Connective tissue
The term connective tissue is used in several contexts. First, it is a general name for a diverse collection of tissues with varying functions. This includes the connective tissues proper, which will be outlined further, as well several specialized tissues.
Although connective tissue histology is extremely varied, there are nonetheless generalities that can be made to compare these tissues with others, for example, epithelial tissues. One of the most apparent is that the relative density of cells is much less than in epithelial tissue. As the name implies, connective tissues serve to support, unify, and "connect" other tissues to allow the creation of more complex tissues and organs. The "connecting" properties of connective tissues depend on products (largely proteins and complex carbohydrates) synthesized and secreted by connective tissue cells. Connective tissue cells (derived from the mesoderm germinal layer) become surrounded by their products so that they are "suspended" in a sea of their own making. So in summary, connective tissue consists of cells and extracellular fibers (protein polymers) embedded in a matrix consisting of ground substance and fluid. These fibers are divided into three types: collagen, reticular, or elastic. If you imagine a gelatin dessert with bits of string, scattered hair, and perhaps some fine paper fibers, youwill have a reasonable mental image of the extracellular matrix of many connective tissues. Now in your imaginary connective tissue, add a random suspension of small raisins to represent connective tissue cells (primarily fibroblasts) and you have an idea of the composition.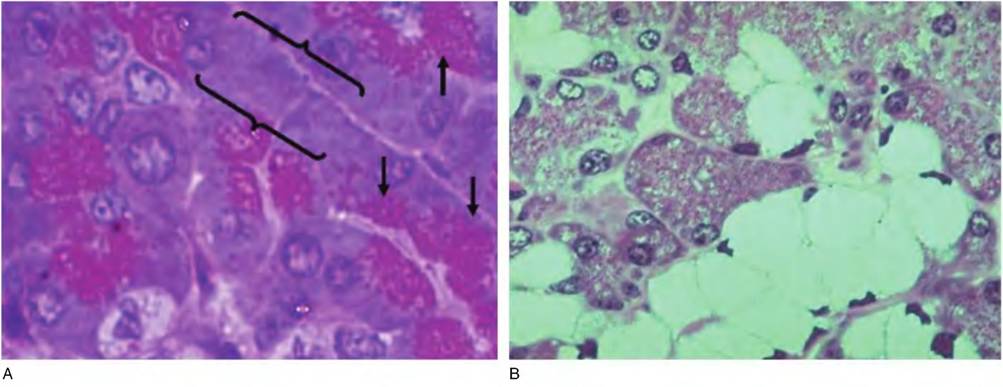
Fig. 4.36. Serous and mucous glands.Panel Ashows an area of pancreatic tissue. These serous-type cells exhibit abundant red-staining secretory granules in the apical regions of the cells (arrows) as well as densely basal cytoplasm (brackets).
Panel Bshows a portion of salivary gland; the pale cells are mucus-secreting cells and the surrounding darker-stained cells serous secretory units.One final idea related to the cells that are present is that some are considered fixed; they are always constituents of the connective tissue in question, that is, fibroblasts or osteoblasts in bone, for example. Other cells called wandering cells, various blood-derived cells, macrophages, neutrophils, plasma cells, and so forth, may be present as well.
When tissues are fixed, embedded, and prepared for histological examination, the material between cells can sometimes appear as empty space, but this is far from the truth. When more specialized stains are used, many of these specialized products can be readily visualized. Our purpose is to simply introduce you to the idea of connective tissues and to provide some examples of the many varieties of connective tissues (Box 4.2).
Classification of connective tissue
It is not easy to formulate a classification of connective tissue that is completely adequate for all situations. However, some organization helps the learning process by producing at least a general frame of reference. General connective tissues, called proper connective tissues, are divided into loose, dense regular, and dense irregular types. The primary distinction here is with the arrangement and relative number of fibers between the connective tissue cells. The loose connective tissue contains aggregates of loosely arranged fibers and a relatively large number of cells. Loose connective tissues have many fibroblasts but many other cells as well. A major site of loose connective tissue is just below the various epithelial layers that cover internal and external body surfaces. These are sites where antigens and other foreign materials can be present; it should not be surprising that many of
Box 4.2 Tissues, cells, and technology
Just as microchips and computer technology have revolutionized communications, advances in digital imaging, microscopy, and software for analysis of images is also dramatically changing information that can be obtained from the study of tissues and cells.
Many of you are likely aware of Western blotting techniques that allow the detection and quantitation of proteins that are first separated by electrophoresis then transferred to membranes. Proteins are then identified based on their molecular weights as well as visual detection of enzyme- linked or fluorescence-tagged antibodies. Not only is it possible to identify the protein, but the amount of the protein present in the sample is proportional to the signal produced.In a recent review, Ellis et al. (2012) describe such techniques to evaluate changes in mammary development (specifically ontogeny of myoepithelial cells) and, perhaps more importantly, indicate the growing importance of imaging in the animal and biological sciences and the need for training of new scientists to take advantage of these emerging technologies.
the other cell types in loose connective tissue are the "wandering" immune-related cells that migrate from local capillaries.
Dense connective tissue has more abundant, thicker fibers but not as many cells. The most abundant cell type is usually the fibroblast, the cell responsible for the synthesis and secretion of the proteins of the fibers. Dense irregular connective tissue is found in the protective capsules around organs and surrounding developing glands. Dense regular connective tissue is more specialized, in that the regular arrangement of fibers is important in providing strength as in tendons and ligaments. Other specialized connective tissues that will be briefly described and illustrated include cartilage, bone, adipose, and blood.
The first illustration in this section (Fig. 4.37) shows a simple mesenteric spread. This image of fixed stained tissue that anchors and protects the intestines demonstrates many of the general features of connective tissues, namely, the presence of extracellular fibers, space between cells, and varying types of cells.
As might be expected, there is some gray area in deciding the difference between loose and dense connective tissues.
How compacted must the material be to be considered dense? The tissue just underneath many epithelial surfaces, for example, the skin or areas of the reproductive tract, is loose connective tissue, but there is certainly variation. The immature 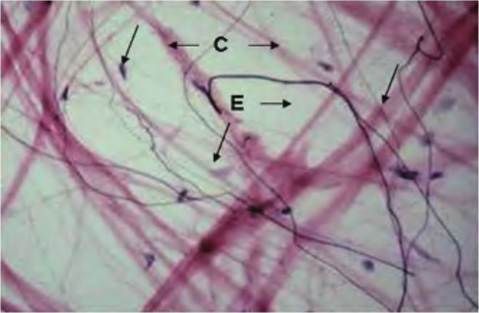
Fig. 4.37. Mesentery spread. The mesentery, which serves to anchor and support the loops of the digestive tract, is very thin. Many of the very thin fibers (E) are elastin fibers, while the thicker pink fibers are collagen (C). Some of the dark specks are nuclei of fibroblasts (arrows).
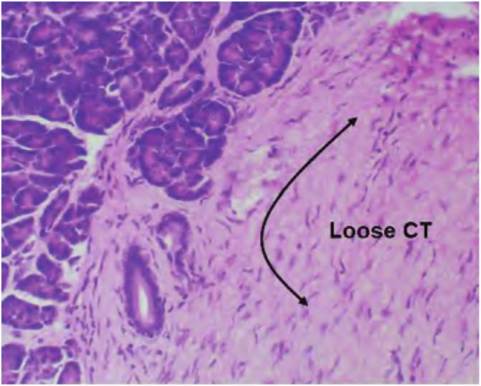
Fig. 4.38. Loose connective tissue (CT). This image from the pancreas illustrates the general features of the loose CT present under the epithelial regions of many tissues. At the low magnification of this image of H&E-stained tissue shows only the amorphous pink of faintly stained collagen fibrils and scattered fibroblast nuclei in the region adjacent to the glandular tissue.
mammary gland provides good examples of this variation. The connective tissue immediately adjacent to the ducts (intralobular) is less dense than the connective tissue some distance away from the developing ducts (interlobular). Other organs also have abundant areas of connective tissue. Figure 4.38 shows a section from the pancreas. The epithelial tissue to the upper left is part of the exocrine glandular tissue of the organ. The pale pink tissue to the right is a region of loose connective tissue that supports and anchors the glandular structure. Notice that relatively little detail
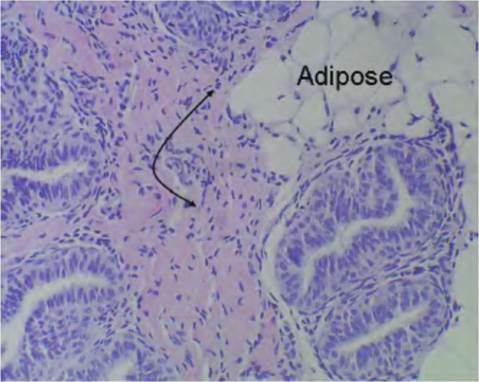
Fig. 4.39. Loose connective tissue bovine mammary. This image illustrates an area of loose connective tissue from the bovine mammary gland. Notice the similarity with the tissue from the pancreas.
However, this section also shows an area with a cluster of adipocytes or adipose tissue, a specialized type of connective tissue, in addition to the band of pinkish connective tissue in the center of the image.or apparent cells are apparent in this region. Collagen fibrils and other extracellular matrix proteins and ground substance occupy most of the space. The scattered fibroblasts and other cells only become apparent at higher magnification. Figure 4.39 illustrates an area of similar loose connective tissue from the bovine mammary gland.
At a higher magnification (Fig. 4.40), some of the fibroblast nuclei become apparent and the swirls of light pink-stained collagen fibrils and other more amorphous proteins can be seen. Many of the fibroblasts are so closely aligned with the collage fibrils that it is difficult to distinguish between cell cytoplasm and the extracellular matrix. This is especially true for paraffin-embedded tissues. However, the darkly stained nuclei of various cells stand out in the preparation. The next example (Fig. 4.41) shows a sample of dense and loose connective tissue in the same specimen. Another difference is the staining of the specimen. It has been stained with a dye called Sirius Red and counterstained with another dye called Fast Green. In this preparation, the collagen appears as bright red, wavy fibers and the epithelium is stained a faint bluish-green, but there is little detail for the epithelium because there is no specific nuclear stain.
The higher-magnification image shown in Figure 4.42 also illustrates the difference between dense irregular and loose connective tissue. It is easy to see that it is largely a matter of degree. The red collagen fibers appear in both regions, but just underneath the epithelial surface of a large epithelial duct (to the upper
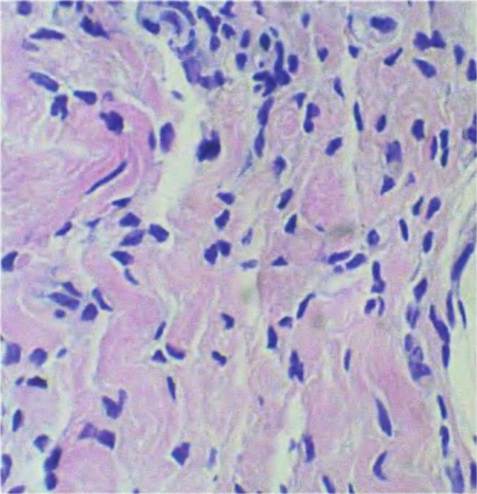
Fig. 4.40. Loose connective tissue detail. An example of loose connective tissue for the mammary gland of a cow is shown at higher magnification.
Most of the darkly stained nuclei are from fibroblasts. The pink, somewhat wavy pattern is largely from collagen fibrils.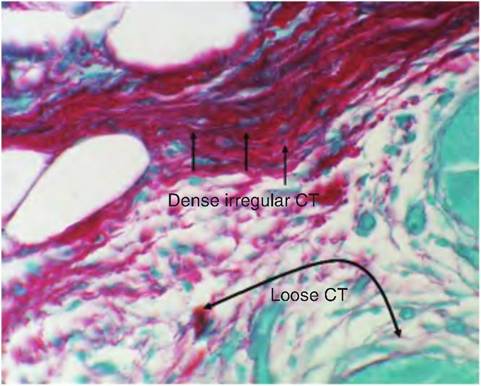
Fig. 4.42. Dense irregular connective tissue. Dense irregular versus loose connective tissue is contrasted with loose connective tissue in this image from the developing mammary gland of a heifer.
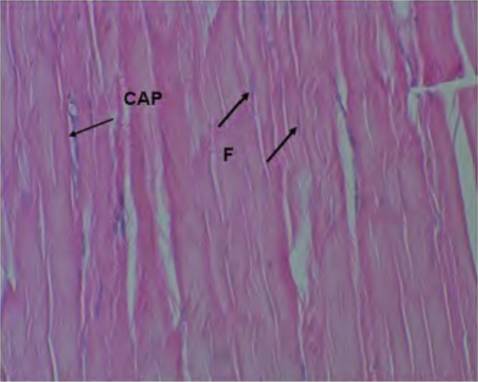
Fig. 4.43. Histology of a tendon. This section through a portion of a tendon demonstrates the largely directional, parallel orientation of the fibers from top to bottom. This corresponds with the direction of greatest strength. Fibroblast nuclei (F) are very difficult to detect, and their pale blue-stained nuclei seem to blend with the general structure of the fibers. An approximate longitudinal cut through a capillary (CAP) is also indicated.
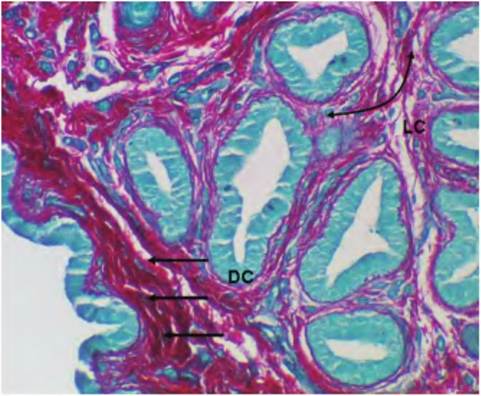
Fig. 4.41. Si rius Red-stained connective tissue. Examples of loose (LC),compared with dense (DC), irregular are indicated. Cross sections of epithelial ducts appear a faint pale bluish-green color.
left), the fibers are much more apparent. The very pale blue structures embedded in this dense collagen matrix are mostly the nuclei of fibroblasts.
As you might suspect, dense connective tissue is especially strong because of the abundant fibers. A subclass of dense connective tissue, dense regular connective tissue, is found in tendons and ligaments. This tissue gets its name because of collagen and other fibers that are uniformly arranged. This is easy to imagine in a tendon or ligament where there is a need for great tensile strength, usually along a particular axis. In fact, injuries and tears to ligaments are especially common when forces are applied perpendicular to the direction in which the fibers are oriented. Figure 4.43 illustrates a histological section through a portion of a tendon. The staining is not specialized for detection of collagen or other fibers (H&E staining), so it is difficult to distinguish individual fibers or fibroblasts, but the general regular orientation of the fibers is evident from top to bottom in this image (Fig. 4.44).
The image in Figure 4.45 shows the result of immunostaining for the presence of one of the subclasses of collagen in the developing tissue. In this procedure, an antibody specific for type IV collagen was incu-
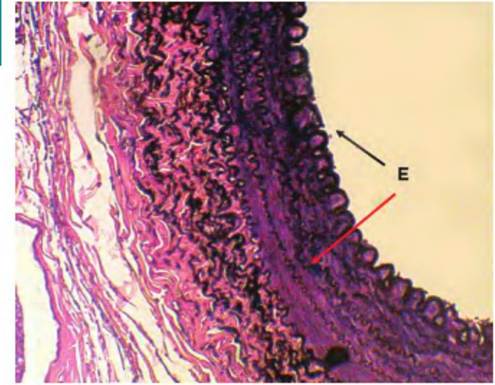
Fig. 4.44. Elastin fibers. This is a section through the wall of a large artery. In this specimen, the sample has been stained with a special dye to emphasize elastic fibers. They appear as black wavy lines (E) oriented around the circumference of the artery. Given the need for many arteries to expand and recoil in response to changes in blood pressure, it is easy to rationalize the function of these fibers.
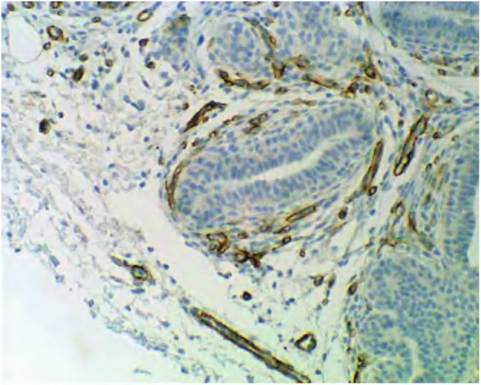
Fig. 4.45. Collagen immunocytochemistry. This specimen is an immunocytochemical preparation showing the expression of type IV collagen in the developing mammary gland. The brown staining (where antibodies against collagen type IV are located) indicates the location of this protein. Notice that it is almost exclusively located around blood vessel endothelial cells as part of the basal lamina of these vessels.
bated with the tissue section, then the location of the bound antibody detected by the use of a secondary antibody linked with an enzyme. When the enzyme is activated in the presence of an appropriate substrate, a reaction product is deposited at the site(s) where the antibody is bound. In this specimen, you can see that type IV collagen is almost exclusively located around the endothelial cells of blood vessels. The use of immunocytochemistry provides a powerful tool to study details of tissue development. If you compare the very evident red stain for "total" collagen fibers in Figure 4.41 and Figure 4.42 with the restricted expression of collagen type IV in Figure 4.46, you can see that the distribution of type IV collagen is very concentrated or localized. In fact, type IV collagen is most often expressed in the basal lamina. This explains its abundance around the endothelial cells of the capillaries, but its apparent absence around the developing epithelial ducts in the tissue is surprising. It may be that basal lamina associated with epithelium does not fully mature until later in development. This is further suggested by the image shown in Figure 4.46, which shows staining for expression of type IV collagen in the mammary tissue of a lactating cow. In this case, the expression occurs around the blood vessels as well as the secretory alveoli. This supports the idea that the basal lamina is not fully developed until major ducts and/or the alveoli are fully formed. Again, the point here is showing how these histology techniques can provide information about tissue growth, development, and ultimately physiological function. Other collagens include types I, II, and III. Type I is associated with connective tissues of skin, bone, tendon, and ligaments. Type II is predominantin cartilage and type III infetal tissues and connective tissue capsules around various organs.
Specialized connective tissues
Adipose tissue is a specialized connective tissue consisting of fat-storing cells called adipocytes. There are two types of adipose tissue, white (or unilocular) and brown (or multilocular). In well-nourished animals (or people), adipose tissue forms a nearly continuous layer within the connective tissue underneath the skin. It is called panniculus adiposus and is part of the hypodermis. It is generally believed that adipocytes are derived from the mesenchymal cells of the mesoderm germinal layer. Unilocular adipocytes are often large cells 100 μm or more in diameter. The size is a reflection of the accumulation of a large lipid droplet that occupies most of the cell area. This causes the nucleus and most of the cytoplasm to be pushed to one side of the cell, so that the cells are often described as having a signet ring appearance when sectioned.
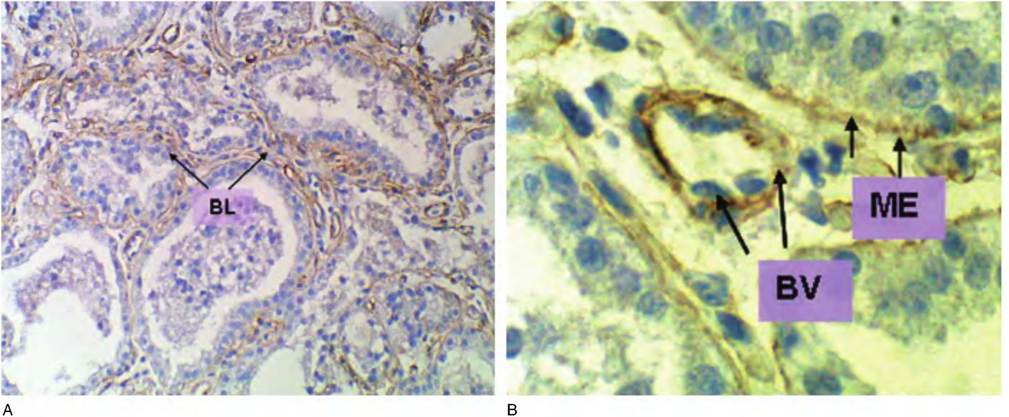
Fig. 4.46. Type IV collagen. Immunostaining for type IV collagen in mammary tissue from a lactating cow is shown. In this functionally and developmentally mature tissue, there is marked expression of type IV collagen in the area of the basal lamina (BL) immediately adjacent to the epithelial cells of the alveoli as well as blood vessels (A). The figure insert (B) at higher magnification (oil immersion) makes the expression around both regions evident. In the center of the figure, there is a cross section through a capillary with each three nuclei of endothelial cells evident (BV and arrows). Other arrows (ME and arrows) indicate staining associated with the basal lamina of the mammary epithelial cells.
In most histological preparations, the lipid is lost because of the lipid solvents (ethanol and xylene) used to prepare the tissue for embedding in paraffin or plastic. White adipose tissue is most abundant. Brown or multilocular adipose tissue is less common but is present in large amounts in hibernating animals. In other animals, it is relatively more common in newborn and young animals. Unlike much of the lipid that is mobilized by lipolysis to produce ATP from white adipose tissue, brown adipose tissue adipocytes have mitochondria that have evolved to use most of the mobilized lipid to produce heat energy rather than ATPThis probably explains the importance of these specialized cells in hibernating animals and in newborns where maintenance of body heat is so critical.
The following series of slides shows examples of white adipose tissue. Figure 4.47 and Figure 4.48 illustrate the general structure adipose tissue at a relatively low magnification. Both of these images are from samples embedded in paraffin. As indicated in an earlier section, samples prepared in this manner are usually relatively thick (4-7 μm), so some details of cellular structure are difficult to see. Figure 4.49 and Figure 4.50 illustrate sections of white adipose from samples embedded in plastic. The stains used are different (H&E vs.Azure II), but more importantly, plastic embedded sections can routinely be cut at 1 μm or less. The difference in cellular detail between paraffin and plastic embedding is striking but especially so with actively secreting epithelial cells, that is, pancreas, liver, mammary gland, and so forth.
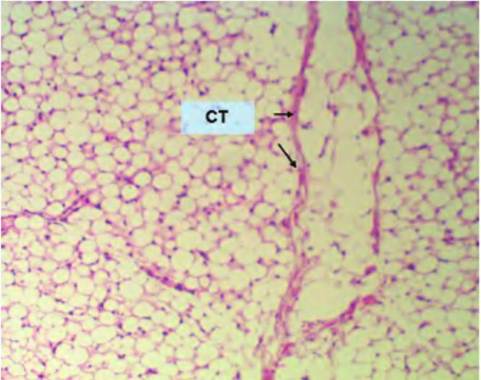
Fig. 4.47. Low-power adipose. This is a low-magnification view (10? objective with 10? eyepiece, i.e., 100-fold) of white adipose tissue. The essential feature is the appearance of lots of seemingly closely packed, circular structures, which are the profiles of sectioned adipocytes. Remember, in the living tissue the spaces would have been occupied by lipid droplets. The other prominent features are several bands of connective tissue, which appear here as pink bands (CT). At higher magnification (Fig. 4.48), the cellular organization becomes more apparent.
Figure 4.51 shows the structure of small parts of the four different adipocytes. This image also provides some perspective on the size of these cells. The connective tissue between the cells has cross sections of two vessels with several RBCs. A reasonable estimate
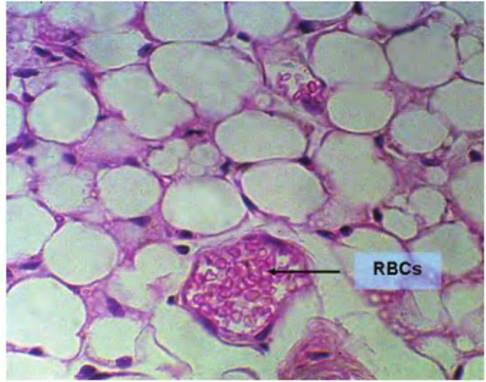
Fig. 4.48. Adipose tissue, paraffin, high magnification. Here, cellular organization becomes more apparent. There is only a small rim of cytoplasm and nuclei positioned at the periphery of the cells.
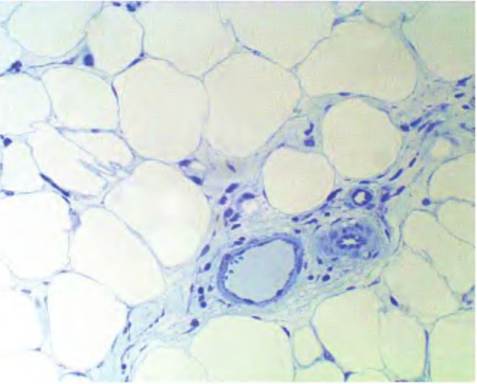
Fig. 4.50. Adipose tissue. Adipose tissue from a region of the mammary gland is shown. The tissue was embedded in plastic, sectioned at about 2μm in thickness, and stained with Azure II. Cell profiles are distinct, but there is little cytoplasm.
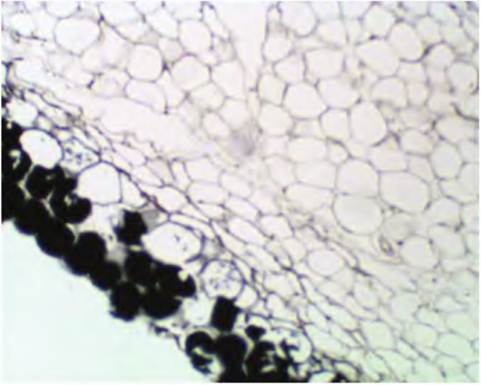
Fig. 4.49. Adipose tissue, low-power, osmium-stained. In this case, some of the lipid was retained in the cells during processing and the tissue was subsequently stained with osmium tetroxide. This chemical has a very high affinity for lipid and causes the appearance of a dense black product. Notice the darkly stained adipocytes in the lower left of this tissue section.
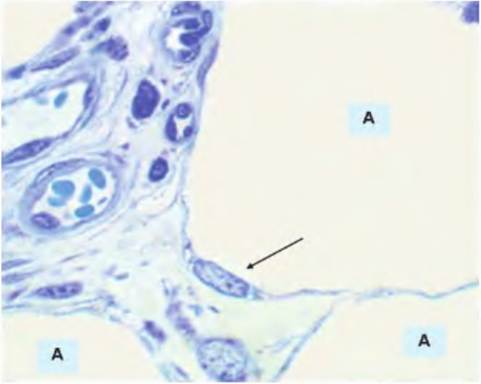
Fig. 4.51. Adipose tissue high magnification. In this section, portions of four adipocytes (A) are visible. The large open spaces contained stored lipid. The nucleus of one adipocyte (arrow) appears at the rim of the cell. The upper left has cross-sectioned profiles of a small artery and companion venule. A number of red blood cells appear in the lumen of the artery.
for an RBC is 7 μm across its widest dimension, so the adipocytes (A) in comparison are clearly many times larger. The nucleus of one of the adipocytes is also apparent compressed to one edge of the cell. Nuclei for the other adipocytes do not appear in this image.
Although the structure of other specialized connective tissues will be discussed as we consider specific systems, it is worth introducing bone and cartilage. In the case of the long bones of the limbs, for example, tibia andhumerus, they begin as cartilage models (essentially miniature versions) in the fetus. As the animal develops, the cartilage is progressively replaced by bone in a process called endochondral ossification. Three cartilage types can be distinguished. These are, in order of abundance, hyaline, fibrocartilage, and elastic cartilage. Hyaline cartilage is grossly characterized by the presence of a shiny, milky, glass-like homogenous matrix. It should be familiar to you as the cartilage that appears at the end of a chicken leg or the gristle you sometimes bite into at the edge of a chicken breast. Cartilage very clearly illustrates the idea that connective tissues have relatively few cells but lots of extracellular matrix components. However, unlike many other connective tissues, which usually have a ground substance with an abundance of collagen fibrils, cartilage is also rich in three types of proteoglycans. These complex polymeric molecules are composed of a core protein with various attached glycosaminoglycan (GAG) chains. The three-dimensional shape of the molecule resembles a miniature bottlebrush with the stem being the core protein and the bristles the GAG chains. The GAGS include hyaluronic acid, chondroitin sulfate, and keratin sulfate. These sugar chains are highly charged (polar) so that cartilage is very highly hydrated. This aids the cushioning associated with cartilage, but the sparseness of cells in the mature tissue and relative lack of blood supply also explains the difficulty of repair to injured cartilage. Figure 4.52 is an example of hyaline cartilage from the trachea. In this instance, these C-shaped rings of cartilage (with the opening of the C facing a dorsal direction) provide the rigid support to hold the entrance to the respiratory tract open. You may recognize the upper right edge of the tissue as an epithelial layer. Although you could not identify its characteristics at this level of magnification, you might recall that pseudostratified columnar epithelial cells cover the respiratory tract above the level of the bronchioles. The cartilage appears as two "islands" within the connective tissue. As cartilage is Createdby chondroblasts, the cells progressively mature into chondrocytes as their own products trap the cells. These mature chondrocytes come to reside in spaces called lacunae.
The insert in Figure 4.52 shows a higher-power image of hyaline cartilage; it appears as a largely avascular field of matrix with scattered chondrocytes. The chondrocytes produce this matrix so that in its fully mature state, the cells are literally trapped. The spaces immediately surrounding the cells are called lacunae. This terminology also applies to bone, but the cells within the spaces are called osteocytes. The outer covering of the cartilage is called the perichondrium and can be divided into an outer, more protective, capsulelike protective layer, and an inner region with more cells that are chondrogenic; that is, these cells can be induced to produce more active chondrocytes. Although the matrix surrounding the cells appears rather homogenous in typical H&E-stained sections, the matrix contains collagenous fibrils, which add strength. Often the chondrocytes appear very close together; these are called isogenous cell clusters because they arise from division from a single predecessor cell. Growth in this manner, with subsequent addition of more matrix material around the cells, allows for interstitial growth of the cartilage or growth within the substance of the cartilage. In contrast, development of additional cartilage in the outer perichondrium leads to growth at the surface called appositional growth. This later style of development explains the continued lengthening of the long bones prior to the time of epiphyseal plate closure at puberty. At this time, there is cessation of the generation of new cartilage needed for appositional growth and to provide the ground substance and matrix necessary for osteogenesis. Sections of the fetal mouse foot provide an excellent tissue sample to study the structure of cartilage and developing bone.
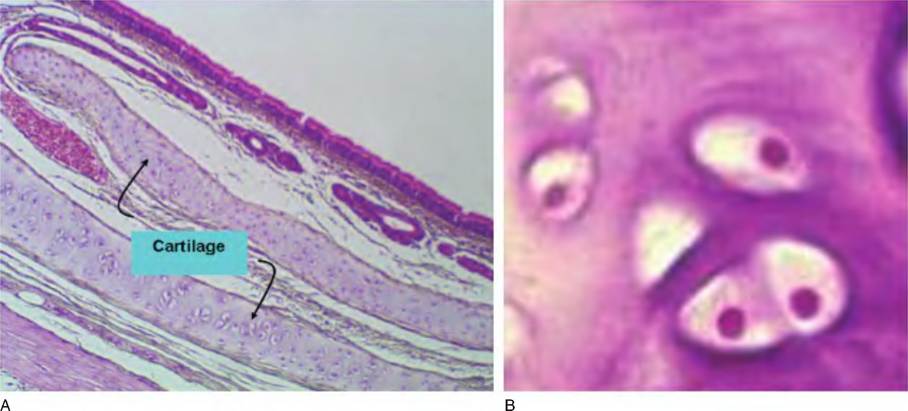
Fig. 4.52. Section of cartilage from the trachea. This low-power image shows the general features of the trachea. (A) The internal surface of the trachea (upper right) is covered by epithelial cells (pseudostratified columnar epithelial cells), and just underneath, there are several mucus-producing glands. Further into the tissue, portions of sectioned hyaline cartilage appear (arrows). The inset (B) shows a grouping of chondrocytes in greater detail. The cell structure is barely visible, but the nuclei appear as distinct dots.
In mature bone, for example, the humerus, two types of osseous tissue can be distinguished. The outer portion of the bone is called compact or dense bone, and it has a very distinctive structure. At the microscopic level, it is often viewed by preparing what are called ground bone samples. Essentially preserved, fixed samples are cut into very thin wafers and mounted on slides. This allows an examination, primarily of the matrix structure. The center or medullary cavity, in addition to marrow components, is composed of an interlacing, spiderweb-like component of bone called spongy or cancellous bone. Both types of bone have osteocytes and various other bone cells (osteoblasts, osteoclasts), but the organization of the matrix is much less regular. Figure 4.53 shows a relatively low-power image of a sample of ground bone to illustrate major features, while Figure 4.54 gives a higher magnification to show some of the details of osteocytes and lacunae.
At higher magnification (Fig. 4.54), structures of an individual Haversian system become apparent. The dark areas are the spaces that in living bone would
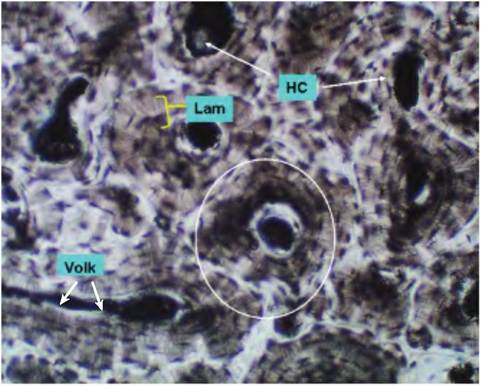
Fig. 4.53. Compact bone low power. The structure of compact bone can be imagined as a series of miniature trees where in the center of the trunk is a canal called the Haversian canal (HC), and the rings or lamellae (Lam) represent the concentric growth rings. These Haversian systems, or osteons (circled area), generally align themselves along the longitudinal axis of the long bones. Other channels called Volkmann canals (VoIk) pass perpendicular to intersect the central Haversiancanal and link the central passageways to provide for entrance of blood vessels and nerves. In compact bone, the spaces between lamellae areoccupied by the osteocytes, which come to be surrounded by the matrix they have produced and secreted, much like Chrondrocytes in cartilage. Small fissures, called canaliculi, also radiate out from the lacunar spaces that hold the mature osteocytes.
contain the osteocytes. Other apparent structures are the canaliculi. Bone located between the circular osteons is called interstitial bone.
Figure 4.55 shows an area of ossification in a developing bone that is destined to be spongy bone. The irregular shape of the ossifying tissue and lack of organization with respect to the lamella is evident. The area of ossification is stained pale blue; areas with lacunae containing trapped osteocytes are scattered in the matrix. Areas to the outside (stained red) are areas of bone marrow.
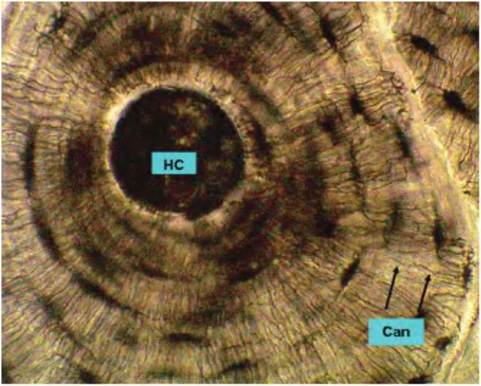
Fig. 4.54. Haversian canal. This high-power image of compact bone shows the ringlike lamellar structure surrounding the Haversiancanal (HC). Each ring is a lamella. The dark areas are the lacunae. These spaces contain the osteocytes. Radiating away from the lacunae are small fissures called canaliculi (Can). These spaces allow for diffusion of nutrients and waste products.
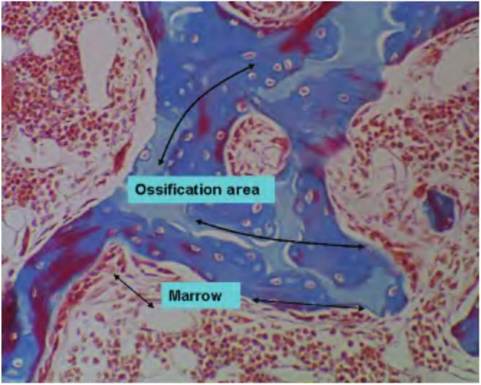
Fig. 4.55. Ossification. This image illustrates the process of ossification in a region of spongy bone. The center (blue-stained) area contains cartilage cells undergoing ossification to become osteocytes. The surrounding area will contain red bone marrow.
A final type of connective tissue to consider is blood. At first glance, it seems odd to think of blood as connective tissue, but once you consider that it consists of a matrix (plasma) with suspended cells, it matches our prior definition of connective tissue. Of course, the majority of cells are erythrocytes or RBCs, typically 4-5 million∕mm3. The white blood cells (WBCs) or leukocytes average 6,000-9,000 ∕mm3 and are divided into two subclasses based on the presence or absence of cellular granules,that is,granulocytes Versusagran- ulocytes. Agranulocytes include large and small lymphocytes (30-35% of the total white cells) and monocytes (3-7%). Granulocytes include neutrophils (55-60%), eosinophils (2-5%), and basophils (0-1%). Platelets, also called thrombocytes, are small structures that average 200,000-400,000∕mm3). Figure 4.56 shows a relatively low-power image of a blood smear to provide some perspective. There are two neutrophils and one lymphocyte in the field. It is clear that RBCs greatly outnumber WBCs. Figure 4.57, Figure 4.58, Figure 4.59, and Figure 4.60 show some of the features of various leukocytes. Specifically, Figure 4.57 shows a lymphocyte; Figure 4.58 shows a monocyte (these are the agranulocytes); Figure 4.59 has a neutrophil on the right and an eosinophil on the left; and Figure 4.60 shows a basophil.