ribeiroia Ondatrae
causative agent (classification, morphology) Three species currently are recognized in the genus Ribeiroia: R. ondatrae, R. marini, and R. congolense (Johnson et al. 2004), with a possible fourth species in Africa (Wilson et al.
2005). Trematodes of this genus recently have been reclassified from Family Cathaemasiidae into the Family Psilostomidae (Jones et al. 2005). The adult trematodes of all three species are nearly identical. They range in length from 0.8 mm to 5.2 mm, averaging about 1.7 mm at 7 days post-infection to 2.8 mm 21 days post-infection (Johnson et al. 2004).host range and distribution Trematodes in the genus Ribeiroia occur in lentic aquatic environments in North America, South America, the Caribbean, and central and southern Africa (Johnson et al. 2004). Ribeiroia ondatrae occurs in 12 countries of North, Central, and South America (Yamaguti 1971, Johnson et al. 2002, Wilson et al. 2005). There also is one record in a little bittern (Ixobrychus minutus) from Spain (Nogueserola et al. 2002).
In North America, R. ondatrae has a wide distribution, with records from at least 26 U.S. states and four Canadian provinces. Within this region, frequently recorded or heavily infected host species include white pelicans (Pelecanus erythrorhynchos), wood ducks (Aix sponsa), ospreys (Pandion haliaetus), and great blue herons (Ardea herodias) (Johnson et al. 2004). In Central and South America, great egrets (Ardea alba) and cormorants (Phalacrocorax olivaceus) commonly are infected (Johnson et al. 2004). In a ranking of the relative importance of hosts, based on abundance of parasites (mean intensity ? prevalence), in a study in Florida (Forrester and Spalding 2003), white pelicans appeared to be the most important hosts of R. ondatrae, followed by osprey (Pandion haliae- tus) (Johnson et al. 2004).
life cycles and variations The life cycles for Ribeiroia spp.
include rams horn snails (Family Planorbidae) as the first intermediate host, amphibians or fish as the second intermediate host, and birds or occasionally mammals as the definitive host (Fig. 4.9) (Beaver 1939, Basch and Sturrock 1969). Unem- bryonated trematode eggs are passed in the feces of the definitive host and embryonate within 2 weeks. After the miracidia hatch and penetrate a planorbid snail, the sporocysts mature in veins near the kidney of the snail and form rediae; mother rediae produce both daughter rediae and cercariae concurrently. Cercariae are released and penetrate the amphibian or fish second intermediate host, where they encyst to produce metacercariae. Within amphibians, Ribeiroia ondatrae cercariae have a specific searching behavior and encyst as metacercariae primarily around the base of the hind limbs, clo - aca, and sometimes within the posterior coelom. Once ingested by a definitive host, the metacer- cariae lodge in mucosa along the proventriculus in birds or in the posterior stomach of mammals (Beaver 1939, Johnson et al. 2004). In the laboratory, conditions leading to metacercarial excystment of R. ondatrae in birds are consistent with those of the acid-pepsin environment of the proventriculus (Fried and Johnson 2005).reservoirs and transmission Natural infections of R. ondatrae have been recorded from at least 38 species of birds and three species of mammals (Johnson et al. 2004). Experimentally, R. ondatrae has been recovered from several additional species of birds and mammals (Johnson et al. 2004).
Snails of the Family Planorbidae serve as in first intermediate host. These include members of the genera Biomphalaria, Helisoma, and Planorbella (Johnson et al. 2004). Prevalences of Ribeiroia spp. among planorbid snails generally average about 5%, with values up to 15% (Johnson et al. 2004). Prevalence of R. ondatrae among molluscan intermediate hosts increases from spring to fall in all size classes, but may have a midsummer temporary decline in prevalence (Peterson 2007).
The cercariae infect many species of amphibians and freshwater fishes. Among amphibians, Ribeiroia ondatrae has been recorded among 20 species naturally and two additional species experimentally (Johnson et al. 2004). Among fishes, R. ondatrae has been recorded from 13 species naturally and six additional species experimentally, mostly involving cichlids from Mexico or guppies in the Caribbean islands (Johnson et al. 2004).
Many freshwater invertebrate species are cerciophagic and consume large numbers of cer- cariae. Such predation of R. ondatrae cercariae may play an important role in reducing their numbers and thus inhibiting completion of their trematode life cycle (Schotthoefer et al. 2007).
In some regions, high densities of planor- bid snail intermediate hosts are associated
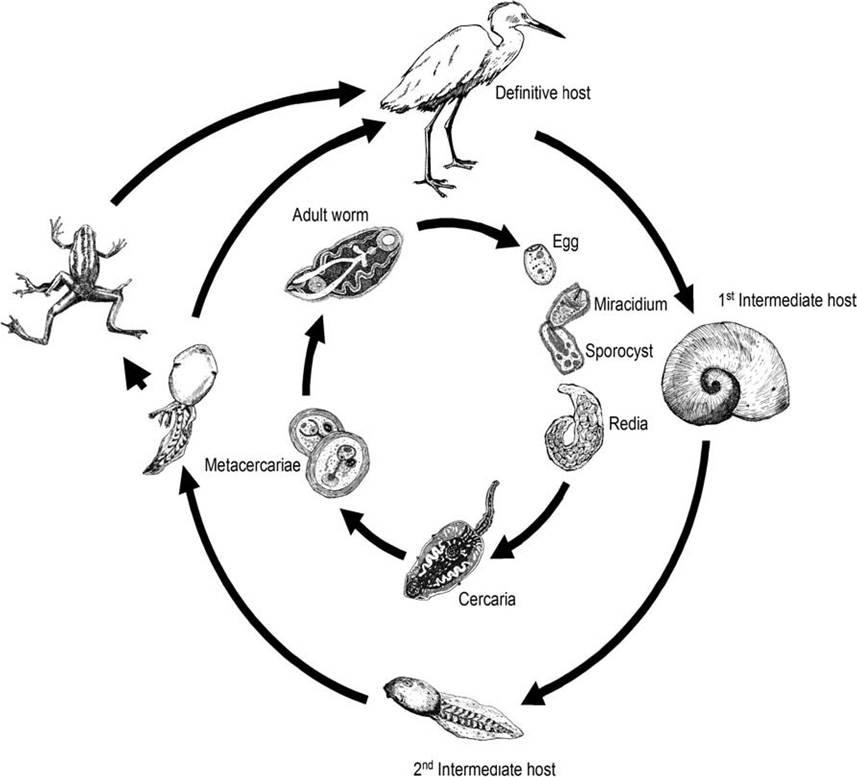
FIGURE 4.9 Life cycle of Ribeiroia ondatrae (from “The complex life-cycle of the trematode, Riberoria ondatrae,” by Brandon Ballangee, 2002/09. Courtesy of the artist and Ronald Feldman Fine Arts, New York, NY).
with artificial ponds containing cattle or fertilizer runoff (Johnson et al. 2004). Eutrophication processes that promote algal production enhance the density of the snail intermediate hosts and are linked to a greater intensity of infection among amphibians and greater per-snail production of infectious parasites (Johnson and Chase 2004, Johnson et al. 2007).
Ribeiroia-induced malformations among infected amphibians may promote completion of the parasite life cycle by increasing the susceptibility of infected amphibians to predation by their definitive hosts (Johnson and Sutherland 2003). Many avian definitive hosts of R. ondatrae prey on amphibians; abnormalities among potential prey items can enhance their capture by bird predators (Fernandez-Llario and HidalgoDeTrucios 1995).
CLINICAL EFFECTS AND IDENTIFICATION In avian and mammalian definitive hosts, infection by R.
ondatrae is generally benign, although it occasionally has caused fatal cases of avian proventriculitis (Newsom and Stout 1933, Cubas et al. 1990). Deep lesions in the mucosa of the proventriculus of experimentally infected birds (Beaver 1939) also have been observed; R. ondatrae was reported as causing similar lesions in naturally infected green herons (Butorides virescens) (Riggin 1956), although the parasite may have been mistaken for R. marini (Basch and Sturrock 1969). Stomach ulcers from R. ondatrae have been reported among experimentally infected mice (Malek 1977).There are no specific reports on the effects of R. ondatrae on its snail intermediate hosts. A related trematode, R. marini, can cause castration and premature death among infected snails (Huizinga 1973, Nassi 1978). There is also little information regarding the pathogenesis of R. ondatrae to infected fish.
However, R. ondatrae has been implicated as a widespread cause of limb malformations in several species of frogs, toads, and salamanders (Johnson et al. 1999, Johnson et al. 2002, 2004). Malformations include extra limbs, missing limbs or parts of limbs, and skin webbings (Johnson et al. 2004). Many of the malformations vary among host species. For example, extra limbs and digits are the most common malformation in experimentally infected Pacific tree frogs (Pseudacris regilla) and northern leopard frogs (Rana pipiens) (Johnson et al. 1999, Schotthoefer et al. 2003); skin webbings were most common in exposed western toads (Bufo boreas) (Johnson et al. 2001a). Also, some infected toads experienced forelimb malformations, including extra and missing limbs, not observed in other species (Johnson et al. 2001a).
The frequency and severity of malformations in amphibians is closely tied to the intensity and timing of infection (Johnson et al. 1999, 2001a, 2002). Infection intensity is influenced by host size and activity, habitat use, and immune response (Johnson et al. 2001a, Kiesecker 2002, Stopper et al.
2002). Ribeiroia ondatrae effects also vary with stage of tadpole development at the time of infection. There is high mortality among tadpoles infected at the pre-limb bud stage, and less mortality but high malformation if infected at the limb bud stage; in contrast, there may be no detectable effect with infections acquired at the later paddle stage (Schotthoefer et al. 2003).The mechanism by which R. onda- trae induces malformations is not known. Mechanical damage to limb buds may be an important cause (Stopper et al. 2002), but the extensive diversity of malformations associated with Ribeiroia spp. infections is evidence that other factors are acting; for example, some signaling molecules may be produced at improper doses or times (Gurrieri et al. 2002, Johnson et al. 2004).
Adults of the three known Ribeiroia spp. are very similar, and often require use of PCR and DNA sequencing to distinguish them (Wilson et al. 2005). A PCR technique also has been developed to identify Ribeiroia spp. in snail intermediate hosts; with this test, Ribeiroia spp. appear to have little cross reaction with the cercariae or adults of other trematodes tested (Reinitz et al. 2007).
population effects Host diversity inhibits transmission of R. ondatrae and reduces amphibian disease as a result of consistent linkages among species richness, host composition, and community competence. Highly competent hosts dominate in species-poor assemblages, whereas more resistant species become progressively more common in diverse assemblages (Johnson et al. 2013)
As a direct or indirect consequence of infection, many amphibians die prior to sexual maturity, causing some to consider Ribeiroia ondatrae as a significant contributor to the widespread declines and extirpations of amphibian populations (Johnson et al. 2001b). See the Case Study on Amphibian Declines in the Special Topics (Chapter 11)” for assessing the importance of this parasite in the context of other causes of amphibian declines.
special problems In 1995, a group of Minnesota school children encountered numerous northern leopard frogs with extra digits and limbs, or missing digits and limbs (Kaiser 1997). While malformed amphibians have been noted for many years (Kaiser 2003), widespread reports have been evident only since the mid- 1990s (Johnson and Sutherland 2003).
The role of R. ondatrae as one cause ofamphib - ian deformities is well established (Johnson et al. 2004). There is evidence for mass malformations among amphibians associated with R. ondatrae back to the 1940s (Johnson et al. 2003). While this problem among amphibians is not new, the prevalence of trematode-induced malformations has increased (Johnson et al. 2003). Eutrophication has been implicated as an important contributor to the increasing impact of R. ondatrae on amphibian populations (Johnson et al. 2007).
In some cases, there may be synergistic effects between R. ondatrae infection and such factors as pesticides, nutrient runoff, introduced species, competition, or predation (Kiesecker 2002, Johnson and Sutherland 2003). For example, salamanders with limbs affected by both injury and trematode infection had 3 to 5 times more abnormalities than salamanders affected by either factor alone (Johnson et al. 2006).
Little is known regarding the specific conditions leading to epizootics of trematode- associated malformations in amphibian populations (Johnson and Sutherland 2003). Distribution and abundance of infected avian hosts probably is important (Johnson et al. 2004), as is the density of infected planorbid snails, which determine the parasite’s distribution (Johnson et al. 2002).
CONTROL AND HOST IMMUNITY No control has been implemented against this parasite. If instituted, control efforts should focus on interrupting or reducing transmission of R. ondatrae among its various hosts, with snails being the most feasible target (Johnson et al. 2004). Suggested measures include reducing nutrient flows into aquatic systems to reduce the planorbid snail populations (Blaustein and Johnson 2003)
Additional snail control could involve molluscicides (Johnson et al. 2004). Another possibility is use of snail predators such as mollus- civorous fish, aquatic insects, or crayfish (Stauffer et al. 1997, Mkoji et al. 1999, Chase 2003). However, many snail predators also adversely affect amphibians and may exacerbate some problems of declining amphibian populations.
There is evidence for host immunity against R. marini in goldfish (Basch and Sturrock 1969, Huizinga and Nadakavukaren 1997). However, there is little information on immunity to R. ondatrae in either fish or amphibians (Johnson et al. 2004).
Tadpoles have some behavioral defenses against invading trematode cercariae. When tadpoles of Rana spp. and Bufo spp. sense cercariae contacting their skin, they make explosive movements to shed those parasites; however, cercariae that enter the junction of a tadpole’s body and tail are particularly difficult to brush off and once there, Ribeiroia spp. can easily enter the inguinal region from this recess (Taylor et al. 2004). Factors that decrease a tadpole’s activities, such as predator threat or certain pesticides, may increase susceptibility to cercarial penetration. In the field, Bufo spp. tadpoles have lower prevalences of Ribeiroia spp. infections than Rana spp. tadpoles; Bufo spp. tadpoles are less palatable than those of Rana spp. and can risk more conspicuous evasive maneuvers (Taylor et al. 2004).
Blood Flukes (Schistosomes)
Schistosomes (blood flukes, Family Schistoso- matoidea) live in the hepatic portal and pelvic veins of birds and mammals (Bush et al. 2001, Huffman and Fried 2008). In contrast to most trematodes, schistosomes generally are dioecious, with male schistosomes having a gynecophoral canal in which the female lies in permanent copula (Platt and Brooks 1997). Although a number of schistosome species occur in wildlife, they rarely cause significant problems. Several genera of avian schistosomes (e.g., Heterobilharzia, Ornithobilharzia, Trichobilharzia) have cercariae that can penetrate the skin of humans and cause an allergic reaction known as schistosome cercarial dermatitis, or “swimmer’s itch” (Roberts and Janovy 2000). However, schistosomes are best known for their significant impacts among humans, which they use as definitive hosts. Schistosome life cycles involve only a snail intermediate host, with no second intermediate host (Roberts and Janovy 2000).
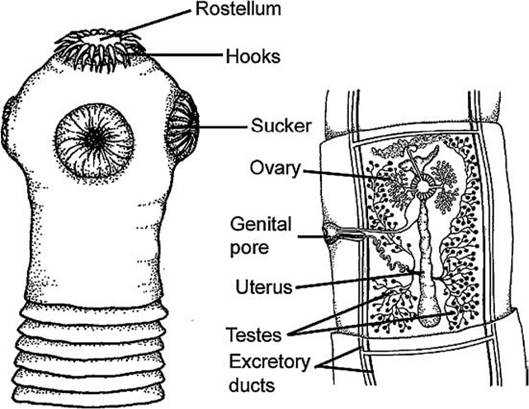
FIGURE 4.10 Typical morphology of a Cyclophyllidean cestode, not to scale (Courtesy of Antoine Morin, Biodidac, biodidac.bio. uottawa.ca).
Although eggs pass out with the feces or urine, they first must pass through a vessel wall and then through the wall of either the alimentary or urinary tract to reach the lumen and ultimately the external environment. Lesions associated with schistosome infections in definitive hosts result from eggs lodged in tissues rather than direct impacts from adult or immature schistosomes (Pence and Rhodes 1982, Bush et al. 2001). A few schistosomes also are of importance to wild birds (Huffman and Fried 2008).
More on the topic ribeiroia Ondatrae:
- Early observations suggest that parasites cause amphibian deformities
- A field experiment suggests that multiple factors influence frog deformities
- A laboratory experiment tests the role of parasites
- The studies of amphibian deformities that we discussed earlier in this chapter illustrate several ways in which ecologists seek to answer questions about the natural world.
- List of Illustrations
- Answers to Figure Legend Questions
- Connections in nature can lead to unanticipated impacts
- Answers to Review Questions
- Answers to Hone Your Problem-Solving Skills Questions
- Boon Andrew. The Ethics and Conduct of Lawyers in England and Wales. Hart Publishing,1999. — 808 p., 1999