Eosinophils
Eosinophils are typically present in allergen- and parasite-mediated inflammatory reactions. Eosinophils are particularly good at activating mast cells. Another major function of the eosinophil is to regulate acute hypersensitivity reactions.
Eosinophils are 12–15 μm in diameter and have a segmented nucleus and several large, brightly eosinophilic, cytoplasmic granules (Figure 2.20). A major component of eosinophil granules is eosinophil major basic protein (MBP). This protein kills parasites by creating pores in the cuticle of the organism. Tissue damage by MBP is prevented by ingestion of the protein by mast cells. Reciprocally, MBP neutralizes heparin secreted by mast cells. Eosinophils also contain per-oxidases, eosinophil cationic protein, and other degradative enzymes (Acharya & Ackerman, 2014). 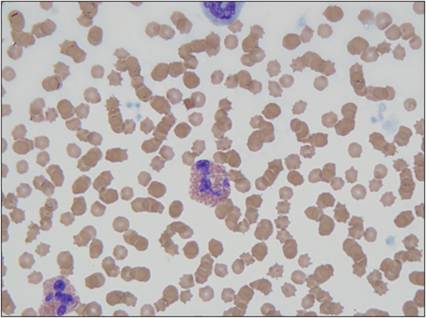
Figure 2.20 Eosinophil. Peripheral blood smear from a 13-year-old, spayed female domestic shorthair cat. Two eosinophils are shown, in the bottom left and in the center of the image. The cell has abundant cytoplasm, filled with eosinophilic granules. The nucleus is segmented and has dense chromatin (Wright–Giemsa, 1,000? magnification).
Eosinophils are attracted to inflammatory sites where cells are secreting IL-5, which is the major mediator affecting eosinophil production and activation. It is produced by mast cells, Th2 lymphocytes, and eosinophils. Eosinophils are ameboid and phagocytic, like neutrophils, but they are more selective in their responses to chemotactic stimuli. Eosinophils are stimulated by antibody–antigen complexes and histamine. However, they are not as responsive to complement. Eosinophils can reside in tissues for several days to weeks.
Large mononuclear cells
Monocytes, macrophages, and DCs are histiocytic cells.
Monocytes are present in the peripheral blood. Once they migrate through the endothelium, they begin to differentiate into macrophages. Macrophages migrate to the site of injury after neutrophils, partly because their migration is enhanced by the extravasation of neutrophils through the endothelium. Released neutrophil granule proteins anchor on endothelial proteoglycans and are adhered to by monocytes rolling along the endothelium (Soehnlein et al., 2009). Macrophages take about 24 hours to accumulate in the lesion. If the agent becomes persistent, then the macrophage will become a prominent cell type in the lesion. Unlike granulocytes, macrophages can enter a tissue site and divide there (Weiss & Souza, 2010). Macrophages regulate inflammatory responses and clean up damaged cellular and microbial debris, making them very important for preparing the site for healing and repair. DCs tend to reside in tissues and play a major role in directing the immune response to injury.Monocytes have an oval to bean-shaped nucleus and gray–blue cytoplasm with small vacuoles and occasional granules (Figure 2.21). These cells are 15–20 μm in diameter. Tissue macrophages have a more pleiomorphic morphology. They tend to have a large round to oval nucleus, abundant blue–gray cytoplasm, and vacuoles (Figure 2.22). Macrophages can be up to 80 μm in diameter and can be swollen and filled with phagocytized debris (Figure 2.23). They can have an epithelioid morphology, making sheets of macrophages difficult to differentiate from epithelial cells (Figure 2.24). Additionally, macrophages can fuse together and become multinucleated giant cells (Figures 2.25, 2.26). Multinucleated giant cells tend to occur when large, hard-to-digest foreign bodies are encountered or when there is chronic infection.
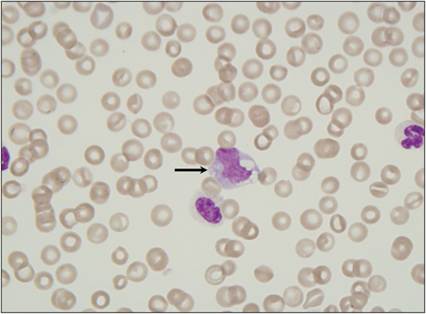
Figure 2.21 Monocyte. Peripheral blood smear from a dog. The monocyte has abundant basophilic cytoplasm that contains several distinct cytoplasmic vacuoles and has an irregularly shaped nucleus with slightly open chromatin (arrow) (Wright–Giemsa stain, 1,000? magnification).
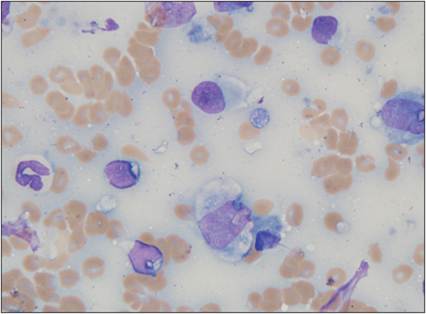
Figure 2.22 Tissue macrophage. FNA from a mass on the ear pinna of an 8-year-old, neutered male Boxer. A mixture of inflammatory cells was observed. A mildly degenerate neutrophil is shown at the left of the image. Three lymphocytes with scant cytoplasm and a smooth, round nucleus are present. The lymphocytes are smaller than the neutrophil. The three cells that are larger than the neutrophil are macrophages (two in the center and one to the far right in the image). Macrophages have abundant, lightly basophilic cytoplasm and a round to oval nucleus. The nuclei of macrophages typically have more stippled chromatin than lymphocyte nuclei. A small amount of eosinophilic material is observed within the cytoplasm of the largest macrophage. A few lysed nuclei and several erythrocytes are present in the background (Wright–Giemsa stain, 1,000? magnification).
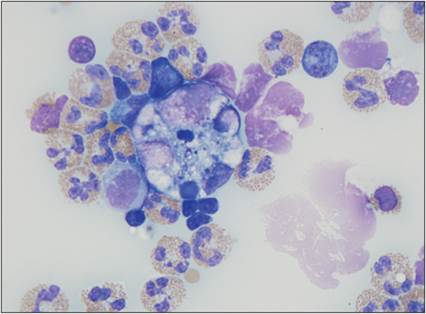
Figure 2.23 Tissue macrophage with phagocytized debris. Cytocentrifuged abdominal fluid from an 8-year-old, spayed female domestic shorthair cat with a history of vomiting and hypoalbuminemia. A thickened gastrointestinal wall and peritoneal effusion were detected ultrasonographically. This image contains several eosinophils and low numbers of macrophages and lymphocytes. The macrophage in the center is large with distended cytoplasm that contains phagocytized cellular debris (Wright–Giemsa, 1,000? magnification).
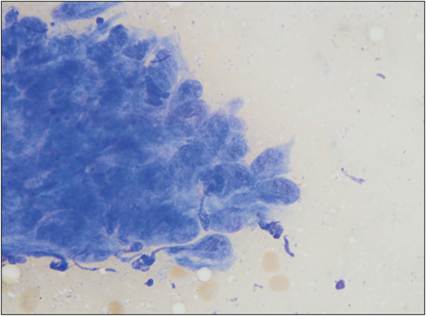
Figure 2.24 Epithelioid macrophages. FNA of a skin mass on a 13-year-old, spayed female Labrador Retriever. A large aggregate of macrophages is shown. Cells have wispy, basophilic cytoplasm and appear cohesive in some areas. Nuclei are rounded with stippled chromatin.
Some of the cell nuclei have dark chromocenters (Wright–Giemsa, 1,000? magnification).
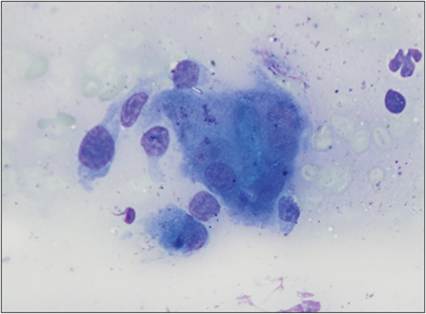
Figure 2.25 Multinucleated giant cell. FNA from an ulcerated dermal mass between the fourth and fifth digits of the left rear limb of a 13-year-old, spayed female Labrador Retriever. There is a large macrophage with several nuclei in the center of the image (the multinucleated giant cell). A few intact macrophages and a neutrophil are also seen (Wright–Giemsa, 1,000? magnification).
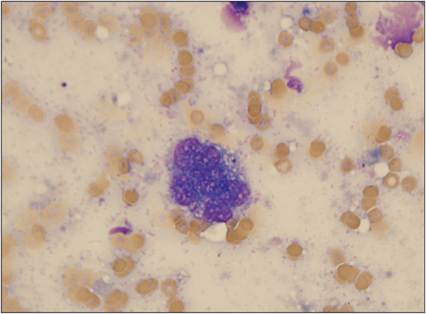
Figure 2.26 Multinucleated giant cell. FNA from a subcutaneous mass on the left hip of a 4-year-old, neutered male domestic shorthair cat. A large vacuolated cell containing several nuclei is shown. Moderate numbers of erythrocytes and a few fragments of lysed nuclei are present in the background (Wright–Giemsa stain, 1,000? magnification).
If there is inflammation present when the monocyte leaves the vasculature, it differentiates into an active inflammatory macrophage (M1), which is microbicidal, proinflammatory, and phagocytic (Ferrante & Leibovich, 2012). IL-12 and IFNγ help to activate histiocytes to become the typical inflammatory macrophage. Recognition of PAMPs and DAMPs is also an important trigger for M1 activation. If other cytokines (IL-4 and IL-13 produced by Th2 lymphocytes) are present, the macrophages tend to differentiate into a classic APC that stimulates the adaptive immune response. The cytokines produced by M1 macrophages drive the inflammatory response. Important examples of cytokines released by M1 macrophages include IL-1, TNFα, IL-6, IL-12, and IL-23.
Like neutrophils, M1 macrophages contain lysosomes that store MPO. They also generate free radicals through a membrane oxidase that converts O2 to O2 − when an opsonized organism is ingested.
Inducible nitric oxide synthase is synthesized, which takes O2 − and arginine and forms NO. The NO then reacts with more O2 − to produce additional free radicals, NO2 − and OH−. During the last phase of the process, lysosomal enzymes degrade what is left of the organism.Macrophages are essential in cleaning up an area for healing and repair. Wound healing macrophages (M2 macrophages) have immunomodulatory functions but are poorly microbicidal (Ferrante & Leibovich, 2012).
Chronic cellular response
Lymphoid cells
Lymphocytes are mononuclear leukocytes that indicate an adaptive immunologic response is ongoing. They are often associated with a response to viral infections. Lymphocytes may be observed several days to weeks after injury occurs. Small lymphocytes are 7–10 μm in diameter and have very sparse basophilic cytoplasm and a small, round nucleus with a dense chromatin pattern (Figure 2.27). Plasma cells are derived from B lymphocytes and are approximately 15 μm in diameter with abundant basophilic cytoplasm and a small, dense, eccentrically located nucleus. These cells often have a pale halo near one side of the nucleus, which is the Golgi apparatus filled with antibody (Figure 2.28).
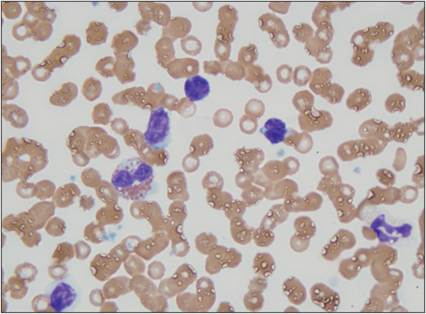
Figure 2.27 Lymphocyte. Peripheral blood from a 10-year-old, intact male mixed-breed dog. Three small lymphocytes (at the top of the image) and a partially lysed lymphocyte (lower left) are shown. Lymphocytes have scant, lightly basophilic cytoplasm and a round to cleaved nucleus. The nuclear chromatin is dense to clumped in mature, intact cells. An eosinophil, a neutrophil, and several erythrocytes are also present (Wright–Giemsa, 1,000? magnification).
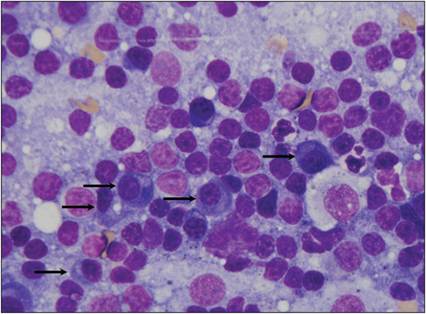
Figure 2.28 Plasma cells.
Reactive lymph node from a dog. The sample contains several lysed cells and small lymphocytes, fewer intermediate-sized lymphocytes, and increased numbers of plasma cells (arrows). A few neutrophils and macrophages are also present (Wright–Giemsa, 1,000? magnification).
Lymphocytes are a major source of polarizing cytokines in inflammatory responses. Several subtypes of CD4+ T lymphocytes are recognized. Each cell subset has a slightly different immune function. Th1 cells secrete IL-2, IL-12, IFNγ, and other proinflammatory cytokines that stimulate cell-mediated responses. Th2 cells tend to produce IL-4, IL-5, IL-10, and other cytokines that promote immunoglobulin production and tend to induce allergic responses. Regulatory T cells predominantly produce IL-10 and are critical in downregulating immune responses. Other subsets of Th cells include Th9, Th17, and Th22 cells (Raphael et al., 2014). Similarly, CD8+ T cell subsets have different functions, driven in part by the cytokines they produce (Mosmann et al., 1997). Tc0 cells secrete IFNγ and IL-4, and Tc1 cells produce IFNγ and TNFα. These cells are cytotoxic through perforin and Fas/FasL pathways. Tc2 cells release IL-4, IL-5, IL-10, IL-13, and low levels of IFNγ. The cytotoxicity of Tc2 cells occurs primarily through perforin.
ILCs are morphologically identical to B and T lymphocytes. Conventional NK cells are a subtype of ILCs that produce large amounts of IFNγ, TNFα, and GM-CSF. These cells inhibit viral replication, induce MHC class I expression, activate macrophages, stimulate granulocytes, and cause apoptosis of certain cells. Several other ILC subsets have recently been identified. The functions of these ILCs are actively being studied (Diefenbach et al., 2014).
Most lymphocytes within inflammatory lesions originate from the regional lymph nodes that drain the site of injury (Halin et al., 2005). Lymph nodes are key sites for the formation of an adaptive immune response to eliminate the agent causing inflammation. Molecules from damaged cells and infectious organisms drain into lymphatics around the site of inflammation. The lymph nodes filter lymphatic fluid and begin processing the antigens present in the fluid to mount an appropriate immune response. In response to antigenic stimulation, T cells proliferate in paracortical regions of lymphoid tissues and begin to coordinate the immune response. This process stimulates B cells to proliferate in follicles. Many of these B cells will develop into plasma cells and migrate to the medulla of the node to form cords. The activated T cells and antibodies produced by this reaction limit the spread of pathogens and protect against reinfection.