Cestodes
Virtually all cestode species of importance to wildlife fall within the Subclass Eucestoda. In this group the adults have an extensive body (strobila), are monoecious, and have multiple segments (polyzoic); virtually all Eucestodes are parasitic in the intestine or accessory ducts of vertebrate animals (Bush et al.
2001). Euces- todes lack an intestinal tract, and have only vestigial oral suckers and pharynxes; nutrition is absorbed through a specialized cuticle (Roberts and Janovy 2000). Life cycles of almost all Eucestodes require at least one intermediate host (Roberts and Janovy 2000). Euces- todes are estimated to have arisen some 350 to 420 million years ago (Hoberg et al. 1999).About 5,000 species of tapeworms currently are known, with new species discovered every year (Zimmer 2000); overall, about 26,000 species of cestodes are estimated to exist (Poulin and Morand 2004). Although considerable progress has been made (Khalil et al. 1994), systematics of cestodes still is in considerable disarray (Hoberg et al. 1997). Currently, about 12 orders of the Eucestoda are recognized (Khalil et al. 1994, Hoberg et al. 1997). Virtually all species of concern to wildlife fall into the Order Cyclophyllidea, which parasitize mammals, birds, reptiles, and amphibians (Leiby and Dyer 1971, Jones and Pybus 2001, McLaughlin 2008); however, occasional species of the Order Pseudophyllidea also are important (McDonald 1969). Coverage in this section is focused on Cyclophyllidean cestodes.
Cyclophyllidean Cestodes: Mammals
Species of Cyclophyllidea have scoleces with four sucker-like hold-fast organelles called acetabula, a neck region of rapid growth, and the main body, called the strobila, comprising many identical segments called proglottids (Fig. 4.10). Immature segments are in the anterior portion of the strobila, mature segments lie in the middle, and old (gravid) proglottids lie on the posterior portion (Bush et al.
2001). Among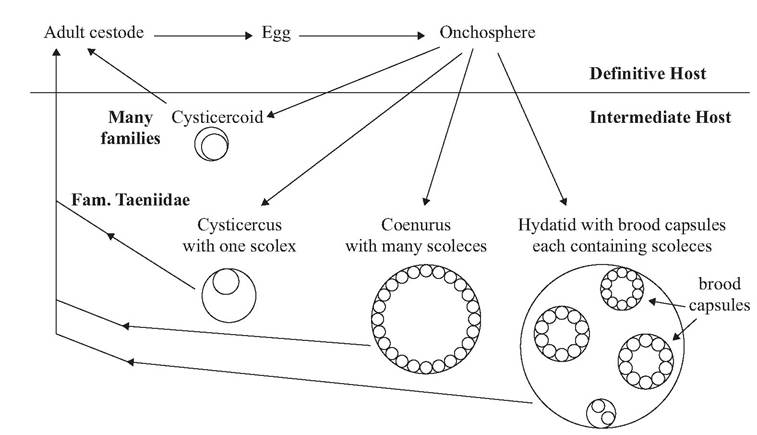
FIGURE 4.11 Life cycle variation among Cyclophyllidean cestodes. Members of the Family Taeniidae may have a cysticercus, coenurus, or hydatid cyst in the intermediate host. Most other Cyclophyllidea, including members of the Family Hymenolepidae, have a cysticercoid.
adult cyclophyllidean cestodes, the most obvious morphological feature is a single compact postovarian vitelline gland. A rostellum, often with an armature of hooks, commonly is present on the scolex. The number of testes varies from one to several hundred. While most species are small, some cestodes can reach 10 m or more.
Adults live in the intestinal tract of the vertebrate definitive host. It is uncommon for adults to cause pathology or other problems in the definitive host. However, in some cases there can be intestinal blockage from numerous strobila, generalized toxic or allergic reactions, mechanical irritation of the intestinal mucosa, reduced availability of select nutrients, or bacterial invasion at sites of attachment of scoleces (Leiby and Dyer 1971).
The life cycles of most Cyclophyllidean cestodes among mammals are indirect (Fig. 4.11). Cestode eggs (Fig. 4.12) are shed with the feces. In some species, eggs are shed from the strobila while the proglottids remain attached (anapoly- sis); in others, the proglottids may detach and leave the host intact (apolysis) before releasing the eggs with the feces (Bush et al. 2001). Within the eggs are hexacanth (six-hooked) embryos, also called oncospheres (Bush et al. 2001).
In rare cases, as among the Family Hyme- nolepidae, no intermediate hosts are required. After eggs are shed with the feces, the oncospheres develop to the cysticercoid state in the egg itself; the oncosphere has a fully withdrawn scolex resembling that of the adult. When ingested by the definitive host, the cysticer- coid emerges and settles in the intestinal tract.
Hymenolepis nana is a well-known example (Soulsby 1968).For the great majority of cyclophyllidean cestodes using intermediate hosts, there are several types of larval stage (metacestode) in the intermediate host (Bush et al. 2001). In some species, the eggs are shed by apolytic proglottids and, when ingested, the oncosphere emerges, penetrates the invertebrate gut wall into the hemocoel, and transforms into a cys- ticercoids, which has a fully withdrawn scolex resembling that of the adult; the life cycle is completed when the cysticercoid is eaten by the definitive vertebrate host. Examples of this pattern include Dipylidium spp. and Raillietina spp. (Soulsby 1968).
The other types of metacestodes among cyclophyllideans are fluid-filled bladders associated with a mammalian intermediate host
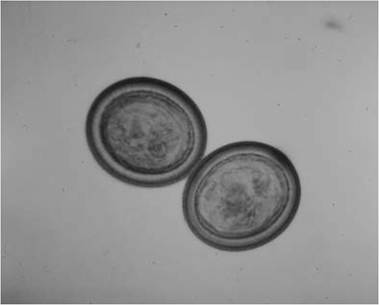
FIGURE 4.12 Ova of Echinococcus granulosus (Courtesy of W. Frank, Universitat Hohenheim, Germany).
(Bush et al. 2001). Once the egg is shed by the carnivore definitive host and ingested by the herbivorous intermediate host, the oncosphere penetrates the gut wall, enters the circulatory system, and is transported to a specific site in the intermediate host. Several types of larval stages are possible, depending on the cestode species, including a cysticercus, coenurus, and hydatid cyst (Fig. 4.11).
A cysticercus (Fig. 4.13) is a fluid-filled bladder with an invaginated scolex including four acetabula and a rostellum with one or two rows of hooks. Once ingested by the definitive carnivorous host, the scolex evaginates in the small intestine, attaches to the gut wall using the acetabulum and rostellar hooks, and begins to grow (Bush et al. 2001). Many members of the genus Taenia follow this body plan (Leiby and Dyer 1971, Jones and Pybus 2001).
Although uncommon, larval stages of a few cyclophyllidean cestodes have a bladder in which the scolex is not inverted; this structure is called a strobilocercus.
Taenia (Hydatigera) taeniaeformis is a parasite using wild canids and felids as definitive hosts; rodents serve as intermediate hosts (Leiby and Dyer 1971, Jones and Pybus 2001). Other cestodes using a strobilocercus include Taenia krepkogorski and T. macrocγstis (Jones and Pybus 2001).In contrast, a coenurus (Fig. 4.14) is a fluid- filled bladder with several to many inverted scoleces developing on the internal membrane, leading to multiplication in the intermediate host; thus, these species are able to multiply both in the definitive host with eggs, and in the intermediate host in the coenurus with a proliferation of (proto) scoleces. Cestodes of wildlife using a coenurus include Taenia (Multiceps) multiceps, T. (Multiceps) serialis, T. endothoraci- cus, T. twitchelli, and T. parva (Jones and Pybus 2001). Taenia multiceps typically uses large canids, foxes, and felids as definitive hosts and uses ungulates as intermediate hosts; in contrast, T. serialis typically uses foxes as definitive hosts and rodents and lagomorphs as intermediate hosts (Leiby and Dyer 1971, Jones and Pybus 2001).
Hydatid cysts (Fig 4.15) are complex, fluid- filled structures that also undergo larval multiplication in the intermediate host. From an internal germinal layer of the cyst, the parasite produces buds (brood capsules) that become free of the cyst wall and float in the hydatid fluid. Within the brood capsules, there is further multiplication as small invaginated protoscolices develop on the brood capsule walls. The life cycle is completed when these larval protoscolices are ingested by the definitive host. The morphology of hydatid cysts varies among the Echinococcus spp. Echinococcus granulosus produces single, fluid-filled vesicles called unilocular hydatids; its life cycle involves large canids as definitive hosts and a variety of wild ungulates as intermediate hosts. In contrast, E. multilocularis produces multilocular (alveolar) hydatid cysts that are able to bud exogenously in the intermediate host and “metastasize” to other tissues in the host (Fig.
4.16); this life cycle involves foxes and other canids as definitive hosts and a wide variety of rodents as intermediate hosts (Leiby and Dyer 1971, Jones and Pybus 2001). Larval stages (metacestodes) sometimes can cause significant pathology among intermediate hosts (Jones and Pybus 2001).Cestode life cycles circulating among wildlife are called sylvatic (literally “forest”) cycles. Those cycling among domestic animals are called pastoral (“pasture”) cycles. Some cestodes, such as
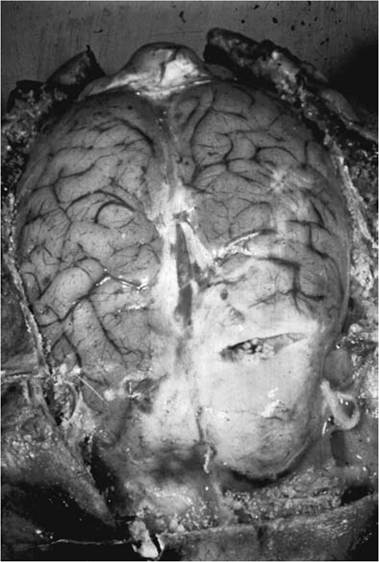
FIGURE 4.14 Taenia multiceps coenurus in sheep brain (Courtesy of W. Frank, Universitat Hohenheim, Germany).
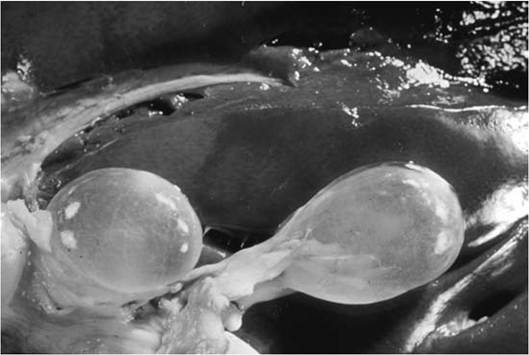
FIGURE 4.13 Cysticercus of Taenia hydatigenea in sheep liver (Courtesy of W. Frank, Universitat Hohenheim, Germany).
Taenia serialis and Echinococcus granulosus, may have independent sylvatic and pastoral cycles.
Control of cestodes in wildlife generally is not feasible and rarely is considered justified. However, treatment of domestic animal definitive and intermediate hosts, including safe disposal of carcasses or offal, would help reduce the prevalences of some cestodes transmissible to wildlife (Jones and Pybus 2001). Tin compounds historically were used against adult tapeworms in domestic birds (Soulsby 1968). More recently a number of anthelminthic drugs have been used for definitive and intermediate hosts (Fraser and Mays 1986). There also is some effort to develop vaccines against cestodes important for domestic animals (Lightowlers 1996, Lightowlers et al. 2000).
There are some good reviews of cyclophyl- lidean cestodes in wild mammals (Leiby and Dyer 1971, Jones and Pybus 2001) and birds (McDonald 1969, McLaughlin 2008). In this chapter, we give a brief overview of several cestode species of importance for definitive or intermediate hosts, or otherwise causing concerns for managers.
Among waterfowl, members of the Family Hymenolepidae are the most common species reported (McDonald 1969). Members of the family Taeniidae generally are considered the most important mammalian cestodes, based on their effects on animal health and economic impacts on livestock (Leiby and Dyer 1971, Jones and Pybus 2001). Within the Family Taeniidae, the genus Taenia is estimated to have 39 (Loos- Frank 2000) or 40 species (Hoberg 2002), with four additional subspecies (Loos-Frank 2000);

FIGURE 4.15 Multilocular hydatid from a pig (Courtesy of W. Frank, Universitat Hohenheim, Germany).
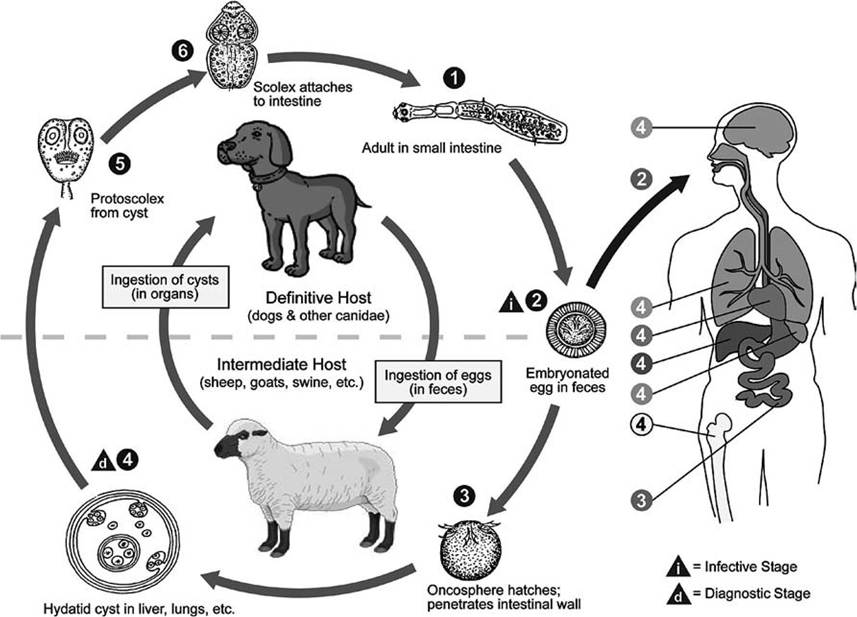
FIGURE 4.16 Pastoral life cycle of the cestode Echinococcus granulosus (Courtesy of Centers for Disease Control and Prevention's Division of Parasitic Diseases and Malaria, www.dpd.cdc.gov/dpdx).
about 25 species involve only wildlife (sylvatic cycles) (Jones and Pybus 2001).
Life Cycles Using a Cysticercus
Taeniasis is an infection of definitive hosts with adult tapeworms of Taenia spp. Eggs or gravid proglottids are passed with feces; the eggs can survive for days to months in the environment. Intermediate hosts become infected by ingesting vegetation contaminated with eggs or gravid proglottids. In the intermediate host's intestine, the oncospheres hatch, invade the intestinal wall, and migrate to the body cavity, striated muscles, or certain body organs, where they develop into cysticerci. Cysticerci frequently can survive for several years in the animal. Definitive hosts become infected by preying upon infected intermediate hosts and ingesting the cysticerci. In the definitive host intestine, the cysticercus develops into an adult tapeworm, which attaches to the small intestine by its scolex and resides in the small intestine, often for years. Length of adult worms can be up to 25 m. The adults produce proglottids which mature, become gravid, detach from the tapeworm, and migrate to the anus or are passed in the stool.
Many species of Taenia producing cysticerci have sylvatic cycles. Few of them cause detectable pathology for the definitive or intermediate host. Taenia hydatigena has a cosmopolitan distribution in many wild and domestic canids and occasionally felids as definitive hosts; many wild animals, principally cervids such as Odocoileus hemionus, O. virginianus, Cervus elaphus, and Alces alces, can serve as intermediate hosts (Jones and Pybus 2001). Cysti- cerci often occur in the body cavity or liver of intermediate hosts.
Taenia ovis krabbei has a predominantly syl- vatic cycle and a circumpolar distribution in wild canids and cervids. Definitive hosts include coyotes (Canis latrans), wolves (C. lupus), dogs (C. familiaris), Arctic foxes (Alopex lagopus), pumas (Puma concolor), and bears (Ursus americanus and U. arctos). Cysticerci are embedded in the musculature of a wide variety of ungulates serving as intermediate hosts. In contrast, T. ovis ovis has a cosmopolitan distribution in a predominantly pastoral life cycle involving domestic dogs and sheep (Jones and Pybus 2001). Some interpret T. ovis krabbei to be a separate species, T. krabbei (Priemer et al. 2002).
Taenia pisiformis is a cosmopolitan parasite using a wide variety of canids (e.g., Canis spp., Lycaon pictus, Vulpes vulpes, Urocyon cinereoar- genteus, and Alopex lagopus) as well as felids (e.g., Lynx spp., Puma spp., Leopardus spp. Panthera spp.) for definitive hosts. Intermediate hosts include lagomorphs (e.g., Lepus spp., Sylvilagus spp.) and rodents (e.g., Sciurus niger, Apodemus flavicollis, Clethrionomys glareolus, Microtus arvalis, Cavia porcella, Ondatra zibethi- cus) (Jones and Pybus 2001). This parasite commonly is found encapsulated in the serosa of the body cavity and viscera of intermediate hosts (Pfaffenberger and Valencia 1988). In addition to its sylvatic cycle, T. pisiformis also has a pastoral cycle in canids and peridomestic rodents and rabbits (Jones and Pybus 2001).
Taenia crassiceps occurs commonly throughout North America, Europe, and Russia. Adults are found in a variety of canids, including wolves (Canis lupus), coyotes (C. latrans), foxes (Alopex lagopus, Vulpes spp.), and domestic dogs. Cysticerci occur in the subcutaneous and muscle tissue of many small mammals, including lagomorphs, moles (Family Talpidae), and rodents such as voles, marmots and woodchucks (Marmota spp.), squirrels (Family Sci- uridae), susliks (Spermophilus spp.), muskrats (Ondatra zibethicus), and lemmings (Family Cricetidae), as well as rats, mice, and hamsters (Jones and Pybus 2001).
Taenia crocutae is an African parasite for which only hyenas (Crocuta crocuta, Hyaena brunnea) serve as definitive hosts, even though lions, hunting dogs, and many other predators ingest cysticerci from the same carcasses. Cysticerci encyst in the muscle of many African bovids, including Adenota (Kobus) kob, Aepyceros melampus, Alcelaphus spp., Connochaetes taurinus, Damaliscus spp., Hippotragus spp., Oryx spp., Tragelaphus spp., Sylvicapra grimmia, and Syncerus caffer (Jones and Pybus 2001)
Although most Taenia spp. using a cysticercus do not cause pathology in the definitive host, there are occasional exceptions among intermediate hosts (Leiby and Dyer 1971). Some cysticerci cause neurological problems (Reitschel 1981, Kirichek et al. 1986), and others can cause liver and visceral damage such as hemorrhage and granulomas (Freeman 1956, 1962; Roth et al. 1991; Jones and Pybus 2001). Such changes make the intermediate host more vulnerable to predation and enhance completion of the life cycle.
Transmission of Taenia spp. among domestic and wild carnivores and herbivores can occur wherever ranges overlap, and this can be exacerbated by increased human pressures on traditional lands used by wildlife; the appearance of T. hydatigenea in East Africa is a recent example of this (Jones and Pybus 2001). Although the larvae of most of these species are not pathogenic, their presence often is of some concern to the public, especially because of the potential that some (e.g., T. solium) have a potential for human infections.
Life Cycles Using a Coenurus
Taenia species using a coenurus more commonly may cause pathology in intermediate hosts. With heavier loads, some coenuri may cause respiratory distress and even mortality (Rausch 1959). At least five species of Taenia form a coenurus in intermediate hosts: Taenia (Multiceps) multi- ceps, Taenia (Multiceps) serialis, T. endothoracicus, T. twitchelli, and T. parva. Of these, T. multiceps can be of significance among wildlife.
More on the topic Cestodes:
- TREMATODES AND CESTODES
- CONTENTS
- INTRODUCTION TO PHYLUM Platyhelminthes
- CONTENTS
- Boon Andrew. The Ethics and Conduct of Lawyers in England and Wales. Hart Publishing,1999. — 808 p., 1999
- Griffiths-Baker Janine. Serving Two Masters: Conflicts of Interest in the Modern Law Firm. Hart Publishing,2002. — 227 p., 2002
- Grisso T.. Evaluating Competencies: Forensic Assessments and Instruments. 2nd edition. — Springer,2002. — 564 p., 2002
- Luban David. Legal Ethics and Human Dignity. Cambridge University Press,2007. — 350 p., 2007
- Ayupova Z.K.. Theory of state and law: textbook. - Almaty: Kazakh University,2015. - 192 pages., 2015
- Allen Danielle, Benkler Yochai et al. (eds.). A Political Economy of Justice. The University of Chicago Press,2022. — 416 p., 2022
- Barnes Rudolph C.. Military Legitimacy: Might and Right in the New Millennium.Frank Cass,1996. — 198 p., 1996
- Bedner Adriaan (ed.).. Real Legal Certainty and its Relevance: Essays in Honor of Jan Michiel Otto. Leiden University Press,2018. — 261 p., 2018
- Fridson M., Alvarez F.. Financial Statement Analysis. John Wiley & Sons, Inc.,2002. — 413 p, 2002
- Banking, Finance, and Accounting: Concepts, Methodologies, Tools, and Applications. IGI Global,2014. — 1593 p., 2014
- Hare C., Neo D. (eds.). Trade Finance: Technology, Innovation and Documentary Credit. Oxford University Press,2021. — 417 p., 2021
- Fligstein Neil. The Banks Did It: An Anatomy of the Financial Crisis. Harvard University Press,2021. — 334 p., 2021
- Cline W.. The Right Balance for Banks. Peterson Institute for International Economics,2017. — 281 p., 2017
- Alsharari Nizar Mohammad (ed.). Banking and Accounting Issues. ITexLi,2022. — 175 p., 2022
- AAP. Guidelines for Air and Ground Transport of Neonatal and Pediatric Patients. 4th edition. — American Academy of Pediatrics,2015. — 488 p., 2015
- Ancha S., Auberle C., Cash D., Harsh M., Hickman J., Kounga C.. The Washington Manual of Medical Therapeutics, 37th edition, LWW, 2022. —1250p., 1250