Parelaphostroncylus tenuis (meningeal worm)
CAUSATIVE AGENT (CLASSIFICATION, morphology) The genus Parelaphostrongylus (para: near, elapho: deer, strongylus: nematode) was named based on this genus being similar to another genus, Elaphostrongylus.
Parela- phostrongylus tenuis (Superfamily Metastron- gyloidea, Family Protostrongylidae) is one of three members of this genus (Lankester 2001). Earlier, this parasite was called Pneumostrongy- lus tenuis, but was reclassified into its current genus about 1971. Two other closely related species, P. odocoilei and P. andersoni, also occur in North America; both are called “muscleworms” because the adult nematodes are found in striated muscle of the deer definitive hosts.host range and distribution Parela- phostrongylus tenuis is a neurotropic nematode of white-tailed deer (Anderson 2000) found in most regions of the eastern United States and southern Canada (Lankester 2001). Generally, this parasite is common among deer of the deciduous forest biome and deciduous- coniferous ecotones of eastern and central North America. It is rare or absent in the coastal plains region of the southeastern United States and has not been documented in western North America (Lankester 2001). Its distribution has been extended into parts of Europe, including Holland, Scotland, Germany, the Czech Republic, Sweden, Finland, Russia (Ural Mountains), central Asia, and south Siberia, through white-tailed deer introductions (Horning 1975).
White-tailed deer are considered the normal definitive host of this parasite (Lankester 2001). However, in North America, the parasite also has been described in mule deer and blacktailed deer (Odocoileus hemionus) (Anderson et al. 1966, Nettles et al. 1977a, Tyler et al. 1980) elk and red deer (Cervus elaphus) (Carpenter et al. 1973), moose (Alces alces) (Karns 1967), caribou and reindeer (Rangifer tarandus) (Anderson 1971), and fallow deer (Dama dama) (Nettles et al.
1977b). In Europe, this parasite also is reported from free-living fallow deer, red deer, reindeer, and moose that have come in contact with introduced infected white-tailed deer (Horning 1975). In addition, infections have been reported in a wide variety of bovids and other ungulates in zoos or game farms situated on white-tailed deer habitats, or through experimental infections (Anderson 1992, Lankester 2001) However, the parasite rarely develops to adulthood or is able to pass infective larvae in hosts other than white-tailed deer (Lankester 2001).life cycles and variations In whitetailed deer, the usual definitive host, the adult worms most frequently are found in the veins and venous sinuses of the cranial meninges (Lankester 2001). Unembryonated eggs released by females into the venous blood are carried to the lungs, where they lodge in alveolar capillaries and hatch L1 larvae (Fig. 3.9). These L1 larvae move into the mucous of alveolar air space and are carried up the lungs by cilia to the pharynx, where they are swallowed and pass through the digestive tract and out with the feces (Anderson 1963). The L1 larvae tend to concentrate in a thin layer of mucous on the surface of fecal pellets (Lankester and Anderson 1968) and can be detected by a Baermann apparatus or other living larvae technique. To develop further, L1 larvae must penetrate or be eaten by one of many terrestrial snails or slugs that can serve as intermediate hosts (Lankester 2001). The L1 larvae molt to L2 and then to L3 larvae in the
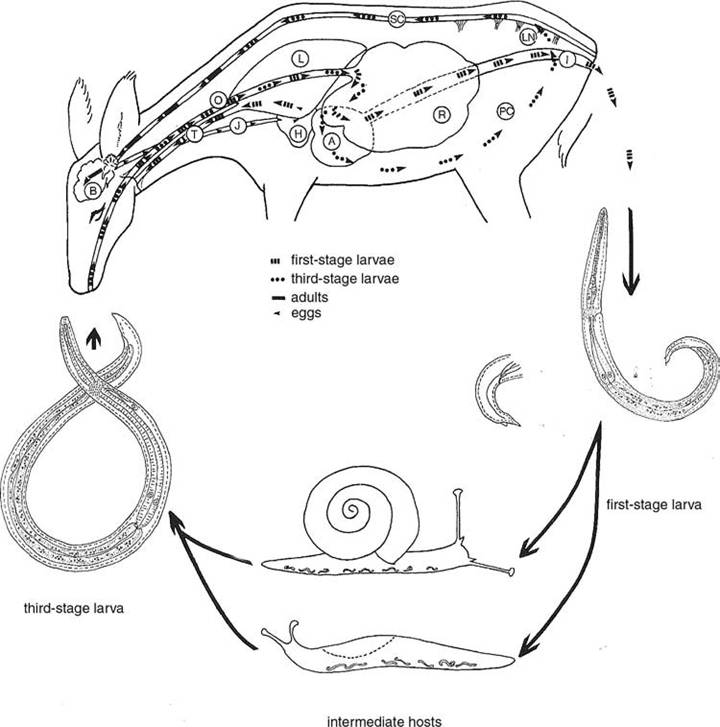
FIGURE 3.9 Life cycle of Parelaphostrongylus tenuis. A. abomasum. B. brain. H. heart. I. intestine. J. jugular vein. L. lung. LN. lumbar nerve. O. esophagus. PC. Peritoneal cavity. R. rumen. SC. Spinal cord. T. trachea (Anderson, 1992; Courtesy of Uta Strelive, copyright holder).
foot of the gastropod over a period of several weeks.
Development is influenced by temperature and the metabolic rates of the gastropods, and the L3 larvae probably remain viable for the life of the intermediate host (Lankester and Anderson 1968).White-tailed deer are infected by ingesting infected gastropods. The L3 larvae penetrate the deer abomasum and other parts of the intestinal tract and migrate through the peritoneal cavity to the central nervous system (Anderson 1963, Lankester 2001). The L3 larvae enter the dorsal horns of gray matter in the spinal cord, molt to the L4 and then to the subadult stage as they move up the spinal subdural space. On reaching the head, they enter the venous sinuses (Lankester 2001). The prepatent period of P. tenuis in white-tailed deer varies from 82 to 137 days (Anderson and Prestwood 1981). The worms are long-lived in white-tailed deer and may persist for the life of the host (Slomke et al. 1995, Lankester 2001).
reservoirs and transmission The whitetailed deer is the usual definitive host, although moose and elk occasionally can become patent and shed some larvae (Lankester 2001). However, it is not evident that the parasite can be established in any geographic region without the presence of the white-tailed deer as the reservoir host.
Two key points of transmission include the L1 larvae moving from infected deer feces to the gastropod intermediate host, and then the L3 larvae from infected snails being ingested by a susceptible definitive host. The ability of L1 larvae to withstand adverse natural conditions and remain infective to gastropods has not been thoroughly studied (Lankester 2001). Although they can be kept frozen in the laboratory for several months (Lankester and Anderson 1968), repeated freezing and thawing greatly reduces survival of the L1 larvae (Shostak and Samuel 1984). The larvae also do not survive well beneath snow (Forrester and Lankester 1998).
There is some inconsistency on whether transmission is more likely in specific sites within a white-tailed deer range (Lankester and Anderson 1968, Maze and Johnstone 1986, Platt 1989).
Where distinct foci of infection seem to occur, distribution of infected gastropods and young deer may be contributing factors to the distribution of the parasite (Lankester 2001).CLINICAL EFFECTS AND IDENTIFICATION In white-tailed deer, the parasite generally causes no clinical signs or pathology (Anderson 1992). However, considerable pathology can occurs in other species that serve as abnormal hosts. The parasite is highly pathogenic in moose, in which it causes a neurological disease (“moose sickness”); clinical effects include lumbar weakness, ataxia, torticollis, blindness, fearlessness, depression, paresis, paraplegia, and death (Anderson 1992). The parasite also is highly pathogenic in caribou (Trainer 1973) and reindeer (Anderson 1971). While elk and red deer are susceptible to neurologic disease, they can tolerate the infection and sometimes even pass L1 larvae (Anderson 1992). The parasite also is pathogenic to mule deer, and partially pathogenic to mule deer and white-tailed deer hybrids (Anderson et al. 1966, Nettles et al. 1977a, Tyler et al. 1980).
In abnormal hosts, the parasite causes neurological symptoms and death through meningitis (inflammation of the meninges), encephalitis (inflammation of the brain), and meningoencephalitis. The severity of infection in abnormal hosts probably is due to the higher proportion of invading worms that reach the central nervous system, their longer developmental period in the spinal cord, their resulting larger size and coiling behavior, and frequent invasion of the membranes lining the ventricles of the brain (Anderson 1968).
Recovering and identifying adult parasites from infected hosts currently is the only way to confirm infection with P. tenuis. However, the first stage larvae passing out with the feces have a distinctive dorsal cuticular spine on the tail, and this often is strong evidence for infection by P. tenuis; such larvae are detected with a Baermann apparatus or other living larvae technique.
There is interest in developing blood tests using immunological and molecular techniques (Dew et al. 1992, Ogunremi et al. 1999).Reported prevalence in white-tailed deer ranges widely, from 1 to 94% (Lankester 2001). However, many of the lower estimates may be biased by a lack of care or skill in finding adult worms in venous sinuses, inclusion of traumatized heads that cannot be completely examined, or sampling young infected animals in autumn before the adult worms mature in the head or L1 larvae are produced (Lankester 2001). Prevalences in infection based on L1 larvae in feces also may reduce estimates of true prevalence (Lankester 2001).
population effects Historically, some authors associated marked declines in moose populations and reports of sick moose with incursions by white-tailed deer into these regions; they proposed that P. tenuis brought with invading deer might have caused the moose sickness and population declines (Anderson 1972, Gilbert 1974). However, the idea that white-tailed deer were preventing moose from becoming established in the eastern United States and Canada was challenged (Nudds 1990). Later, using available data on deer and moose population dynamics in several sites, Nudds argued that, with the available information, meningeal worms could contribute to moose declines, but that moose could coexist with deer, at lower densities, even in the absence of habitat refuges from meningeal worm (Schmitz and Nudds 1994). In a retrospective study of moose sickness, there was not adequate data to support a relationship between white-tailed deer densities and moose declines (Whitlaw and Lankester 1994). In another study of a declining moose population in Minnesota, meningeal worms were assessed to be a mortality factor of less impact than liver flukes; while there was no relationship between annual population growth and deer abundance, the authors did view deer as potentially serving as reservoir hosts for parasites for the moose (Murray et al.
2006). The role of this parasite as a source of moose decline still is controversial.In contrast, meningeal worms clearly have been associated with the failure of several caribou or reindeer introductions in areas with white-tailed deer (Anderson 1971, Trainer 1973). Based on a review of 33 reintroduction attempts in eastern North America, no caribou reintroductions were successful where whitetailed deer had a high frequency of meningeal worm infection (Bergerud and Mercer 1989).
special problems Parelaphostrongylus tenuis is reported from free-living fallow deer, red deer, reindeer, and moose that have come in contact with introduced infected white-tailed deer in parts of Europe, including Holland, Scotland, Germany, the Czech Republic, Sweden, Finland, Russia (Ural Mountains), central Asia, and south Siberia (Horning 1975). The parasite causes a meningoencephalitis where these free-living cervids are in contact with introduced whitetailed deer populations (Horning 1975).
control At doses of 0.1 mg/kg, ivermectin did not prevent infection or eliminate P. tenuis in white-tailed deer, but routine use of ivermectin may temporarily reduce the number of larvae shed in the feces of infected deer (Kocan 1985). However, there is some evidence that an acquired or concomitant immunity follows low-dose infections in white-tailed deer (Slomke et al. 1995) and fallow deer (Davidson et al. 1985).
Filarial Nematodes
Filarial nematodes are in the Order Spirurida, Superfamily Filarioidea, Family Onchocerci- dae, (Anderson 2000). Filarioid nematodes are highly specialized parasites of tissues and tissue spaces of vertebrates other than fish. Mature filarial nematode females are ovoviviparous or viviparous and produce modified L1 larvae called microfilariae that enter the blood or lodge in the skin of the definitive host (Lee 2002), where they are available to hematophagous arthropods that serve as intermediate hosts or vectors (Anderson 2001). Elaeophora schneideri, a parasite of wild ungulates, will serve to illustrate this group.
Although not addressed further in this text, a number of other species affect wildlife. Dirofilaria immitis, the canid heartworm, is transmitted by mosquitoes; although primarily described for domestic dogs, there are increasing numbers of cases being observed in wild canids and a few other species (Abraham 1988). There are a number of other poorly studied but potentially interesting filarial worms of wild mammals and birds (Anderson 2000).
More on the topic Parelaphostroncylus tenuis (meningeal worm):
- ELAEOPHORA SCHNEIDERI (ARTERIAL WORM)
- AMIDOSTOMUM ANSERIS (GIZZARD WORM)
- STOMACH AND INTESTINAL WORMS
- CONTENTS
- Bibliography
- Cryptococcosis
- Taenia taeniaformis Infestation
- Parasites can influence host population cycles
- PARASITIC CAUSES
- Enslaver Parasites: A Case Study
- phylum Nematoda
- CLINICAL EVALUATION OF GIT DISEASE
- FILARIASIS
- INTRODUCTION TO PHYLUM Platyhelminthes
- Boon Andrew. The Ethics and Conduct of Lawyers in England and Wales. Hart Publishing,1999. — 808 p., 1999
- Griffiths-Baker Janine. Serving Two Masters: Conflicts of Interest in the Modern Law Firm. Hart Publishing,2002. — 227 p., 2002
- Grisso T.. Evaluating Competencies: Forensic Assessments and Instruments. 2nd edition. — Springer,2002. — 564 p., 2002
- Luban David. Legal Ethics and Human Dignity. Cambridge University Press,2007. — 350 p., 2007