ABOMASAL PARASITES
causative agent (classification, morphology) A distinctive array ofnematodes of the Family Trichostrongylidae lives in the abomasum (fourth chamber of the ruminant stomach) of many ruminants.
Genera of aboma- sal nematodes include Haemonchus, Marshalla- gia, Mazamastrongylus, Obeliscoides, Ostertagia, Pseudostertagia, Spiculopteragia, Teladorsagia, and Trichostrongylus (App. 1: Table 1) At least 21 distinct species have been reported among these genera from wild bovids and cervids of North America alone (Hoberg et al. 2001), with many additional species in other parts of the world (Keep 1971, Wetzel and Rieck 1972, Anderson 1992, Chowdhury and Aguirre 2001). Because many of these nematodes have been associated with diseases and because they often occur in mixed infections among hosts where individual species cannot readily be assessed, they will be addressed collectively. Additional trichostrongyloid nematodes more commonly associated with the small intestine (e.g., Nema- todirus spp., Cooperia spp.) also occasionally are found in the abomasa of ruminants.host range and distribution Aboma- sal parasites typically are found in all ruminant animals, including members of the Bovidae, Cervidae, and Giraffidae. The Camelidae (llamas, camels) have only three chambers in the ruminant stomach, but also are infected with some abomasal parasites (Hoberg et al. 2001). Abomasal parasites have a worldwide distribution (Kotrly and Kotrla-Erhadova 1970, Keep 1971, Wetzel and Rieck 1972, Anderson 1992), although some of this extended distribution may have been influenced by translocations and introductions of hosts and parasites from Europe and other countries (Hoberg 1997). On a worldwide basis, cross-transmission of aboma- sal parasites between wild ruminants and domestic hosts may be common (Dunn 1969), but helminths of wild ruminants to which domestic animals are exposed appear generally to be of low pathogenicity (Hoberg et al.
2001).life cycles and variations Abomasal nematodes have direct life cycles. Eggs are passed by gravid females; the L1 larvae hatch from the eggs (Herman and Wehr 1954) and molt to L2 and then L3 larvae in the pasture (Anderson 2000). The L3 larvae are infective and crawl onto herbage until ingested by their grazing definitive hosts. Some L3 larvae of abomasal parasites can survive in host feces or exposed habitat for 4 months or more (Durie 1961); however, the larvae also commonly are susceptible to high temperatures and desiccation.
Once ingested, L3 larvae exsheath in the abomasum and invade the gastric pits and glands, where they elicit host nodules and swellings in which the larvae develop (Sommerville 1953, 1954); they molt to L4 larvae in a few days. The L4 larvae leave the mucosa as early as 4 days post-infection and enter the lumen to mature or, in some cases, have the fourth molt before returning to the lumen (Anderson 1992). Worms may be either free or attached to the abomasum mucosa. The prepatent period typically is 3 weeks or more (Anderson 1992). However, some trichostrongyloid larvae may undergo a development arrest for several weeks or months in their definitive hosts (Michel 1974, Gibbs 1986).
reservoirs and transmission Infected ruminants are the reservoirs of abomasal nematodes, and susceptible ruminants usually acquire these parasites by ingesting infective L3 larvae contaminating their food. Infective larvae of abomasal parasites have a tendency to climb films of moisture up onto vegetation where they become more readily available to the host (Anderson 1992). Earthworms commonly ingest L3 larvae, which likely pass through the oligochaete intestinal canal to surrounding soil; earthworms also may greatly accelerate the breakdown of feces and contribute to a higher concentration of infective L3 larvae available in the vicinity of the feces (Gr0nvold 1979). The coprophilous fungi Pilobolus spp.
may contribute to the dispersal of some trichostrongyloid larvae (Bizzell and Ciordia 1965); these fungi are addressed in more detail in the discussion of the lungworm, Dictyocaulus viviparus.clinical effects and identification Among the abomasal parasites, Haemonchus contortus can be pathogenic in low numbers and is particularly significant for young deer (Odocoileus spp.) (Davidson et al. 1980). These parasites cause blood loss and damage to the abomasal lining; anemia is a common symptom (Prestwood and Pursglove 1981). Abomasal nematodes typically can cause inflammation, edema, and necrosis of the abomasum, as well as diarrhea and emaciation (Soulsby 1968).
Traditionally, abomasal parasites have been identified through species-specific features of male bursas, including size and number of spicules, presence and shape of a gubernaculums,
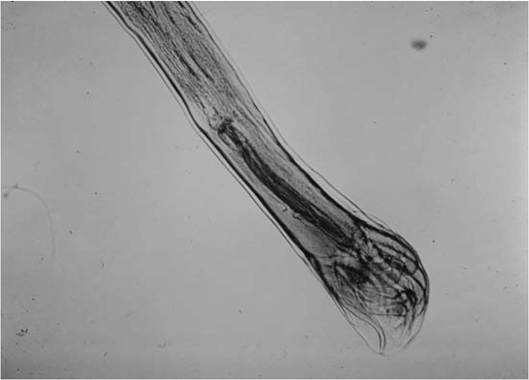
FIGURE 3.5 Male Ostertagia spp. with spicules and curled bursa from a black-tailed deer Odocoileus hemionus californicus.
and numbers and configurations of their rays (Fig. 3.5) (Little et al. 1998). Females can be identified by the structure of their longitudinal cuticular ridge system (synlophe) (Lichtenfels and Hoberg 1993).
population effects Abomasal nematodes have been important mortality factors among white-tailed deer (Odocoileus virginianus) in the southeastern U. S. (Prestwood and Pursglove 1981). Abomasal parasites, along with intestinal nematodes, long have been important mortality factors among California black-tailed deer (Odocoileus hemionus columbianus) along the northern California coast (Longhurst et al. 1952). An estimated 30,000 abomasal nematodes were found in one coastal elk of northern California (Brunetti 1960). However, there are only limited studies on the population impacts of abomasal parasites on any host of a region.
special problems Abomasal parasites have had significant impacts on white-tailed deer in the southeastern United States; in habitats where deer are above carrying capacity, they had significantly higher abomasal counts than at sites where deer populations were at or below carrying capacity (Eve and Kellogg 1977).
Consequently, abomasal parasite counts (APC) of deer were proposed as an indicator of the condition of deer habitat as well as an indicator of the health and nutritional status of a given population of white-tailed deer (Eve and Kellogg 1977, Little et al. 1998).However, conflicting data emerged with these findings. For white-tailed deer in Mississippi (Demarais et al. 1983) and Texas (Waid et al. 1985), abomasal parasite counts were not related to the physical condition of the deer. For black-tailed deer (Odocoileus hemionus columbianus) of coastal and inland Humboldt County, California, there also was no relationship between deer health measures and aboma- sal parasite counts (Botzler 1979); the numbers of abomasal parasite varied consistently between three habitats assessed, but the differences were independent of physical condition of deer from the three habitats. The abomasal parasite counts and physical condition of desert mule deer (Odocoileus hemionus hemionus Rafinesque) from southwestern Texas were not consistently related as they were for white-tailed deer in the southeastern U. S. (Moore and Garner 1980). This was also true for southern mule deer (Odocoileus hemionus fulginatus) in San Diego County, California, which had low abomasal parasite counts despite being in relatively poor physical condition (Ladd-Wilson et al. 2000).
Host age of deer and season have been linked to variation in abomasal parasite counts. Abomasal parasites were most commonly observed among younger animals (Foreyt and Samuel 1979, Davidson et al. 1980) and in late summer and early fall (Eve and Kellogg 1977, Waid et al. 1985).
Soil moisture is another factor that has been evaluated in relation to differences in APC. Arid conditions may influence the abomasal parasite counts. For example, the study sites for whitetailed deer in Texas (Moore and Garner 1980, Waid et al. 1985) and southern mule deer in southern California (Ladd-Wilson et al. 2000) were characterized by dry conditions; such dry conditions may reduce the survival of the free- living phases of these direct life cycle nematodes as they wait to be ingested by the definitive hosts.
Black-tailed deer in Humboldt County, from a cutover redwood temperate rainforest, were intermediate in measures of physical condition but had the highest counts of abomasal parasites; deer from a brushland habitat were in very poor physical condition but had an intermediate level of abomasal parasites (Botzler 1979). Thus, the APC may only be of consistent value for whitetailed deer in the southeastern United States (Eve 1981). Alternatively, specific standards may need to be individually tailored to particular herds, habitats, or geographic regions. However, if the animals are killed for collection, parasite loads may be a less direct method to assess herd health than directly measuring health-related physical or chemical parameters of the deer killed.control Currently, control and treatment remain problematic (Hoberg et al. 2001). While many anthelminthics, including benzimidazoles and avermectins, reduce Haemonchus and other nematodes, few are approved for use in wild cervids and bovids; thus, effective dosages often have not been established (Hoberg et al. 2001). There also are complications associated with attempting anthelminthic therapy in free-ranging ruminants (Prestwood and Pursglove 1981). Haemonchus reportedly is reduced by rafoxanide among wild deer in Europe (Barth and Schaich 1973a, 1973b).
Lungworms
More on the topic ABOMASAL PARASITES:
- In addition to the parasites outlined in this section, there are other parasites that are rarely seen in rats maintained in well-managed facilities.
- CONCEPT 13.2 Hosts have mechanisms for defending themselves against parasites, and parasites have mechanisms for overcoming host defenses.
- Parasites can drive host populations to extinction
- Parasites can influence host population cycles
- APPENDIX ONE Systematics of Major Parasites Groups
- Enslaver Parasites: A Case Study
- Parasites Can Change Ecological Communities
- Germs, parasites, insects and nematodes
- A laboratory experiment tests the role of parasites
- Parasites have mechanisms that circumvent host defenses
- Early observations suggest that parasites cause amphibian deformities
- CONCEPT 13.4 Hosts and parasites can have important effects on each other's population dynamics.