins and lactose |
| Phagocytosis | ATP | So-called cell eating ameboid-like engulfing of extracellular materials by pseudopodia | Removal of foreign debris or dead cells by macrophages |
| Bulk-phase endocytosis | ATP | So-called cell drinking, involves the appearance of depression of the plasma membrane and progressive incorporation of fluid-filled vesicles | Occurs essentially in all cells |
| Receptor-med iated endocytosis | ATP | Ligand binding to surface receptors causes clustering of bound receptors in clathrin-coated pits which are then taken into cells for processing by lysosomes | Uptake of cholesterol and iron; downregulation of protein hormone receptors |
Box 2.2 Practical anatomy and physiology
The plasma membrane is a barrier to the extracellular space. It was long believed that water simply passed across cell membranes by diffusion. However, realization that water transport varied dramatically in different membranes, for example, proximal tubular cells versus cells of the ascending loop of Henle in the kidney, lead to the discovery of proteins called aquaporins (AQPs). These membranespanning proteins are especially abundant in cells such as those in the kidney nephron and are believed to be long sought-after "water channels or pores" hypothesized in the past.
The first of these proteins was identified in 1992 (Benga, 2012). There are now 13 different members of this family of proteins in humans, as well as many analogous proteins in plants, yeast, bacteria, and amphibians. Skowronski (2010) reported the expression of AQPs 1, 5, and 9 in the pig uterus during the estrous cycle and early pregnancy, thus suggesting that changes in AQPs are involved in changing the uterine environment to support reproduction.
production increases, and the urine that is produced is more dilute (Box 2.2).
Many important nutrient molecules, glucose, and certain amino acids are too polar to dissolve in the lipid bilayer for diffusion and too large to pass into the cells by passage through the plasma membrane pores or channels. These substances nonetheless pass into the cell by diffusion because they can interact with carrier proteins located in the plasma membrane. Precise mechanisms for this transport process are not well understood. But it is generally believed that binding of the substance to be transported causes a conformational change in the protein carrier that acts (1) to shield the molecule from interactions with the lipid bilayer and (2) to simultaneously move the molecule across the membrane. With the change in conformation, the carrier protein no longer can bind the molecule so it is released to the inside of the cell. This type of transport is called facilitated diffusion because substances transported still must pass down their concentration gradients but presence of the carrier proteins is required to mediate the transport. Unlike with simple diffusion, there are limitations. First, the carrier proteins themselves are typically highly specific. For example, carrier proteins that facilitate the diffusion of glucose would not be expected to also transport ribose. Secondly, the process can be saturated. This is essentially the reason why diabetic animals have glucose in their urine. Under normal conditions, a combination of facilitated diffusion and active transport is capable of removing and recovering all of the glucose molecules that appear in the kidney filtrate. However, concentrations of glucose in the blood and consequently the urinary filtrate of an animal with diabetes can be many times higher than normal. Since the number of transport proteins in the membrane of the kidney tubule cells is limited and the rate of transport is finite, not all of the glucose can be removed before the fluid passes out of the nephron and into the renal pelvis. This is an example of saturation. Simply stated, when all of the binding sites are occupied, the rate of transport is maximized. This rate of transfer, called the Vmax, is unique for each class or type of carrier protein. The capacity of the carrier to function is also affected by the relative ability of the binding site on the carrier protein to attract and hold the molecule for transport. This attraction is described as affinity. Carrier proteins with high affinity for their transport partner are able to sequester, bind, and transport their partner molecules even when concentrations are very low. Estimating the Km or binding constant for this interaction provides a quantitative measure of affinity. The Km is the molar concentration at which half of the carrier proteins are occupied. The lower this value, the greater the affinity. Interestingly, similar aspects apply related to the binding of hormones to their specific receptors.
Carrier proteins may also be affected somewhat like enzymes. Competitive inhibitors are able to block binding of the ligand (molecule to be transported). These inhibitor molecules essentially compete for the same binding site. In some cases the competing molecule is transported, but in other cases it is not. Some inhibitors can bind to regions other than the ligandbinding site of the carrier protein and induce a change in conformation that then makes the usual binding site incapable of interacting with the transport molecule. These effects are similar to the actions that serve to modulate enzyme activity. Carrier proteins that facilitate the transport of a single molecule or class (single binding reaction) across the membrane are called uniporters or uniport transporter proteins. Their Vmax and Km values determine the rate of reaction for these carriers. Other transporters are more Complicatedbecause the transfer of one solute depends on the simultaneous binding and transport of another molecule. If the molecules to be transported both move in the same direction, the carrier is called a symport (something like a symbiotic relationship), but if the molecules move in opposite directions, the carrier is called an antiport. Clearly, the kinetics that affect the actions of symport and antiport carriers are more complex than for simple uniport carriers. As an example, consider the uptake of glucose. Most animal cells have uniport carriers that take up glucose from the extracellular fluids where its concentrations are usually higher than in the cytoplasm. This is passive transfer that depends on facilitated diffusion. However, intestinal and kidney cells continue to take up glucose even when concentrations are low. These cells actively sequester glucose across their membranes via a symport carrier linked with the diffusion of Na into the cell. In other words, the movement of Na down its concentration gradient allows for the coupled transport of glucose into the cell. While cellular energy is not directly needed, maintenance of an effective concentration gradient for Na does require energy. Figure 2.30 illustrates differences between simple diffusion and carrier-mediated diffusion as well as theoretical models of three types of carrier proteins.
Filtration is the physical process whereby water and solutes are forced through a membrane or a capillary wall by simple hydrostatic pressure. Despite the non- Selective nature of this type of transport, it is critical for many physiological functions. Creation of urinary filtrate or movement of fluids across capillary beds throughout the body demonstrates physiological filtration. Filtration is considered a passive process, but it is obvious that pressure in the cardiovascular system depends on continued cardiac activity and muscle tone of arteries.
Hydrolysis of ATP and release of chemical bond energy that fuels transport of substances across membranes is active transport. Mechanisms are variable but fall into two broad groups. The first, like facilitated diffusion, depends on specific membrane carrier proteins. The second is the formation and uptake of vesicles. Carrier-mediated transport is specific but, unlike diffusion processes, moves molecules against their concentration gradients. In primary active transport, ATP is used to phosphorylate the carrier protein. Binding of the phosphate group to the protein alters its conformation. This causes the transport molecule to be shuttled across the cell membrane and released. One of the best studied of these carriers is the sodium-
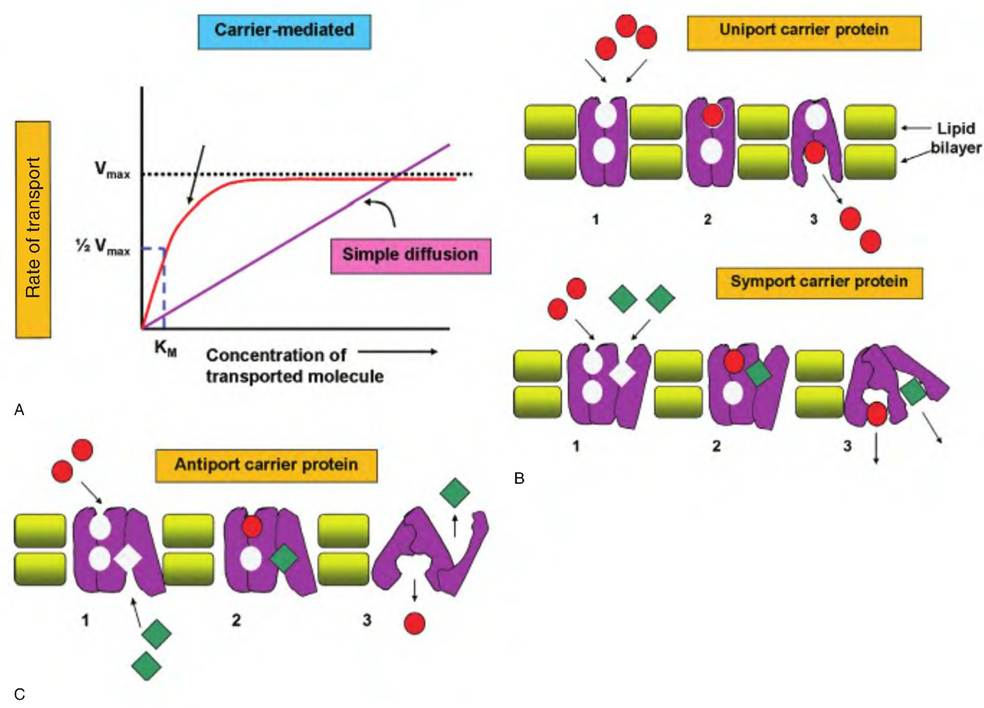
Fig. 2.30. Carrier protein-mediated transport properties. In panel A, the kinetics of simple diffusion compared with carrier-mediated transport are shown. For simple diffusion, the rate is directly proportional to the concentration of the transported molecule. Carrier-mediated transport shows a maximal rate (Vn,aJ regardless of further increase in concentration. The concentration at 1/2 Vmax is an estimate of the binding affinity (Km). Panel B illustrates the action of a uniport carrier protein. Step 1 shows binding of the transport molecule to the protein, followed by a change in carrier conformation (2), and then release (3) of the molecule to the inside of the cell. Symport carriers follow a similar pattern except that solute molecules (red and green symbols) bind and are transferred simultaneously in the same direction. In antiport carriers, transport of one molecule depends on the simultaneous binding and transfer of another molecule in the opposite direction.
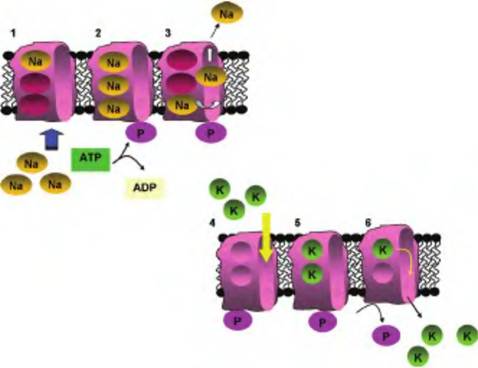
Fig. 2.31. Action of ATPase pumps. In step 1, Na ions from the cytoplasm bind to three sites on the protein. In step 2, complete loading stimulates the hydrolysis of ATP and phosphorylation of the protein. Phosphorylation causes a conformation shift and unloading of Na ions to the outside of the cell (step 3). Unloading of Na exposes two K binding sites available to K ions from the extracellular space (step 4). Complete loading of K ions (step 5) triggers release of the phosphate group and release of K ions to the inside of the cell (step 6). Once K ions are released, the protein returns to the conformation as in step 1 and is ready for loading of Na ions once again.
potassium ATPase pump. The carrier in this instance is a membrane-bound enzyme that cleaves ATP and functions to maintain steep gradients for Na (~10-fold higher outside) and K (~10-fold higher inside) relative to the plasma membrane. These ion gradients are essentially for normal signaling in nerve cells and initiation of muscle contractions. Another example of active transport is closely linked with muscle activity. When muscle contraction occurs, the cytoplasm of the cell is flooded with calcium. To regulate contraction events, calcium ions are pumped uphill back into storage inside the sarcoplasmic reticulum or back out of the cell in some situations. The working of the sodium-potassium ATPase pump is illustrated in Figure 2.31.
In addition to its relevance maintaining Na and K gradients and osmotic balance, the sodium-potassium ATPase pump is also responsible for transport of a number of important molecules. This is called secondary active transport. With greater concentrations outside the cell and the fact that the inside of the cell has a net negative charge, there is a continuous electrochemical gradient that promotes the diffusion of Na ions back into the cell. This is a bit like filling a pond behind a dam. Open a sluice and water pours out. The energy of the water flow can be used to spin a turbine to produce electricity or, as in the 1800s, to turn grinding wheels to produce corn meal or flour. The cell can take advantage of energy from this downhill diffusion of Na by linking it with the uptake of other important molecules. For example, recovery of
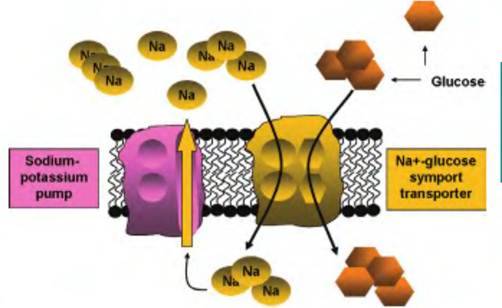
Fig. 2.32. The sodium-potassium ATPase pump and sodiumglucose symport. As detailed in Figure 2.31, the Na-K pump maintains a gradient so that concentrations of Na are higher outside the cell. As Na ions diffuse down their electrochemical gradient, the inside of the cell also has a net negative charge, and movement of Na is linked with the simultaneous transfer of glucose into the cell (transporter on the right). With respect to glucose uptake, this is sometimes called secondary active transport, since ATP is not directly required for action of the glucose-Na+ symport, but it is needed to create and maintain the Na+ gradient across the cell membrane.
glucose and some amino acids against their concentration gradients can be achieved in this manner. This is illustrated in Figure 2.32. Ion gradients are critical for the actions of many different membrane transport systems. Some of these are similar to the symport for glucose. For example, other specific symport carriers also linked with the electrochemical gradient for Na+ cotransport uptake of some amino acids and other sugars. Ion gradients are also important in the operation of the antiport carrier that operates in the membrane of cells in the gastric pits in the stomach to regulate the pH of gastric juices by secretion of HCl.
In addition to the transport involving specific membrane carrier proteins, substantial numbers of molecules associated with the cell surface or molecules simply positioned near the surface can be moved into the cells by either pinocytosis (engulfment of fluids) or endocytosis (engulfment of particles). Both of these processes require energy. Control of endocytosis is complex and poorly understood. In some cases, the process is receptor mediated. That is, the binding of a specific ligand to proteins on the cell surface initiates a cascade that leads to endocytosis of the receptor and associated ligand. This process often begins in areas of the membrane containing called clathrin-coated pits. These endocytosed receptor proteins can be processed in at least three different ways, depending on the particular receptor-ligand combination and cellular conditions. Multiple vesicles are believed to fuse to create a larger membrane structure called an endosome, from which smaller vesicles bleb or bud. A common fate is for receptors to be either recycled to the same region of membrane (recycling), moved to another area of
membrane (Iranscytosis), or shuttled to lysosomes for destruction. As for pinocytosis, it is generally believed that essentially all cells continually ingest bits of their plasma membrane along with fluids via the continuous generation of these very small vesicles.
Specialized blood-related cells, especially neutrophils and macrophages, actively undergo phagocytosis. This process is familiar from our images of ameba surrounding food particles. In similar fashion, these leukocytic cells send pseudopods around cell debris or bacterial cells and engulf the material. The ingested material is captured in a membrane-bound body called a phagosome. Most often, this is followed by fusion with lysosomes to degrade the material. Interestingly, in some cases, protein fragments from the degraded material, especially in the case of foreign substances, are returned to the cell surface so that other leucocytes can detect it. Macrophages that carry out this activity are called antigen-presenting cells. This process allows other immune cells to generate memory cells and initiate the production of antibodies against the foreign invader. Figure 2.33 illustrates types of endocytosis and the trafficking associated with endocytosis.
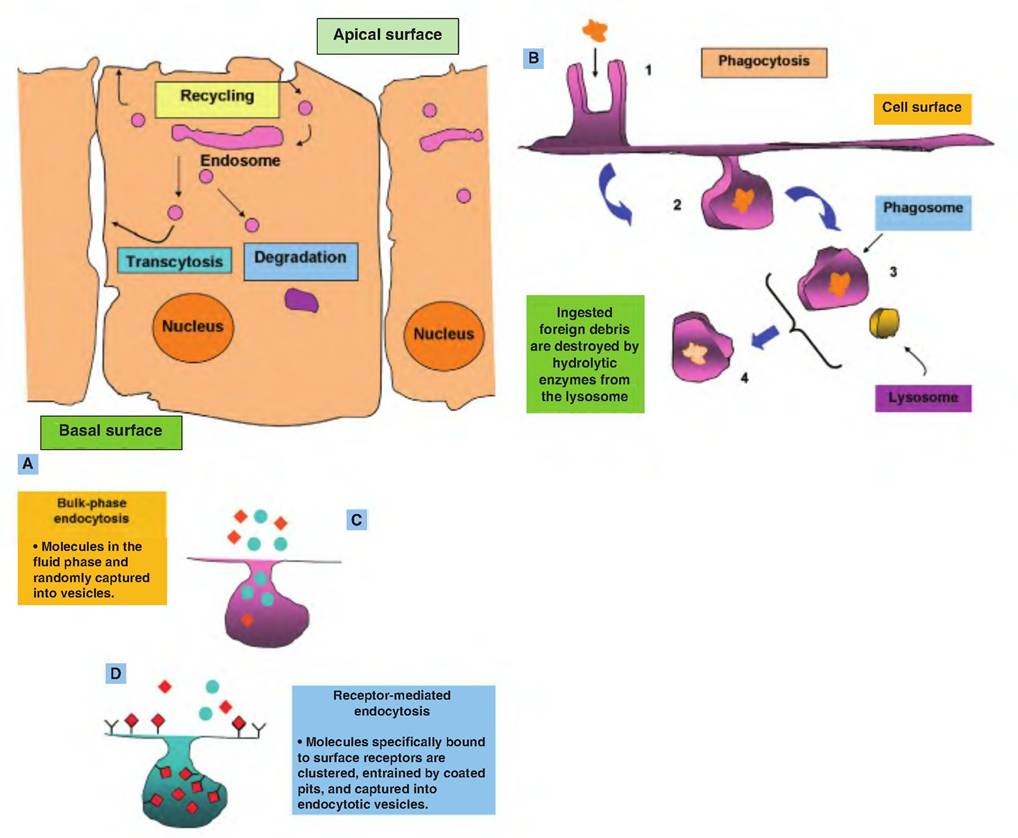
Fig. 2.33. Processing of vesicles derived from receptor-mediated endocytosis. Processing of vesicles derived from receptor-mediated endocytosis is illustrated in panel A. Once vesicles enter the cell they fuse to produce the endosome. Vesicles that bleb from the endosome are subjected to one of three fates. Many of the receptor proteins are returned to the plasma membrane to be reused at the original site (recycling) or in a different location of the plasma membrane (transcytosis). Other vesicles along their proteins (receptors and/or ligands) are degraded by lysosomal enzymes following fusion. Panel B illustrates a specialized type of endocytosis (phagocytosis) that is employed by certain white blood cells. Macrophages, for example, identify cellular debris or perhaps bacterial cells for engulfment. Cytoplasmic extensions called pseudopods extend and surround the targeted particle (1). The material is engulfed and captured in a vesicle called a phagosome (2). Fusion between the phagosome and lysosomes (3) lead to destruction of the captured material (4). The next two panels (C and D) illustrate differences between pinocytosis and receptor-mediated endocytosis. Pinocytosis is more random since the cell has no control over specific molecules that can be secured.
More on the topic Extracellular environment and cell function:
- Extracellular environment and cell function
- Contents
- Akers R. Michael, Denbow D. Michael. Anatomy and Physiology of Domestic Animals. 2nd edition. — Wiley-Blackwell,2013. — 685 p., 2013
- FGF family
- Cell Physiology
- Epithelial cell junctions
- Cellular Anatomy and Morphology
- Chapter summary
- Anatomy of the Cell
- Kidney excretion of wastes and control of pH