Epithelial cell junctions
Along the adjoining borders of epithelial cells there are specialized cell junctions. These are regions or sites where some special contact between cells can be recognized. Three functional classes of junctional complexes include (1) occluding junctions, (2) anchoring junctions, and (3) communicating junctions.
Some of the communicating junctions, for example, the gap junction, also appear in other cell types. However, because of the importance of epithelial tissue to create tissue compartments or barriers, it is important to understand the role of cell junctions in this process. To illustrate the idea of barriers, consider the differences between milk and blood. It is clear that blood circulates throughout the mammary gland within the capillaries just underneath the secretory epithelial cells of the mammary alveolus (see Fig. 4.8). However, the composition of blood or interstitial fluids and milk is very different. The same can be said for the environment of the gut lumen versus the interstitial fluid of the Iacteals of the villi of the intestinal tract. How are these differences developed and maintained? This is where junctional complexes come into play.In circumstances where it is necessary to maintain a seal between epithelial cells, the lateral margins of the cells become fused together along a system of membrane ridges between adjacent cells. These ridges extend completely around the perimeter of the cells to create a sort of belt located near the apical ends of the cells. These junctional complexes are called zonaoc- cludens or tight junctions to indicate that they produce an effective barrier. For example, during the latter stages of mammary development in the pregnant heifer, the approach of parturition signals both the structural differentiation of the alveolar cells and maturation of tight junctions between the cells.
Increases in circulating glucocorticoids along with declining progesterone seem to be especially important. Once this occurs, paracellulartransport (transport of components between the cells) is dramatically reduced. This creates an effective blood-milk barrier so that transfer of serum components into milk or milk constituents into blood is minimized. This does not mean that transport cannot occur but that wholesale leakage is prevented. The effectiveness of this barrier function is readily apparent from study of secretions obtained from animals with acute mastitis or experimental treatments known to disrupt the tight junctions. One of the effects of this disruption is the appearance of serum proteins in milk, for example, albumin. Conversely, these situations also allow abrupt increases in the appearance of lactose and Oc-Iactalbumin (and likely other milk components) into serum.Adhering junctions are a second class of membrane specializations that act to anchor epithelial cells
together. These complexes also appear as bands or belts that circumnavigate the perimeter of the cells below the level of the tight junctions. In intestinal epithelial cells, for example, these complexes are called zona adherens. As you traverse along the lateral membrane toward the basal end of the cell, a second type of adhering junction, the desmosome or macula adherens, appears. Anchoring junctions are widely distributed and allow the epithelium to maintain structural integrity by linking cells together and by linking cells to the underlying extracellular matrix. These complexes are plentiful in tissues that are subjected to mechanical stress, for example, skin. The adherens junctions are focal points where actin filaments attach to the junctional proteins. In general terms, there are two basic parts of these complexes. The intracellular attachment proteins create a plaque or thickening on the cytoplasmic side of the cell membrane and provide sites for attachment of cytoskeleton proteins and for transmembrane linker proteins.
The transmembrane linker proteins have cytoplasmic tails that attach to the plaque structure, but the extracellular domains of the proteins interact with the extracellular domains of adjacent junction proteins or with other extracellular matrix proteins,that is,hemidesmosomes. In the case of the adhesion belts in epithelial sheets, the complexes in companion cells are directly apposed. The transmembrane linker protein is a member of a family of Ca2+-dependent proteins called cadherins. The plaque or adhesion belt through the actions of several linker proteins (catenin, vinculin, and others) binds bundles of actin fibers that radiate into the cytoplasm interacting with the cytoskeleton. It is thought that changes in the orientation and contraction of these bundles explain the folding of epithelial sheets to create tubular structures during tissue development.Desmosomes, unlike the bands of the zona adherens, are limited to spots or patches of the membrane between adjacent cells. They could be envisioned as spot welds or small dollops of glue to help bind adjacent epithelial cells together. To carry this analogy a bit further, the zona adherens could be thought of as miniature packing straps that bind the epithelial cells. Both the zona adherens and desmosomes are closely associated with microfilaments within the cytoskeleton of two cells that are linked. A variant of the desmosome, the hemidesmosome, has the structure of only half a desmosome. This complex serves to anchor the epithelial cells to the underlying basement membrane. For desmosomes, the transmembrane linker proteins also belong to the cadherins family of proteins, but the specific intracellular protein associated with the plaque varies. In most cells, these are keratin filaments, but desmin filaments fulfill the same function in cardiac cells.
A final type of structure is the gap junction. In this instance, proteins aligned in neighboring cells essentially create pores that pass from one cell to the other.
This can be imagined as small pipes passing between two adjacent apartments. Gap junctions appear not just in epithelial tissue but are prominent in cardiac muscle, some types of smooth muscle cells, and between cells of the nervous system. Gap junctions allow for the direct passage of small molecules (typically less than 300 MW) between cells. This is important for cell- to-cell communication and rapid responses necessary for nerve function and muscle contraction. We will discuss the specific physiological events related to cell junctions in subsequent chapters. A stylized view of cellular junctions is provided in Figure 4.32. Figure 4.33 illustrates the structure of the gap junction.Gap junctions are composed of transmembrane proteins called ConnexinsWhen arranged to create acom-
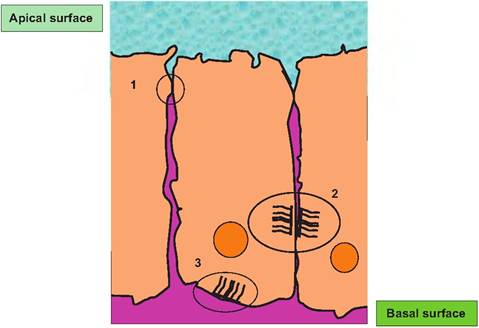
Fig. 4.32. Cellular junctions. Tight junctions (1) serve to effectively seal the apical from the basal surfaces of the epithelium. Notice that apical or basal molecules penetrate between the cells only to the region of the junction. Desmosomes (2) act to link cells together something like spot welds, while Itemidesmosomes anchor the cells to extracellular matrix molecules. Adhesion belts are not illustrated but would typically occur in the region just below the tight junctions.
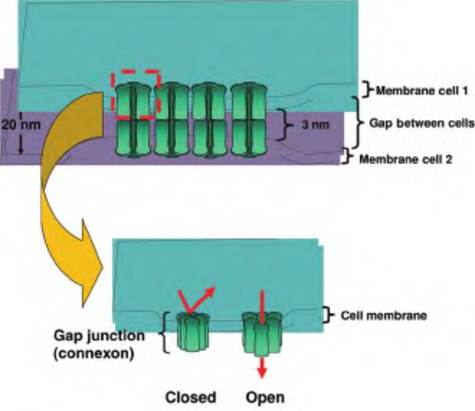
Fig. 4.33. Gap junction structure.
plex, six connexin proteins align to form a pore or channel called a connexon. As illustrated in the upper panel, when connexons of two adjacent cells become aligned, they create an aqueous pore that connects the two cells. However, unlike tight junctions, the outer leaflets of the adjacent cells are not fused. Gap junctions can also alternate between open and closed states. For example, a decrease in pH or an increase in Ca2+ concentrations promotes closure. Thus, the function of gap junctions can be regulated.