Neutrophils
Most types of injury cause the release of mediators that are chemotactic for neutrophils. The primary chemokine that draws neutrophils to a site is CXCL8 (IL-8). Large numbers of neutrophils may accumulate in one area and form pus.
Neutrophils are 10–12 μm in diameter and have a highly segmented nucleus (Figure 2.17). The number of nuclear segments can vary in segmented neutrophils. Neutrophils with fewer than two segments are considered immature, band neutrophils (Figure 2.18). Those with more than five segments are termed hypersegmented neutrophils, and are near the end of their life span (Figure 2.19). If not activated to migrate into tissues, neutrophils will die via apoptosis about 10 hours after release into the circulation. If activated to migrate into tissues, neutrophils will live for 1–2 days (Webb & Latimer, 2011b). When activated, neutrophils move rapidly by ameboid motion and are intensely phagocytic. 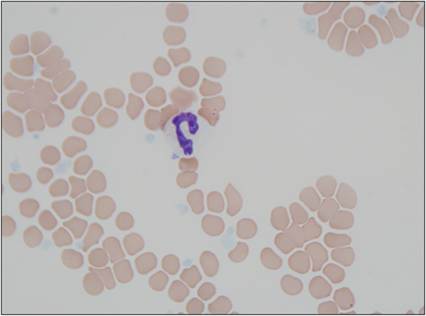
Figure 2.17 Segmented neutrophil. Peripheral blood from a 10-year-old, intact male mixed-breed dog. The neutrophil is mature with nonstaining cytoplasm and a segmented nucleus with dense chromatin (Wright–Giemsa, 1,000? magnification).
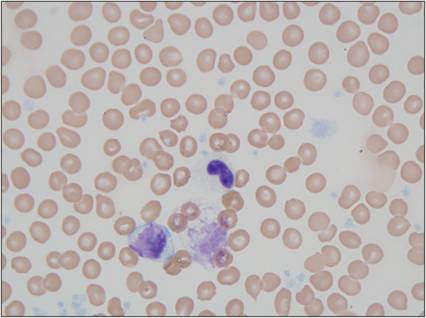
Figure 2.18 Band neutrophil. Peripheral blood from a 10-year-old, intact male mixed-breed dog. The neutrophil is immature with nonstaining cytoplasm that is speckled with small pink primary granules. The nucleus is horseshoe-shaped and has dense chromatin and a dark, round chromocenter (Wright–Giemsa, 1,000? magnification).
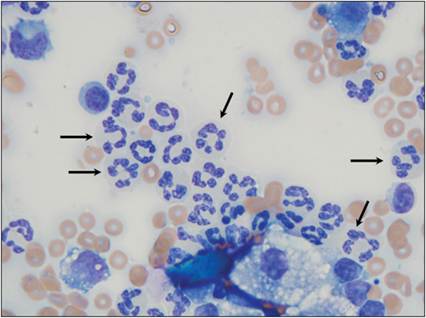
Figure 2.19 Hypersegmented neutrophils.
Synovial fluid from an 8-year-old, neutered male Labrador Retriever. Several neutrophils and lower numbers of large mononuclear cells are present. Neutrophils have segmented nuclei with crisp edges and dense chromatin. Neutrophil nuclei with more than five segments are considered hypersegmented (examples indicated by arrows) (Wright–Giemsa, 1,000? magnification).
A key function of neutrophils is to phagocytize foreign material or injurious agents and neutralize them (van Kessel et al., 2014). Neutrophils send out processes that surround and enclose the offending particle in a vacuole. They utilize Fc and C receptors to detect Ab–Ag complexes and complement proteins on the surface of a particle, respectively. These receptors are upregulated by inflammatory mediators. The phagocytized material stimulates a respiratory burst by the neutrophil. During this process, the phagocytic vacuole fuses with a neutrophil lysosome to begin degrading the particle. If the neutrophil cannot ingest the particle, it will release its lysosomal enzymes out onto the surface of the particle, which can inadvertently digest and harm the surrounding tissues. Elastases, collagenases, and gelatinase released by neutrophils are especially damaging because they destroy the stromal framework of the tissue.
The respiratory burst is essential for killing bacterial and fungal agents by the neutrophil. First there is a dramatic increase in respiratory activity due to an increase in O2 consumption. The enzyme NADPH oxidase in the membrane of the phagocytic vacuole converts O2 to O2 − (superoxide anion). This molecule is toxic and reacts with water to form H2O2. If iron is present, hydroxyl radicals are generated. Lactoferrin in the specific granules of neutrophils serves as a source of iron for this reaction. Azurophil and tertiary granules contain cytochrome b, which also generates free radicals. Additionally, myeloperoxidase (MPO) is present (mainly in azurophil granules) and converts Cl− ions to oxidizing agents.
Another way neutrophils eliminate pathogens is through the formation of neutrophil extracellular traps (NETs, Nauseef & Borregaard, 2014). NETs are comprised of DNA, proteins, and granule constituents that are released from neutrophils following NETosis, a type of programmed cell death for neutrophils, which occurs in response to neutrophil receptors binding PAMPs and certain inflammatory mediators. Neutrophils can either actively release NETs from their cytoplasm or can depolarize and rupture, subsequently releasing NETs (Papayannopoulos, 2018). These form a fibrillar matrix, which entraps bacteria, yeast, and other pathogens. Neutrophil extracellular traps are effective at killing bacteria. They also limit the spread of released granule constituents to minimize tissue damage.
Neutrophils also stimulate the inflammatory response by generating kinins, PGs, LTs, PAF, chemotaxins, and many cytokines. They cleave complement as well as activating mast cells and platelets to release histamine. Neutrophil granule proteins promote synthesis of monocyte-attracting chemokines by endothelial cells and macrophages. Dying neutrophils also attract monocytes/macrophages.