Dictyocaulus viviparus
CAUSATIVE AGENT (CLASSIFICATION, morphology) The lungworm Dictyocaulus viviparus is one of four species of the genus Dictyocaulus in the Family Dictyocaulidae, Superfamily Trichostrongyloidea (Anderson 2000).
Others include D. arnfieldi, a parasite of equines, D. filarial, a common parasite of sheep and goats, as well as wild antelope and deer, and D. eckerti, also in deer (Schneider et al. 1996, Anderson 2000). All of these worms are medium-sized and occur in the trachea and bronchi of their hosts.host range and distribution Dictyo- caulus viviparus is considered a cosmopolitan parasite among many domestic animals such as cattle, sheep, and goats (Soulsby 1968), as well as members of the deer family (Cervidae) (Soulsby 1968, Anderson 1992), and additional wild bovids including bison (Bison bison) (Locker 1953, Boddicker and Hugghins 1969), muskoxen (Ovibos moschatus) (Samuel and Gray 1974), and nyala (Tragelaphus angasi) (Keep 1971).
life cycles and variations Adult D. viviparus live in the air passages of the lungs, rather than within the parenchyma as do most other lungworms (e.g., Protostrongylus spp.). The D. viviparus eggs hatch in the lungs, and the L1 larvae crawl up the respiratory passages, are coughed up by the host, swallowed, and pass out with the feces (Fig. 3.6) (Anderson 1992); such L1 larvae can be detected with a living larvae technique such as the Baermann Test (Zajac and Conboy 2006). The L1 larvae are susceptible to desiccation and sunlight, and many die within the first 2 weeks on pasture (Anderson and Prestwood 1981). Under suitable conditions of temperature and moisture, the L1 larvae molt to the L2 stage in a few hours, which, in turn, molt to the infective L3 larvae in a few more hours (Anderson 1992). The L3 larvae are fairly resistant to cold and freezing conditions and moderate drying (Anderson 1992).
reservoirs and transmission As a direct life cycle nematode, the reservoirs are the infected cervids and bovids in the regions where D. viviparus occurs. The definitive host becomes infected by ingesting infective L3 larvae contaminating its food. When ingested, the L3 larvae penetrate the mesenteric lymph nodes of the ileum, cecum, and upper colon and remain for 3 to 8 days (Jarrett et al. 1957,
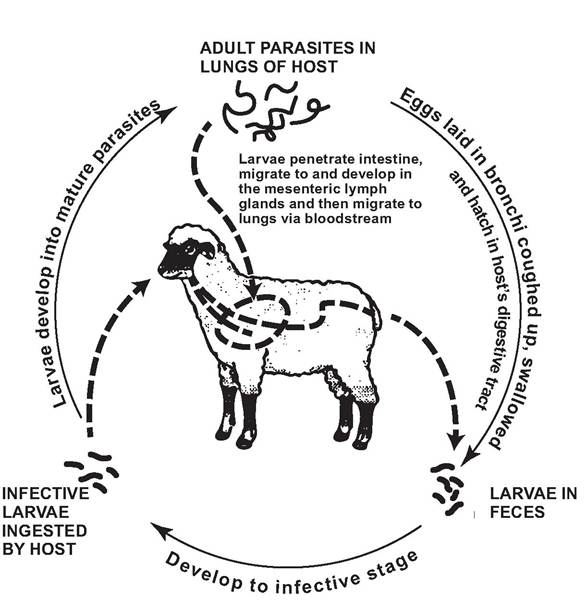
FIGURE 3.6 Life cycle of Dictyocaulus viviparus (Redrafted with permission from Food and Agriculture Organization of the United Nations, Fig. 169, Manual on meat inspection for developing countries http://www.fao. org/docrep/003/t0756e/ T0756E.htm#ch5.4.1.5).
Jarrett and Sharp 1963). The larvae then continue their migration to the lungs via the thoracic duct and heart (Soulsby 1968). The prepatent period in cattle is 21-30 days (Jarrett and Sharp 1963, Soulsby 1968), and the patent period is 27-72 days (Rubin and Lucker 1956).
For these lungworms, transmission to susceptible hosts can have several complications. The definitive hosts are not coprophagic and many avoid feeding near fecal material; thus, once the L1 larvae have passed out with the feces, the larvae must be able to leave the infected fecal pads or pellets to have a reasonable chance of being transmitted to the next host. The best likelihood is associated with the larvae being on vegetation used by the definitive host. While there is some evidence that they are negatively geotactic and weakly climb vegetation (J0rgensen 1980), they also are relatively inactive after being deposited with the feces (Soulsby 1968, Anderson and Prestwood 1981). It is possible that the diarrhea associated with long lush vegetation or gastrointestinal parasite infections (Soulsby 1968) can lead the feces to be deposited directly onto vegetation (Michel and Rose 1954).
An interesting aspect of the transmission of D. viviparus is the involvement of a coprophilous fungus (Pilobolus spp.) to spread L3 away from the feces onto surrounding herbage (Robinson 1962, Croll 1966).
These fungi characteristically discharge their sporangia explosively (Buller 1934). Large numbers of D. viviparus have been observed to migrate to the upper surface of the sporangiophore of Pilobolus spp.; when the sporangia discharged, the larvae were thrown into the air up to distances of 1.2 m under laboratory conditions and up to 3.05 m from fecal pads in the field (Robinson 1962, Doncaster 1981). Cattle pastures with Pilobolus spp. had far more infective D. viviparus larvae than plots without Pilobolus spp. (J0rgensen et al. 1982). Pilobolus spp. also occurs in Yellowstone National Park, and elk (Cervus elaphus nelsoni) have a high D. viviparus prevalence there; thus the same relationship between parasite and fungus may hold here (Foos 1989). While dung beetles have been proposed as a means of mechanically spreading D. viviparus larvae (Poynter and Selway 1966), there also is evidence that dung beetles can reduce the likelihood of infection in hosts by trichostrongyloid nematodes (Fincher 1975, Bergstrom et al. 1976).In some cases the L3 larvae may disappear from the herbage of infected pastures during winter but reappear the following spring or early summer (Duncan et al. 1979). This may be a result of heavy rains washing larvae from the vegetation into the soil; earthworms may contribute to the overwintering survival of D. viviparus larvae by ingesting the previous year's L3 larvae in the soil and returning them to the surface (Oakley 1981).
CLINICAL EFFECTS AND IDENTIFICATION The prepatent phase in the lungs of cattle is associated with blockage of many bronchioles, eosinophilic exudate, and collapse of alveoli; this is associated with rapid shallow breathing and coughing and is sometimes followed with emphysema. Infected black-tailed deer lose weight, have increased coughing, and have an abnormal increase in the depth and rate of breathing (hyperpnea) (Presidente et al. 1973). Among white-tailed deer, gross pathologic lesions attributable to D.
viviparus are confined to the respiratory tract and may range from a foamy exudate in the air passages in mild infections, to a bronchitis and peribronchitis (Goble 1941), to thick purulent exudate in the air passages, and even obstruction of the airways by infecting worms and exudate (Jarrett et al. 1957, Anderson and Prestwood 1981). Opportunistic secondary invaders are common and often may be the eventual cause of death (Anderson and Prestwood 1981); examples include Mannheimia haemolytica and Pasteurella multocida, as well as a variety of viruses and mycoplasma.population effects Dictyocaulus vivipa- rus infections may be relatively common in some deer populations (Prestwood et al. 1971). Among white-tailed deer in New York, D. viviparus lungworms were found in higher prevalence among winter-killed deer, compared to hunter-collected deer in the same area; however, this observation was compounded by three other parasites, including another lungworm [Varestrongylus (= Leptostrongylus) alpe- nae], also having higher prevalences among the winter-killed deer (Cheatum 1951). Thus the relative contribution of D. viviparus was difficult to ascertain. Losses to dictyocaulosis, especially among fawns, may occur annually among white-tailed deer in some areas, and dic- tyocaulosis has been proposed as a population control mechanism in these areas (Anderson and Prestwood 1981), but there are no known systematic studies establishing significant impacts by D. viviparus on wild populations. There have been past reports of D. viviparus having significant effects on black-tailed deer populations in California and British Columbia (Cowan 1946, Longhurst et al. 1952), but no systematic studies on their impacts.
special problems There are many reports of sporadic mortality from D. viviparus. However, there are no clearly established ongoing problems reported for these lungworms.
control Infections in cattle generally occur on over-grazed pastures where the grass is short (Levine 1968).
It has been observed that white-tailed deer in the southeastern U.S. with D. viviparus problems often are associated with areas of poor range condition or which are overpopulated; thus population control and range management are important. Under these circumstances it was proposed that controlled burning in the late winter or early spring may destroy larvae that overwintered on a pasture (Anderson and Prestwood 1981). Keeping range in good condition also would be a sensible strategy—both to reduce the benefit of any upward climbing by lungworm larvae and to keep deer in good health. Among penned black-tailed deer, anthelminthic therapy was not successful (Presidente et al. 1973).Direct Life Cycle Nematodes with Unusual Features
Baylisascaris procyonis
causative agent (classification, morphology) Baylisascaris procyonis (Nema- toda, Family Ascarididae) is a large roundworm of the small intestine (Kazacos and Boyce 1989, Kazacos 2001). The definitive host for adult worms is the raccoon (Procyon lotor); while young raccoons can be infected by ingesting eggs as in a direct life cycle, adult raccoons typically are infected by ingesting infected intermediate hosts or paratenic hosts. Baylisascaris procyonis is one of at least seven species in the genus (Sprent 1968, 1970). Other species in the genus include B. columnaris in skunks, B. laevis in marmots and ground squirrels, B. devosi in martens and fishers (Mustela spp.), B. transfuga in all species of bears, B. melis in European badgers (Meles meles), B. schroederi in giant pandas (Ailuropoda melanoleuca), and B. tasmaniensis in Tasmanian devils (Sarcophilus harrisii) (Sprent 1968, 1970). The genus Baylisascaris is closely related to the genus Ascaris, and to a lesser extent Toxascaris and Parascaris in the family (Zhu et al. 2000).
host range and distribution Baylisas- caris procyonis adults are found primarily in adult raccoons, but also have been found in at least two other species as natural infections, including kinkajous (Potus flavus) (Overstreet 1970) and domestic dogs (Greve and O'Brien 1989); there also are several hosts in which adult parasites can form after experimental infections (Miyashita 1993, Kazacos 2001).
Bay- lisascaris procyonis has a geographic distribution that basically corresponds with the distribution of its main host, the raccoon, and covers primarily the United States; while less common in the southeast (Kazacos 2001, Acha and Szyfres 2003), it appears to be increasing in this region (Eberhard et al. 2003). The parasite occurs among raccoons in parts of Canada and also has been reported among kinkajous of Colombia (Overstreet 1970) and raccoons in zoos or pets in Germany, Poland, Czechoslovakia, and Japan (Miyashita 1993, Kazacos 2001).Among raccoons in the northeastern United States, autumn was the period of highest prevalence of eggs in the feces (Kidder et al. 1989). There also was a significantly higher prevalence of patent infections in juveniles compared to adults (Kidder et al. 1989).
life cycles and variations Raccoons are the normal definitive host, and adult female nematodes in the small intestines of raccoons produce up to 180,000 eggs per worm per day (Fig. 3.7) (Kazacos 1982). Among these ascarid nematodes, the eggs commonly develop infective L2 larvae in 11 to 14 days (Sakla et al. 1989), and these embryonated eggs can remain infective in the environment for years (Kazacos 2001). Infected raccoons can shed feces with over 30,000 eggs per gram, with highest egg counts occurring in the fall and the lowest in winter (Evans 2002b). In young raccoons, L2 lar vae hatching from ingested eggs penetrate and develop within the mucosa of the small intestine for several weeks before entering the intestinal lumen to mature in 50 to 76 days (Kazacos 2001).
A wide variety of birds and mammals, but especially rodents, can serve as intermediate hosts (Tiner 1953, Kazacos 2001, Evans 2002a), in which ingested L2 larvae molt to L3 larvae. Once eggs are ingested, the L2 larvae penetrate the intestinal wall, migrate through the liver to the lungs, and enter the pulmonary veins to gain access to the arterial circulation. The parasite produces a larval migrans, a prolonged migration and persistence of helminth larvae in the organs and tissues of infected intermediate hosts (Beaver 1969). Some larvae become encapsulated in visceral and somatic tissues in eosinophilic granulomas until ingested by raccoons (Kazacos 2001). In older raccoons, L3 larvae from intermediate hosts develop to adults in the intestinal lumen in 32 to 38 days without evidence of any migration through other body tissues (Kazacos 2001).
reservoirs and transmission In the midwestern United States, both the prevalence and intensity of B. procyonis infections are higher among raccoons from rural areas than those in urban locations. Although raccoons may be very common in urban sites, the lower levels of infection may follow from their decreased dependence on infected intermediate hosts as a source of food in urban areas and decreased exposure of young raccoons to B. procyonis eggs (Page et al. 2008).
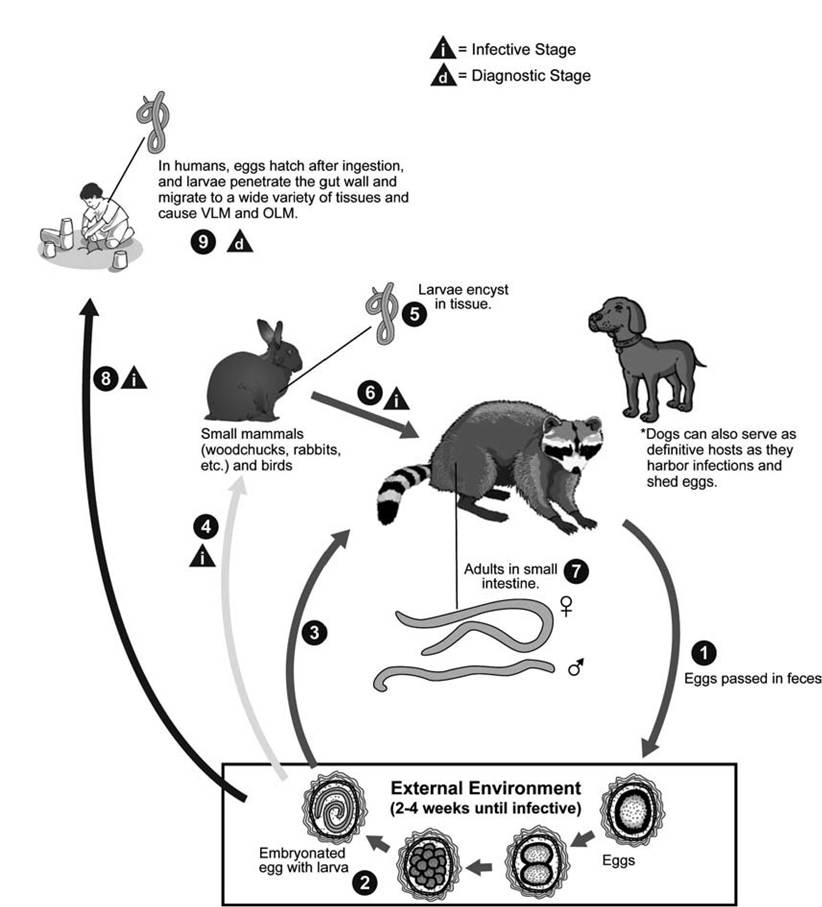
FIGURE 3.7 Life cycle of Baylisascarisprocyonis (Courtesy of Centers for Disease Control and Prevention’s Division of Parasitic Diseases and Malaria, www.dpd.cdc.gov/dpdx).
Transmission to raccoons is by ingesting contaminated nutrients or infected intermediate hosts. Young raccoons become infected at an early age by ingesting eggs from their mother’s contaminated teats or fur, from the contaminated den, or from raccoon latrines near their den (Kazacos 2001). Older raccoons become infected by ingesting L3 larvae in intermediate hosts (Kazacos and Boyce 1989). Transmission among raccoons can be enhanced by altering resource distributions. Prevalences of B. procyonis in raccoons was significantly higher in a habitat with clumped food distributions compared to a similar habitat in which an equal amount of food was dispersed uniformly; the clumped food distribution was postulated to enhance intraspecific contact (Gompper and Wright 2005).
In nature, most transmission to intermediate hosts occurs at raccoon latrines, preferred sites of raccoon defecation where their feces with B. procyonis eggs accumulate (Page et al. 1999). Large numbers of eggs can occur in these latrines, and they can become important long-term sources of infection (Kazacos 2001). Many species of mammals and birds visit these latrine sites, and a considerable number of species actively forage there (Page et al. 1999, 2001b). The development of the central nervous system disease is viewed as a benefit for B. procyonis because the debilitation or death of the host enhances transmission by predation or scavenging so as to reinfect the raccoons (Tiner 1953, Kazacos and Boyce 1989).
CLINICAL EFFECTS AND IDENTIFICATION This nematode is a native parasite that has co-evolved to a benign relationship with raccoons of North America. However, it can be very pathogenic in many of the species that serve as intermediate hosts (Tiner 1953, Sprent 1968, Kazacos 2001, Evans 2002a). Among intermediate hosts, the Ls larvae undergo a larval migrans, causing fatal central nervous system disease, pulmonary hemorrhage, and other somatic disruptions (Tiner 1953, Kazacos and Boyce 1989). Those migrating to the brain produce a progressive central nervous system disease; the onset and severity are dose related (Kazacos 2001). A single larva in the brain of a mouse or small bird usually is fatal (Tiner 1953, Sheppard and Kazacos 1997). Identification of the adults is based on morphological characters and the site of infection (small intestine) of the definitive host (Kazacos 2001).
population effects This parasite has become well known as an important cause of morbidity and mortality in individuals and populations of small vertebrates sharing or frequenting the habitat of infected raccoons (Kazacos 2001). In one assessment, B. procyonis was associated with about 5% of the mortality measured among wild white-footed mice (Pero- myscus leucopus) (Tiner 1954).
Baylisascaris procyonis may have been an important factor in the extirpation of the Allegheny wood rat (Neotoma magister) from parts of its range in the Appalachian Mountains of the eastern United States (McGowan 1993, LoGiudice 2003), linked to the foraging behavior of the woodrat. Woodrats carry whole raccoon feces to their nests and generally do not collect them until they are several weeks old, when the eggs had enough time to embryonate (LoGiudice 2001). LoGiudice also speculated that the nematode’s benign relationship with raccoons, combined with its highly patho - genic effects on the Allegheny woodrat, may have weakened typical density-dependent relationships in which the hosts could avoid being driven to extinction by parasites (LoGiudice 2003).
The abundance of B. procyonis appears to depend more on the density of the raccoon than on that of any one species of intermediate host; thus the raccoon can function as the reservoir for the parasite by continuously reintroducing the parasite to the intermediate host and allowing it to persist in populations that otherwise might be too small to maintain it (Dobson and May 1986, LoGiudice 2003).
special problems This parasite is emerging as an important human pathogen, principally affecting young children. Raccoons increasingly are peridomestic animals living in close proximity to humans and their high densities, which, along with the high prevalence of infective eggs in their feces in raccoon latrines, has led to increased concern about this helminthic zoonotic organism (Sorvillo et al. 2002, Roussere et al. 2003). This is a particularly severe problem among children because the very young can be neurologically devastated despite treatment with anthelminthic drugs and corticosteroids (Gavin et al. 2002).
Translocation to other geographic regions is a potential concern. Although this parasite is found primarily in raccoons of North America, it has been found in mammalian hosts of other countries to which raccoons have been introduced. For example, this parasite caused an outbreak of larval migrans among rabbits (Oryctolagus cuniculus) of a wildlife park in Japan (Sato et al. 2002).
control Under controlled conditions, naturally and experimentally infected dogs harboring B. procyonis had their infections cleared or significantly reduced after administration of milbemycin oxime (Bowman et al. 2005). Decontamination of raccoon latrines in urban settings offers promise for controlling the human risk from this nematode (Page et al. 1999). The diet of raccoons may influence the risk of infection to potential intermediate hosts. For example, white-footed mice (Peromyscus leu- copus) may have higher prevalences of infection when corn is present in raccoon feces (Page et al. 2001b). Likewise, differential foraging strategies can influence the risk of ingesting embryonated eggs among intermediate hosts; white-footed mice tended to select corn particles from fresh feces and had lower risk of ingesting infectious eggs than woodrats, which brought back older, whole fecal pellets to their nests containing embryonated eggs (LoGiudice 2001). Habitat disturbance also can affect the risks to intermediate hosts; prevalence, intensity of infection, and average number of larvae were significantly higher among white-footed mice in a highly fragmented, predominantly agricultural landscape, compared to a homogeneous, predominantly forested landscape (Page et al. 2001a).
Trichinella spp.
CAUSATIVE AGENT (CLASSIFICATION, morphology) Trichinella spp. are members of the Superfamily Trichinelloidea, which all contain a distinctive esophagus with a short muscular anterior portion and a long glandular posterior portion. The genus Trichinella is the sole member of the Family Trichinellidae. The Trichinella species generally are indistinguishable based on morphology (Dick and Pozio 2001), and species identification in much of the earlier literature is not clear. Thus much of the following discussion will be limited to the genus as a whole, with development of species and genotype differences given when adequate information is available.
Trichinella spiralis is well established as a species, and for many years was considered the only accepted species in the genus (Zimmerman 1971). However, the systematics of Trichinella has been considerably complicated by evidence that there are consistent differences in isolates from animals throughout the world where the parasite occurs (Anderson 2000). Based on isoenzymic patterns in isolates of Trichinella spp., from many hosts and sites, there are at least seven distinct electrophoretic clusters (LaRosa et al. 1989, Pozio et al. 1989).
There now are at least 12 taxa, including eight species, with four additional genotypic variants not yet taxonomically defined (Gasser et al. 2004, Pozio 2013). Currently, two main clades are recognized, one comprising species that encapsulate in host muscle tissue and parasitize only mammals, and the other that does not encapsulate and whose species parasitize mammals and birds or mammals and reptiles (Pozio and Zarlenga 2005).
Five species comprise those forming capsules. Trichinella spiralis has a cosmopolitan distribution in temperate and equatorial climatic zones, because it has been passively imported into most continents due to its high infectivity to swine and synanthropic rats; it infects a variety of sylvatic carnivores (Dick and Pozio 2001). It is the species causing most human infections, and it is among the most pathogenic species because of the large number of larvae produced by females (Pozio et al. 1992b); it generates a strong immune response among infected humans (Gomez-Morales et al. 2002).
Trichinella nativa and the closely related genotype T6 infects sylvatic and marine carnivores living in frigid zones of Asia, North America, and Europe; the isotherm -4°C in January seems to be the southern border of this species, and larvae can survive in frozen muscles of carnivores for up to 5 years (Pozio et al. 1992a, Dick and Pozio 2001, Pozio and Zarlenga 2005). Trichinella britovi and the T8 genotype infect sylvatic carnivores of the Palearctic region in Europe, Asia, and also Africa (Pozio et al. 1992a). Trichinella murrelli infects sylvatic carnivores in temperate areas of the Nearctic region (Pozio and LaRosa 2000, Hill et al. 2008). Trichinella nelsoni infects sylvatic carnivores from East Africa to South Africa (Pozio et al. 1992a, Pozio and Zarlenga 2005).
Three additional species do not induce capsule formation during the muscle phase of infection. Trichinella pseudospiralis is a cosmopolitan parasite of both birds and mammals, including marsupials. Trichinella papuae has been found only in Papua New Guinea; it infects both mammals and reptiles, with wild pigs appearing to be the primary reservoir. Trichinella zimbabwensis has been found only in crocodiles and monitor lizards of Africa under natural conditions and has infected several mammals in the laboratory (Pozio and Zarlenga 2005).
HOST RANGE AND DISTRIBUTION Host ranges and distribution have been addressed in the presentation of species above. The genus Trichinella is unique among animal parasite associations because it has at least 150 species of definitive hosts (Campbell 1983). Virtually any mammal can be infected with Trichinella spp. (Dick and Pozio 2001, Pozio and Zarlenga 2005). In addition, T. pseudospiralis can infect birds, and T. papuae and T. zimbabwensis both infect reptiles. Although the various Trichinella species and genotypes have varied distributions, many of the species are cosmopolitan and the genus itself has a broad and worldwide distribution, extending from the Arctic to the tropics, including Oceania (Dick and Pozio 2001, Pozio and Zarlenga 2005). While often considered a zoonotic parasite of domestic habitats, involving pigs, synanthropic rats, and humans, there is evidence that total Trichinella spp. biomass is greater in wild than in domestic animals (Pozio 2013).
There are variations among the various geographic regions. Trichinella nativa and T. spiralis both are reported in Arctic regions. Polar bears (Ursus maritimus) are a key component of transmission in the Arctic and are likely infected by cannibalism, or by scavenging carcasses of Arctic foxes and sled dogs; Trichinella larvae can survive in bears for at least several years (Born and Henriksen 1990, Dick and Pozio 2001). Walruses (Odobenus rosmarus) are another important source; larvae also have been found in bearded seals (Erignathus barba- tus), ringed seals (Phoca hispida), and Beluga whales (Delphinopterus leucas) (Rausch et al. 1956, Madsen 1961, Forbes 2000).
In contrast with the Palearctic region, there are fewer reports of trichinellosis from wildlife in the Nearctic region. Black bears (Ursus americanus) are important in parts of eastern United States and New Brunswick, Canada, as well as throughout the Rocky Mountains, Arctic regions of Canada, and Alaska. Raccoons appear unimportant in central and western Canada, but are important in parts of the United States. Foxes are important in temperate regions with large urban communities and large tracts of agricultural lands as well as in the Arctic. In contrast, small furbearers such as the marten (Martes americana) appear important for transmission in areas with large tracts of relatively intact forest lands. In the Rocky Mountains, cougar (Puma concolor), bears, and perhaps marten are important hosts (Dick and Pozio 2001). Although some of these strains may be T. spiralis, most are identified as Trichi- nella T5 and Trichinella T6 genotypes (Dick and Pozio 2001).
Details on additional geographic regions, including the temperate Palearctic region (T. britovi), the Ethiopic region (probably T. nelsoni), the Southeast Asia and Oceania region, the Central and South American region, are covered in other sources (Dick and Pozio 2001, Pozio and Zarlenga 2005).
life cycles and variations Despite the wide variety of hosts and geographic regions involved, the species and genotypes of Trichinella spp. generally have very similar life cycle strategies. As a model for the other species, Trichi- nella spiralis infective L1 larvae are acquired by ingesting meat containing cysts with larvae of T. spiralis (Fig. 3.8). After exposure to gastric acid and pepsin, the L1 larvae are released from the cysts and invade the small bowel mucosa, where they undergo four molts and develop into adult worms within 36 hours (Ali Khan 1966, Anderson 2000). The females release L1 larvae that invade veins or lymphatics (Anderson 2000)
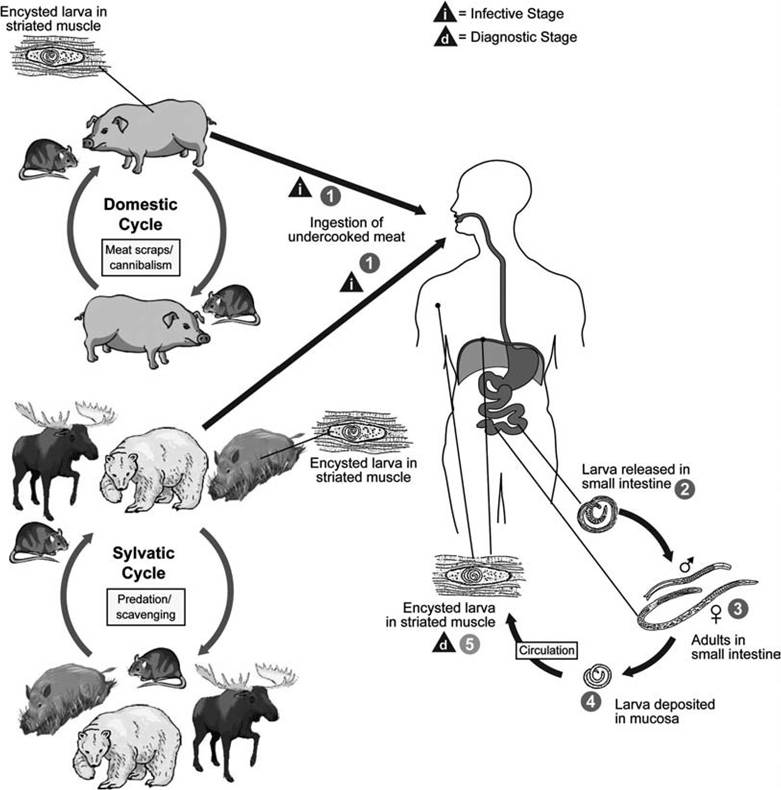
FIGURE 3.8 Life cycle of Trichinella spp. (Courtesy of Centers for Disease Control and Prevention's Division of Parasitic Diseases and Malaria, www.dpd.cdc.gov/dpdx).
and migrate to the striated muscles, where they encyst. The muscles most commonly infected are the diaphragm, larynx, tongue, intercostal, biceps, abdomen, psoas, pectoral, gastrocnemius, and deltoid muscles (Anderson 2000). The infected muscle cell is modified into a “nurse cell,” which functions to nourish the parasite or protect it from host responses (Stewart and Giannini 1982).
The concept of an intermediate host is difficult to apply in the case of Trichinella spp. Trichinelloids always infect the definitive host as L1 larvae (Anderson 2000); in contrast, an L1 larva is considered the infective stage for the intermediate host in almost all other parasitic nematodes requiring an intermediate host. There is no pause in development at the L3 larva stage, the stage commonly viewed as infective to definitive hosts. Thus, in the Trichinella spp. life cycle, the parasite’s definitive host functionally serves as its own intermediate host.
reservoirs and transmission Trichinel- losis is primarily a disease of sylvatic carnivores with cannibalistic and scavenger behaviors (Dick and Pozio 2001, Pozio 2013). While ingestion is established as the route for transmission, some aspects of transmission among marine mammals are not well established. Trichinella nativa in marine mammals has a circumpolar Arctic distribution and a narrow host range (Pozio 2013). It is most commonly found in polar bears and walruses, where it presents a significant zoonotic hazard. Cannibalism probably is the main route for maintaining infections among polar bears (Forbes 2000).
Arctic carnivores as polar bears, foxes, and domestic dogs have high prevalences of Trichinella spp., and the carcasses of at least some of these animals are deposited in the ocean. While scavenging on these carcasses by walruses probably occurs, it likely does not account for the high prevalence of Trichi- nella spp. seen in walruses. Active predation, carrion feeding, and cannibalism have been documented for walruses, and a sylvatic cycle similar to that of bears may exist in walrus populations (Forbes 2000).
Although uncommon, Trichinella is found in seals and whales; these hosts likely are infected through occasional exposure to infected carcasses, either directly by scavenging or indirectly by consuming amphipods or fish that have fed on infected carcasses. The inefficiency of such transmission may help account for the low prevalence of Trichinella larvae in seals and whales (Forbes 2000).
CLINICAL EFFECTS AND IDENTIFICATION There are few observations of clinical effects or pathology of Trichinella spp. in free-living wild animals. Diarrhea, sloughing of gut epithelium, and local hemorrhaging occur in the intestines of experimentally infected domestic and laboratory animals; domestic animals generally tolerate high levels of infection and have few clinical signs (Dick and Pozio 2001). Experimentally infected animals tend to have reduced activity and reduced reproductive success (Dick and Pozio 2001).
The simplest method to recover larvae of Trichinella spp. is to compress pieces of striated muscle from the tongue, diaphragm, or intercostal muscles of mammals between glass slides and view them with a dissecting microscope. One also can digest muscle tissue in 1% HCl/pepsin solution at 37°C for several hours (Dick and Pozio 2001).
Identification of the various Trichinella spp. is complicated. Except for the existence of a capsule and some possible size differences in one of the groups, all species and genotypes of the genus are morphologically indistinguishable at all development stages; thus only biochemical or molecular methods can be used reliably to identify the genotype (Pozio and Zarlenga 2005). A number of methods have been devised, with the polymerase chain reaction (PCR) of single larvae being most common (Zarlenga et al. 1999, Gasser et al. 2004, Pozio and Zarlenga 2005).
population effects While it has been speculated that Trichinella may have some influence on carnivore population densities in Kenya and other parts of Africa (Nelson 1982), there is no clear evidence for any Trichinella spp. causing significant population effects among free- living wildlife.
special problems Emerging problems are related to non-encapsulated species of Trichinella infecting a wide spectrum of hosts (humans, other mammals including marsupials, birds, reptiles), as well as encapsulated Trichinella species infecting horses and other herbivores (Pozio 2001). It is not clear how long these newly discovered host-parasite systems have been established, nor the role humans have played in influencing their importance.
The sylvatic cycle is complicated by the existence of a variety of Trichinella species and genotypes in different regions, and the existence of “new” transmission patterns. Also, game animals have become a more common source of infection for humans in developed and developing countries (Pozio 2001).
control There are no recommended means of control of trichinellosis in free-living wildlife. For domestic animals, the primary control methods are to prevent ingestion of infected meat by susceptible animals; this involves cooking garbage fed to domestic pigs for 30 minutes at 100°C, and rodent control (Fraser and Mays 1986). Use of ACTH and adrenal corticosteroids also can serve as supportive therapy in some cases (Fraser and Mays 1986).
For humans, thiabendazole can be used to treat infections. Likewise, raw pork or products lacking a governmental seal should be cooked at 58°C for about 6 minutes or be frozen at -25°C for 10 to 20 days (Fraser and Mays 1986). However, meat in the Arctic is best treated by cooking rather than freezing (Acha and Szyfres 2003).
Indirect Life Cycle Nematodes