Muscle tissue
Epithelial, muscle, and nervous tissues are sometimes called composite tissues. This simply means that although the tissue primarily is composed of the cells that give the tissue its name, there is also the incorpo-
ration of at least some connective tissue.
For muscle tissue in particular, the connective tissue elements are essential for functioning of the tissue. First, because of the metabolic demands of the muscle tissue, availability of nutrients and capacity to get rid of wastes is critical. The connective tissue sheaths that surround skeletal muscles and the connective tissue layers that penetrate around even individual muscle cells (fibers) provide passageways for capillaries. Second, the various connective tissue sheaths from around the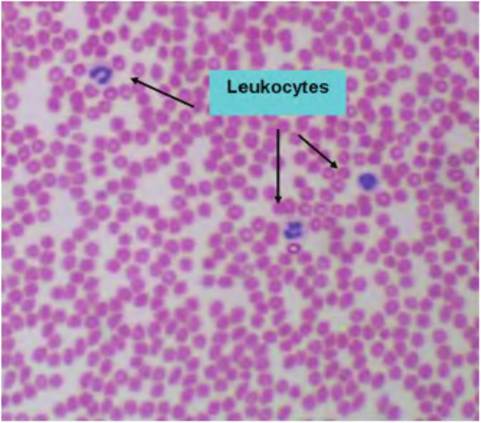
Fig. 4.56. Blood smear. This low-power image shows a field of pale-staining erythrocytes and several leukocytes (three neutrophils).
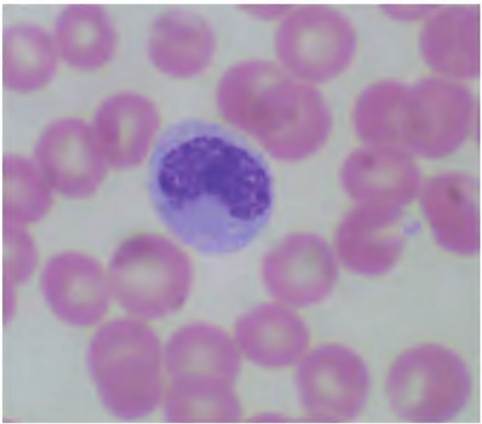
Fig. 4.58. Monocyte. Monocytes are characterized by the presence of a kidney bean-shaped nucleus and under normal conditions are relatively rare, -2-8% of the total.
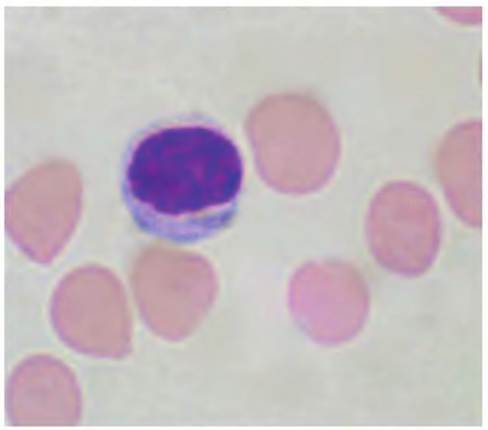
Fig. 4.57. Lymphocyte. Lymphocytes have a large nuclear to cytoplasmic ratio; that is, typically only a thin rim of cytoplasm may be evident.The nucleus is uniformly stained roughly ovoid in shape, and cells are common in the circulation, -25% of the total.
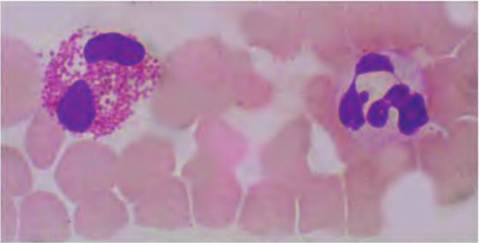
Fig. 4.60. Eosinophil and neutrophil. The eosinophils that appear to the left of the image gets its name from the dark staining of its granules with the red acidic dye eosin.
The nucleus is Iobed but relative rare, ~1-2% in the circulation. The neutrophil on the right is commonly ~50% of the total and, unlike other granulocytes, the granules stain poorly with either basic or acidic dyes, for example, the neutral of the name. The nucleus has two to five lobes, a bit like beads on a string. Neutrophils are also called polymorphonuclear leucocytes (PMNs) because of this characteristic.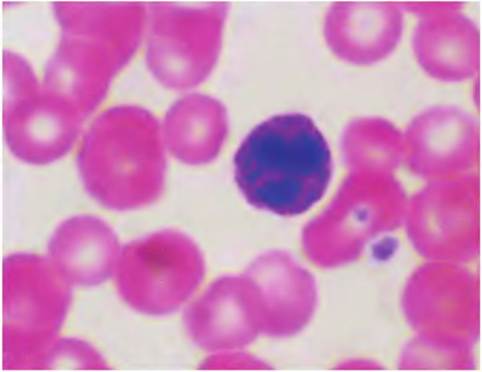
Fig. 4.59. Basophil. Basophils have abundant darkly stained blue or purple granules (with standard basic dyes), and the cells are smaller than neutrophils or eosinophils. The nucleus is typically Iobed and numbers are usually very low, ~1-2% of the total.
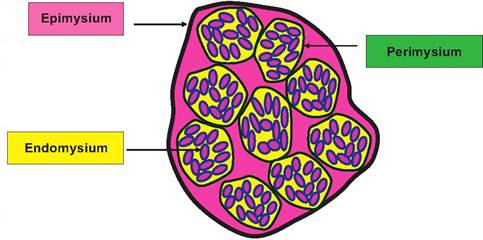
Fig. 4.61. Cross section, skeletal muscle. A thin layer of connective tissue, the endomysium, wraps each skeletal muscle fiber or cell. Bundles of muscle fibers called fascicles (outlined in yellow) are wrapped by the perimysium and the entire muscle by the epimysium that is continuous with the tendon that attaches skeletal muscles to bone.
outside of a muscle to bundles of muscle fibers to individual muscle fibers (epimysium, perimysium, and endomysium, respectively) means contractile elements are anchored together. This allows for unified, smooth functioning of the muscle.
There are three types of muscle cells. First, the familiar skeletal or voluntary muscle is named because its contraction is linked with movement of the skeletal system. A second, cardiac muscle, shares many features with skeletal muscle including the presence of striations. The third type, smooth muscle, is very widely distributed and gets it is name because it does not exhibit striations present in the other types. Like epithelial tissue, muscle tissues are highly cellular but in contrast are also highly vascularized. Muscle cells have elegantly constructed myofilaments, primarily combinations of actin and myosin protein filaments that allow for contract and tissue movement.
Details of muscle cell structure and the sliding filament model for contraction will be described in a subsequent chapter. Our purpose in this section is to introduce the basics of muscle tissue histology.Skeletal muscle
At a gross level, each skeletal muscle is a distinct unit composed of several tissues. Of course the muscle cells or fibers predominate, but adequate functioning requires connective tissue sheaths, blood vessels, and nerve fibers. One of the most unusual features of skeletal muscle cells is that they are multinucleated, large elongated cells. Furthermore, the internal volume of the cell is chiefly dedicated to the packaging of a complex array of myofibrils. These individual myofibrils are composed of overlapping thin filaments, primarily made of the protein actin, and the thick filaments made of the protein myosin. These myofibrils, like tightly packed wires in a telephone cable, are arranged along the longitudinal axis of the individual muscle cells. Let us consider organization of a common muscle (the biceps) cut in cross section across the belly or gaster of the muscle. Our examination would allow us to distinguish the three connective tissue sheaths as diagramed in Figure 4.61. In this diagram, the smallest circular units (dark pink) within the yellow background represent the individual muscle cells that have been cut in cross section. The endomysium that surrounds individual cells is indicated by the black outline of each cell. The cells are grouped together in fascicles, outlined in yellow, and the perimysium that anchors these muscle bundles by the black outline around each. Groups of fascicles come together to create the muscle, which is surrounded by the epimysium. When skeletal muscle cells are sectioned along their longitudinal axis, the highly organized arrangement of the myofilaments is evident by distinct banding or striations. These alternating dark and lightly stained regions reflect areas in which only thin filaments, thick filaments, or both overlap.
This explains why skeletal and cardiac muscle is also referred to as striated muscle. Some of the microscopic features of skeletal muscle are outlined in subsequent figures as well as the molecular organization of the contractile filaments (Fig. 4.62).At higher magnification,with cells cut in cross section, you can distinguish profiles of individual cells (Fig. 4.63) and in some preparations individual myofibrils within the cells. It also becomes apparent that there are multiple nuclei per cell and that they are located at the peripheral edges of the cell. It is only when cells are sectioned longitudinally that striations are seen (Fig. 4.64). However, only at very high magnification is it possible to decipher the detail of the filaments responsible for the banding pattern of the muscle sarcomere (Fig. 4.65). A diagrammatic representation of the sarcomere is provided in Figure 4.66. Mechanics of muscle contraction will be discussed in a subsequent chapter.
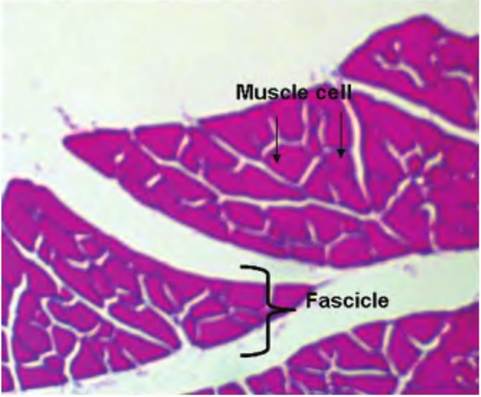
Fig. 4.62. Cross section, skeletal muscle. The image at very low magnification shows portions of several muscle bundles or fascicles; profiles of individual muscle cells or fibers appear as the irregular shapes within the muscle bundles.
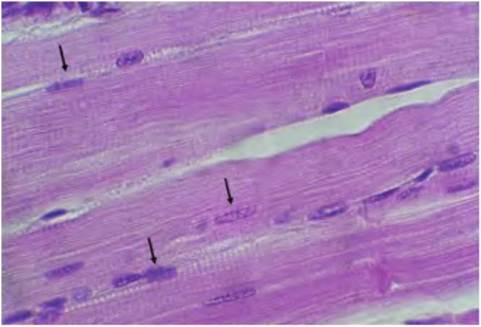
Fig. 4.64. Skeletal muscle, longitudinal. This section from the bovine tongue illustrates the peripheral location of nuclei (arrows) and the pattern of striations characteristic of skeletal muscle.
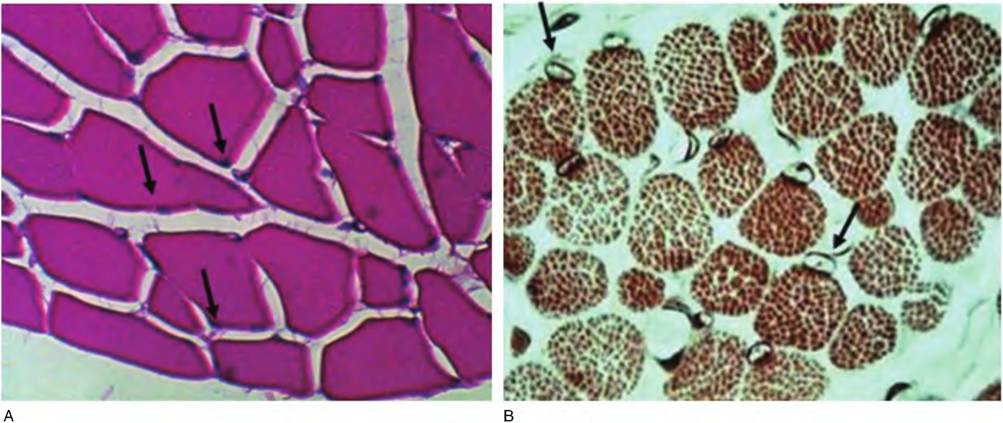
Fig. 4.63. Skeletal muscle, cross section, high. At progressively higher magnification, it is possible to see profiles of individual muscle cells and the fact that many of the cells are multinucleated (arrows, A). Depending on the preparation, the myofibrils may also be discernible (B).

Fig. 4.65. Transmission electron microscope (EM) image, skeletal muscle. This image shows portions of several sarcomeres. A single sarcomere is bounded on either side by the Z-Iine, the site where the thin filaments are anchored. The region bounded by the pale band is called the I band of the sarcomere. Notice that the I band at each end of the sarcomere also contains the I band for the adjacent sarcomere. The darker band is the A band. Depending on the degree of contraction, a lighter band (the H band) can be seen in the center of the A band. In this particular sample, because the muscle was contracted at the time it was prepared, the H band is barely visible as the thin pale stripe in the center of the A band. The darker stripe (M-Iine) is the location where the thick myosin filaments are anchored.
At first glance, the striations of the muscle cell fibrils seem to be repeating disks stacked along the length of the cell. However, when examined in the electron microscope (Fig. 4.65), the striations clearly appear only in the myofibrils andnot within the cytoplasm of the cell. The alternating dark and light bands are due to the relative density in regions where there are only thin or thick filaments. Furthermore, the pattern is very highly ordered. This is because the fibers are linked together into an organizing unit called a sarcomere. The sarcomeres are linked together, something like train cars along the entire length of the myofilaments. The myofilaments are essentially repeating groups of linked sarcomeres. The contraction process is explained by the ability of the sarcomere to shorten and then relax. Regulatory details will be discussed in a subsequent chapter, but the fundamental process involved is the movement of the thinner actin filaments moving along the thicker myosin filaments. As indicated by the diagram in Figure 4.66, as pairs of thin filaments (attached at the Z-line) on either end of the sarcomere move toward one another, the sarcomere shortens.
Since the sarcomeres are linked and the myofibrils are anchored and cells are bound to other cells by the endomysium, this results in shortening of entire bundles of muscle cells and, if stimulated sufficiently, the entire muscle.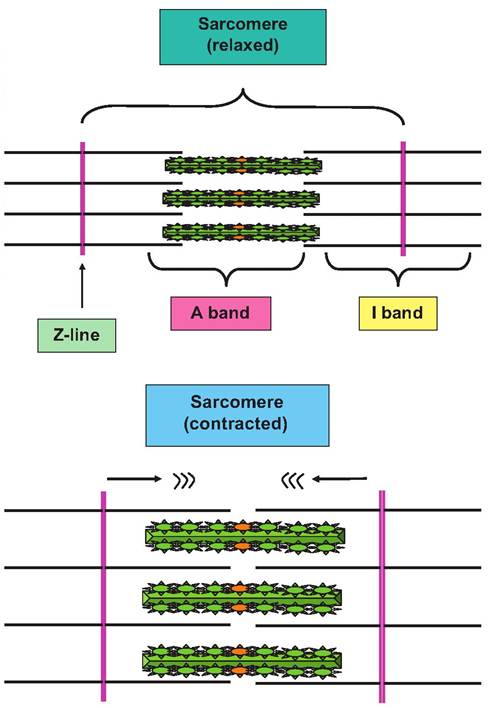
Fig. 4.66. Structure of sarcomere. A schematic view of sarcomere shortening is shown to illustrate changing relationships between I, A, and H bands as contraction occurs. The thick filaments are outlined as green bundles with protruding myosin heads and the thin filaments as simple black lines. Note that the A band stays constant but that the H and I bands get smaller as the Z-Iines and ends of the thin filaments approach.
Cardiac muscle
Skeletal muscle and cardiac muscle are very similar. Both have striations and essentially identical sarcomere structures. However, individual muscle cells (fibers) are typically shorter and contain fewer nuclei per cell, that is, they are often binucleated. There are also some differences in the mechanics of contraction regulation. In skeletal muscle, the calcium needed for interaction between the actin and myosin comes from storage within the muscle cells, the sarcoplasmic reticulum. For cardiac cells, extracellular calcium plays a more important role than in skeletal muscle. Also, although it is possible to learn to change one'sheart rate under special circumstances, cardiac contraction is usually considered involuntary. Control of cardiac
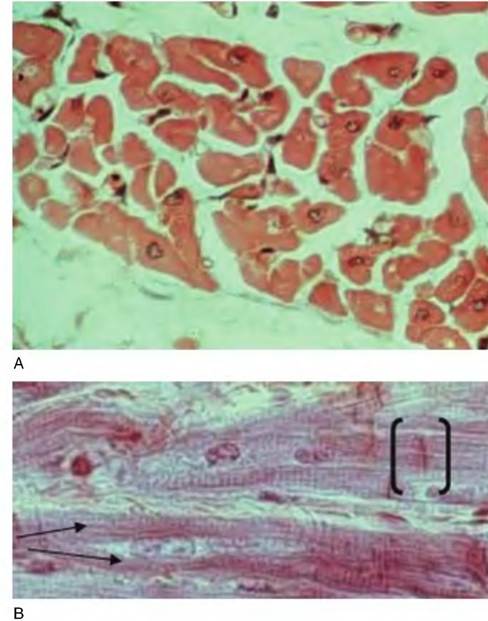
Fig. 4.67. Cardiac muscle. The upper panel (A) shows a group of cardiac muscle cells cut in cross section. In profiles where the nucleus is present, it is located toward the center of the cell, in contrast to skeletal muscle. The lower panel (B) shows a longitudinal section of cardiac muscle. Note the striations and, at the arrows, branching of a cell. The dark bars (brackets) are intercalated disks that join cells in series.
rate and force of contraction is clearly important to supply the oxygen and nutrients necessary for varying levels of activity. It should be no surprise that regulation involves close coordination between the nervous and endocrine systems as discussed in our review of cardiovascular physiology.
At a histological level, cardiac cells have striations but also frequently are branched; nuclei are located centrally within fibers; and groups of cells are linked longitudinally by complexes of gap junctions within regions called intercalated disks. These features allow skeletal muscle fibers to be distinguished from cardiac muscle fibers in histological sections as illustrated in Figure 4.67.
Smooth muscle
Although it is common to describe all muscle cells as fibers, it is easier to think of the large multinucleated skeletal muscle cells or the cardiac cells linked by intercalated disks as fibers in a commonsense view. Smooth muscle cells by comparison are small, spindle- shaped cells that taper on either end. Like the cardiac cells, smooth muscle cells have a single, centrally located nucleus. Despite their small size, groups of smooth muscle cells are physiologically vital. These cells are found in the walls of hollow viscera and in the walls of all but the smallest blood vessels. In the walls of many tubular visceral structures, the smooth muscle cells appear in two distinct layers. For example, in the muscularisexterna of the GI tract, there is an outer layer of longitudinally oriented cells and an inner layer that goes around the circumference of the tract. Smooth muscle cells that are arranged in these coordinating layers of cells are called single unit smooth muscle and are the most common type. In contrast, about 1% of the cells in smooth muscle act rather independently and are called multiunit cells. Both classes are involuntary. However, the single unit smooth muscle is more likely to be impacted by the secretion of various hormones than the multiunit cells that depend primarily on neural input for contraction to occur.
For example, within the muscularisexterna of the GI tract, groups of smooth muscle cells act as pacemaker cells for the tissue. When these cells spontaneously depolarize, this induces rhythmic contractions that pass through the tissue. The level of autonomic nervous system activity or secretion of some hormones (i.e., estrogen or progesterone influence on uterine smooth muscle) can also alter the normal pattern of spontaneous contractions that occur or the strength of contractions.
As the name suggests, there are no apparent striations in smooth muscle cells. There are, however, thin and thick myofilaments anchored to the cytoskeleton that allow shortening of the cells. Since the cells are linked together (single unit cells), this allows the coordinated contraction of the tissue layer. Multiunit cells are typically anchored to extracellular proteins. Extracellular calcium concentrations also have a greater impact on contraction of smooth muscle cells than skeletal muscle cells because of the lack of sarcoplasmic reticulum for intracellular storage of calcium. Differences in the appearance of crosssectioned versus longitudinally sectioned smooth muscle cells are illustrated in the diagrams shown in Figure 4.68 and examples of smooth muscle tissue from the small intestine shown in Figure 4.69 and Figure 4.70.