Kidney excretion of wastes and control of pH
As we learned in earlier sections, maintenance of blood and extracellular fluid pH within relatively narrow boundaries is critical for homeostasis. Movement of carbon dioxide from tissues and the bicarbonate buffering system of blood is a critical component in maintenance of pH.
Clearly, the ratio of ventilation rate to the rate of CO2 production determines the body concentration of CO2. Changes in ventilation rate therefore have an important role in modulation of pH, especially in the short term. However, the capacity to regulate excretion of CO2 via the lungs and excretion of acid via the kidneys combine to provide long-term control of blood and extracellular fluid pH. In particular, the excretion of acid or H+ ions in the urine is an important aspect of maintaining the plasma bicarbonate (HCO3 ) concentration in mammals. It is worth remembering that normal digestive and metabolic processes produce large quantities of acids that must be buffered and/or excreted.At this point, a brief reminder of some pH and buffering fundamentals is in order. Since all functional proteins (enzymes, receptors, etc.) are influenced by pH, it follows that acid-base balance is critical for homeostasis. When the pH rises above the optimal value (~7.4 for most animals), the situation is referred to as alkalosis; if pH drops below pH 7.35, this is described as acidosis. Because in a chemical sense a pH of 7.0 is neutral, a pH of 7.35 is not, chemically speaking acidic, but the hydrogen ion concentration is higher than optimal for the animal. So for most animals, any arterial pH between pH 7.35 and 7.0 is deemed physiological acidosis.
While all animals ingest small amounts of acidic substances, most H+ are derived as by-products of metabolism. Consider the breakdown of amino acids from proteins or fatty acids from triglycerides or the buildup of lactic acid.
Concentrations of H+ in blood and consequently extracellular fluids are controlled by (1) buffer systems, (2) respiratory centers in the brain stem, and (3) the kidneys. The chemical buffers act virtually instantaneously to prevent dramatic swings in pH. Changes in respiration rate and depth of respiration begin to compensate for either acidosis or alkalosis within a matter of minutes. The kidneys have very potent effects, but these changes require hours or perhaps days before they are fully effective.The chemical buffers act by converting strong acids or bases into weaker acids or bases. The point to remember is that strong acids or bases dissociate rapidly and completely so that the change in free H+ ion concentration can be dramatic. Weaker acids or bases dissociate less effectively so changes in H+ ion or OH concentration are reduced. Important buffer systems in the body include the bicarbonate buffer system, phosphate buffer system, and the protein buffer system.
The respiratory system regulation of H+ ion concentration revolves around the elimination of carbon dioxide generated by cellular respiration. Briefly, CO2 that enters the circulation combines with water to create carbonic acid. It then dissociates to produce H+ and bicarbonate ion as illustrated in the following:
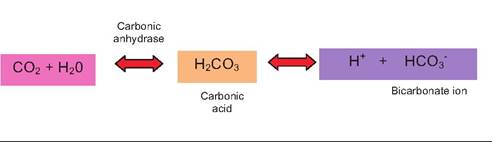
In reality, most of the CO2 that enters blood through the capillary beds in transit for exhalation in the lungs also passes through the red blood cells. Much of the H+ that is liberated from carbonic acid in the creation of bicarbonate ion is captured by oxygen-depleted hemoglobin (Hb). CO2 is also carried by depleted hemoglobin (Carbaminohemoglobin). Bicarbonate ions diffuse out of the red blood cells in exchange for chloride ions from the plasma. The situation is essentially reversed as the blood gets to the lungs.
Oxygen entering the red cells displaces the protons from the Hb, and CO2 enters the plasma. Carbonic anhydrase in the membrane of the endothelial cells also converts some of the bicarbonate into CO2.The various chemical acid-base buffer systems in the body are essentially pairs of molecules that act to resist changes in H+ concentration when a strong acid or base is added to the system. The three systems are the bicarbonate, phosphate, and protein buffer systems. The idea of protein buffering was illustrated by the capacity of Hb to capture protons coming from carbonic acid production. The bicarbonate buffer system is especially important in the extracellular fluid compartments and is a mixture of carbonic acid (H2CO3) and its sodium salt (NaHCO3). In this combination, if a strong acid (HCl) is added, bicarbonate ions of the salt act as a weak base to capture much of the H+ produced from the dissociation of HCl. This produces more of the weaker carbonic acid. Again, since it is a weak acid, it dissociates only modestly so the pH of the solution is lowered much less than would have been expected. In a similar way, if a strong base (NaOH) were added, more of the carbonic acid reacts with the NaOH to produce more of the weak base (sodium bicarbonate) and water. The net result is that the pH of the solution rises only modestly. These reactions are illustrated as follows:
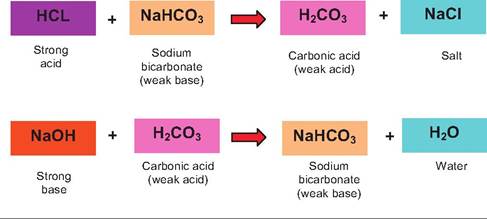
The phosphate buffer system operates in a similar fashion. Its components are the sodium salts of dihydrogen phosphate (H2POy) and monohydrogen phos-
phate (HPO∣2 ). NaH2PO4 acts as a weak acid, while Na2HPO4, acts as a weak base. The phosphate buffer system is only in low concentrations in the extracellular fluids, but it is an important buffer in urine and within intracellular compartments where phosphate concentrations are typically higher.
As illustrated in the following, hydrogen ions from strong acids are captured by converting a weak base to a weak acid and strong bases countered by conversion of a weak acid to a weak base.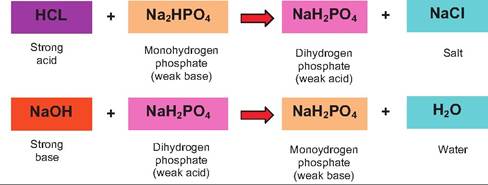
Proteins located in the blood plasma and intracellular proteins are also critically important buffering elements (protein buffering system). Because of their abundance, proteins account for most of the buffering capacity of the body. As we discussed in earlier chapters, proteins are chains of amino acids. Some of the amino acids have reactive or free carboxyl residues (so-called organic acid groups [-COOH]), which dissociate to release H+ ions when the pH begins to rise. Other amino acids have reactive groups that can act as bases to accept protons. For example, an exposed NH2 group can bind a hydrogen ion to become NH3+. This action effectively removes free hydrogen ions, which prevents the solution surrounding the protein from becoming too acidic. The same proteins can function in a reversible manner either as acids or bases depending on the pH of their environment. Molecules with this capacity are called amphoteric molecules.
These buffer systems can effectively capture excess hydroxyl or hydrogen ions in the short term, but they cannot eliminate excess acids or bases from the body. The lungs can dispose of carbonic acid by exhalation of CO2, but the kidneys are critical in eliminating most of the other acids produced by metabolic activity. These include phosphoric, uric, and lactic acid. Calling these metabolic or fixed acids is sometimes used to make a distinction between carbonic acid derived from CO2 and these other metabolic acids. For example, acidosis that is caused by the accumulation of these "metabolic" acids is sometimes called metabolic acidosis to distinguish it from acidosis that can be caused by a failure of the lungs to eliminate sufficient CO2.
This failure can produce an accumulation of carbonic acid; thus, acidosis is referred to as respiratory acidosis. Clearly, both types of acidosis depend on alterations in metabolism, but sources of the problem can be distinguished.The kidneys, then, are critical for long-term regulation of body pH because of their ability to compensate for acid-base imbalances caused by changes in diet or disease. One of the most important aspects is the capacity of the kidney to either conserve (reabsorb) or produce new bicarbonate ions as well as the ability to excrete bicarbonate ions if required. The concentration of HCO1 in the plasma of most mammals is about 25 ? 10,∖4∕L and H+ ion concentration about 40 ? 10'j∖4∕L. Since the composition of the filtrate that enters the nephrons is about the same; there are relatively large amounts of bicarbonate but very much less hydrogen ion. However, urine has a pH of about 6.0 with a minimal amount of bicarbonate ions. As we discussed, metabolic processes demand that excess acid produced from catabolic events must be excreted. So how is this accomplished? The goal is to add excess H+ to the filtrate to be lost in urine with little bicarbonate. If you consider the operation of the bicarbonate-carbonic acid buffer system described earlier, losing a HCO3- from the system pushes the equation to the right and effectively increases H+ ion concentration. In the opposite manner, the creation or reabsorption of HCO3- from the kidney filtrate is the same as losing H+ as the equation is pushed to the left. To summarize, to reabsorb bicarbonate, H+ ions are secreted, but when excess HCO3- is excreted, H+ is retained.
Let's first consider the events that allow for excretion of H+ into the filtrate (remember, this is in addition to H+ that was initially filtered; you might also recall that urine is typically slightly acidic, ~6 compared with blood plasma).
Hydrogen ion excretion occurs primarily within the PCT as well as so-called type A cells of the collecting ducts. The hydrogen ions to be secreted arise from the dissociation of carbonic acid. However, as in the red blood cells, the carbonic acid is produced within the tubular epithelial cells from the diffusion of CO2 from the peritubular capillaries and the action of carbonic anhydrase, which combines water and CO2 to produce carbonic acid. As each H+ ion is excreted, a Na+ ion is reabsorbed. This maintains the electrochemical balance across the tubular cells. In general, the rate of H+excretion varies with changes in the CO2 content of the peritubular blood. Since blood CO2 is directly related to blood pH, these systems can respond to adjust to either rising or falling pH. High CO2 (corresponding with low pH) would lead to a larger diffusion gradient for movement of CO2 from the peritubular blood into the tubular cells. This would be followed by increased production of carbonic acid, its dissociation, and therefore excretion of more H+. Some of this excreted H+ can combine with urinary bicarbonate ions (HCO3 ) to produce CO2 and water. Some of this urinary filtrate CO2 can also enter the tubular cells and promote further excretion of H+. A reduction in blood CO2 would produce the opposite effect.The A cells are located primarily in the DCT and CD and serve as acid-secreting cells. These cells have a proton pump in the apical membrane and a chloridebicarbonate exchange system in the basolateral membrane. The cells also have high concentrations of carbonic anhydrase, which serves to combine CO2 and H2O making protons and bicarbonate ions available. The H+ ions are moved across the apical membrane (facing the filtrate) and the bicarbonate ions transported across the basolateral membrane into the surrounding interstitial fluid. The H+ ions that are secreted can react with bicarbonate ions in the filtrate to create carbon dioxide and water, which can diffuse back into the cell. The net result of this process is that activated type A cells can produce a positive uptake of bicarbonate into the blood through the recycling of CO2 while adding protons the filtrate. Thus, the A cells are acidsecreting cells (Figure 16.19).
The activity of the A type cells and therefore the degree of acid secretion can be regulated. For example, in periods of acidosis, the activity of the cells is stimulated. This is believed to involve direct changes in the proton ATPase activity (increases synthesis of transporter proteins and/or activity) as well as increased activity or synthesis of the basolateral membrane bicarbonate-chloride exchange proteins. In this way,
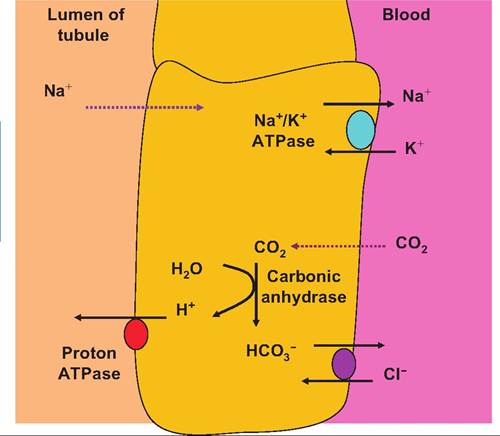
Fig. 16.19. Acid-secreting type A cells. A-type cells pump protons into the filtrate of the nephron by action of an apical H+ ATPase. This acidifies the filtrate so that the potential favors reabsorption of Na+ ions. The intracellular Na+ concentration is reduced by the activity a basolateral Na7K+ ATPase pump.
increases in blood bicarbonate would serve to counter the acidosis. For example, aldosterone is known to stimulate the activity of the proton ATPases.
As H+ secretion by the tubular cells continues, the pH of the filtrate drops, and consequently, continued secretion is against an increasing concentration gradient. The ability to secrete more protons decreases with decreasing pH (higher H+ concentration) until, at a pH of about 4.5, acid secretion stops. However, more protons can be secreted if needed if the acidic filtrate can be buffered. Depending on conditions, bicarbonates, phosphates, and ammonia can accomplish this mission. Hydrogen ions can react with bicarbonate to form carbon dioxide and water, with HPO∣2 to produce H2POy, or with NH3 (ammonia) to produce ammonium ions (NH4+). Thus, the kidney has three systems (bicarbonate, phosphate, and ammonia) to act as buffering agents. The cells of the nephron are nearly impermeable to both ammonium ions and phosphates so that these agents effectively capture excess hydrogen ions for excretion from the body. The phosphates that appear in the filtrate are derived when filtration of the blood takes place in the glomerulus; ammonia, on the other hand, diffuses from the blood into the tubular cells where it gets converted into the less toxic ammonium ion. The availability of phosphate for buffering is dependent on the diet.
Under acidosis conditions, plasma bicarbonate concentrations can fall so that the buffering capacity of the renal filtrate is also reduced. When this occurs, ammonia can become a major pathway for elimination of excess acid. Ammonia is synthesized in the tubular cells by the enzymatic deamination of amino acids. The amino acid glutamine is especially important in this process. Ammonia is formed routinely in hepatic cells, but it is rapidly converted to less toxic urea and glutamine. Effectively, ammonia is carried to the kidney cells in the form of glutamine, where its deamination produces ammonia. In its nonpolar, unionized form (NH3), it readily diffuses across the cells into the filtrate to combine with H+ to yield the highly polar ammonium ion (NH4+). Because of its polarity, it is essentially trapped in the filtrate so that it is excreted in the urine. This pathway therefore allows for the excretion of excess nitrogen as well as excess hydrogen ions.
Bicarbonate ions are also clearly critical in the bicarbonate buffer system. If this reserve of base is to be maintained, then the kidneys must not only act to counter rising H+ concentrations by excreting more H+, they must also either recover HCOy ions that have been filtered or secreted or create additional bicarbonate ions. This involves a convoluted process because the tubule cells are nearly impermeable to HCOy ions present in the filtrate. A second type of cells, the B type cells, function as base-secreting cells. These cells are
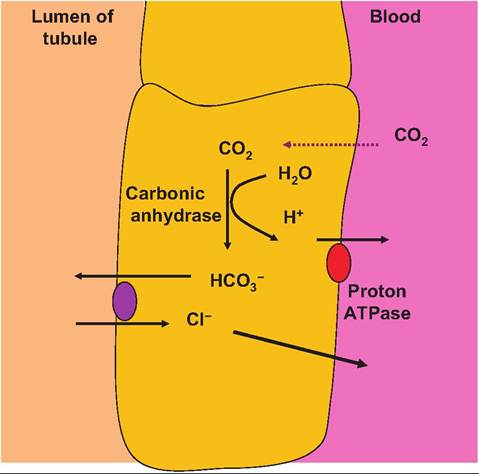
Fig. 16.20. Base-secreting type B cells. B-type cells use the proton ATPase proteins in the basolateral membrane to pump protons into the blood in concert with recovery of chloride ions.
arranged so that they express a proton ATPase transporter in the basolateral membrane and a chloridebicarbonate exchange protein in the apical membrane. These cells also express carbonic anhydrase. The result is that bicarbonate is secreted into the filtrate while protons are recovered (Fig. 16.20).