Cellular response
Acute cellular response
Once vascular dilation occurs, an acute cellular response to an inflammatory stimulus can begin. An early leukocyte response to inflammation is the increased release of neutrophils from the marginating pool in blood vessels and the storage pool in the bone marrow into the circulating pool in the bloodstream.
This can be transiently mediated by epinephrine and corticosteroids (Webb & Latimer, 2011a). Glucocorticoids also cause mild, transient lymphopenia and eosinopenia. After the initial response, inflammatory mediators (including ILs and colony-stimulating factors) promote the division and differentiation of progenitor cells in the marrow and drive a persistent leukocytosis. Typically, this response results in increased numbers of circulating mature neutrophils and monocytes. If the response is severe, band neutrophils may be released by the bone marrow, resulting in neutrophilia with a left shift (Figure 2.14). If the stimulus is overwhelming, a neutropenia may occur as the neutrophils are rapidly consumed at the site of inflammation and the marrow response to inflammatory mediators is unable to meet demand. An eosinophilia may be induced by parasitic or allergic inflammation, whereas a lymphocytosis can be seen with more chronic antigenic stimulation and viral infection. Leukocytes move from the bloodstream into inflamed tissues by adhering to endothelium then migrating by diapedesis between cell junctions in the capillary walls (Figure 2.15). They follow a gradient of chemical mediators to the site of an inflammatory stimulus. Adherence/adhesion of leukocytes to endothelial cells has a reversible and an irreversible phase. 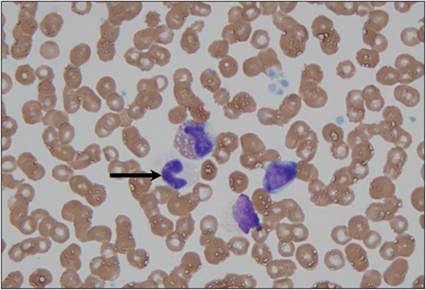
Figure 2.14 Immature granulocytes in circulation. Peripheral blood from a 10-year-old, intact male mixed-breed dog.
A band neutrophil (arrow) and a band eosinophil (upper center) are shown. A small lymphocyte (right), a lysed nucleated cell (lower center), platelets, and erythrocytes are also present (Wright–Giemsa, 1,000? magnification).
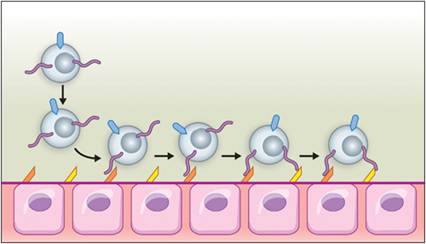
Figure 2.15 Diapedesis. During an inflammatory event, leukocytes are triggered to alter the expression of cell surface proteins and follow a gradient of chemical mediators to the site of an inflammatory stimulus. During this process, leukocytes adhere to endothelial cells. Purple lines indicate P-selectin glycoprotein ligand 1 (PSGL-1), expressed by leukocytes. Orange structures are endothelial P-selectins, which allow for leukocyte rolling on binding PGSL-1. Yellow structures are endothelial E-selectin, which slows leukocyte rolling. Teal structures on the leukocytes are β2-integrins, utilized in pavementing and illustrated in Figure 2.16. (Courtesy Veronica Kinn.)
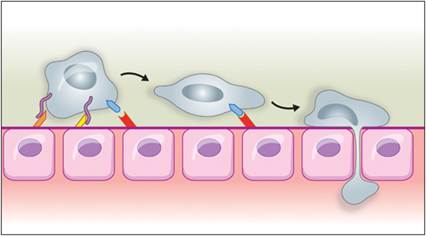
Figure 2.16 Pavementing is an irreversible process of leukocyte adherence to endothelial cells that leads to the migration of leukocytes between cell junctions in the capillary walls and toward a site of inflammation. It is induced by continued IL-1 and TNFα stimulation and mediated by β2-integrins (CD11/CD18) on leukocytes (teal structures) and intracellular adhesion molecule 1 on endothelial cells (red structures). (Courtesy Veronica Kinn.)
Reversible adherence occurs as vascular changes and inflammatory mediators cause the cells to marginate toward vessel walls. Blood flow through affected tissue is slowed by vasodilation, allowing leukocytes to interact with the endothelium and adhere to it. This is mediated by endothelial cell expression of P-selectin in response to histamine, thrombin, PAF, and other mediators. P-selectin is pre-formed in endothelial granules called Weibel–Palade bodies.
Within 2 or 4 hours, the endothelium expresses E-selectin in response to TNFα or IL-1, respectively (Wyble et al., 1997). Leukocytes express P-selectin glycoprotein ligand 1 (PSGL-1), which interacts with endothelial selectins. Binding of PSGL-1 to P-selectin initiates rolling. Binding to E-selectin slows down rolling (Choi et al., 2009). Leukocytes also express L-selectin, which recognizes mucosal addressin cell adhesion molecules on high endothelial venules in the intestine and other sites (Ogawa et al., 2005). Similarly, L-selectin recognizes glycosylation-dependent cell adhesion molecules in vessels of lymph nodes, mammary glands, the uterus, and lung (Hemmerich et al., 1994). If there is no further activation of the leukocytes or endothelium by mediators, then selectins are shed by the leukocytes and recycled by the endothelium, and the leukocytes return to circulation. If further stimulation occurs, an irreversible process called pavementing occurs. This is induced by continued IL-1 and TNFα stimulation and is mediated by β2-integrins (CD11/CD18) on leukocytes and intracellular adhesion molecule 1 on endothelial cells (Dimasi et al., 2013; Figure 2.16).Diapedesis begins when platelet/endothelial-cell adhesion molecule (PECAM-1) and junctional adhesion molecules (JAMs) on leukocytes bind to PECAM-1 and JAMs expressed within endothelial junctions (Dimasi et al., 2013). This occurs at postcapillary venules and sometimes capillaries. Neutrophils are particularly good at extravasation and they are the first to get into a site of inflammation. Monocyte/macrophage migration through the endothelium is partially dependent on initial neutrophil migration (Soehnlein et al., 2009).
Chemotaxis enables leukocytes to migrate to the site of an inflammatory stimulus. A chemoattractant binds to its receptor on the leukocyte cell membrane. This is followed by changes in membrane fluidity that allow the leukocyte to send out long pseudopodia. The front end of the cell then moves toward the stimulus, and the rest of the cytoplasm is pulled along by contraction of actin and myosin (Weninger et al., 2014).
The first cell types recruited to an inflammatory lesion are often granulocytes, specifically neutrophils and/or eosinophils. Basophils are also granulocytes, but they are rarely observed in an inflammatory response; if they are present, they are typically associated with chronic inflammation. Granulocytes, or polymorphonuclear cells, have a lobed or segmented nucleus and contain cytoplasmic granules with various characteristics, due to their contents. The staining characteristics of the granules are the basis for the names of the different granulocytes. Neutrophils contain three types of granules (azurophil granules, specific granules, and tertiary granules); all are neutral staining or nonstaining with Romanowski stains. Eosinophil granules take up eosins to stain bright red or orange. Basophil granules take up basophilic stains to stain dark blue. Granulocytes are terminally differentiated and have short half-lives. Granulocytes normally circulate in the blood with neutrophils being the most common, eosinophils less common, and basophils rare.