phylum Nematoda
Introduction
Members of the Phylum Nematoda (Greek “nema” = thread) are bilaterally symmetrical worm-like metazoans that are round in cross section. The overall body shape of a cylinder tapered at both ends tends to be uniform among most nematode species (Bush et al.
2001) (Fig. 3.1). While lacking a body cavity, nematodes have a fluid-filled false cavity without a peritoneal lining (pseudocoel) between the somatic musculature and digestive tract that serves as a primary component of the hydrostatic skeleton, and thus is critical for movement; it also serves to transport nutrients between tissues (Bush et al. 2001). Nematodes are covered by a non-cellular cuticle that allows selective permeability and helps maintain their shape by hydrostatic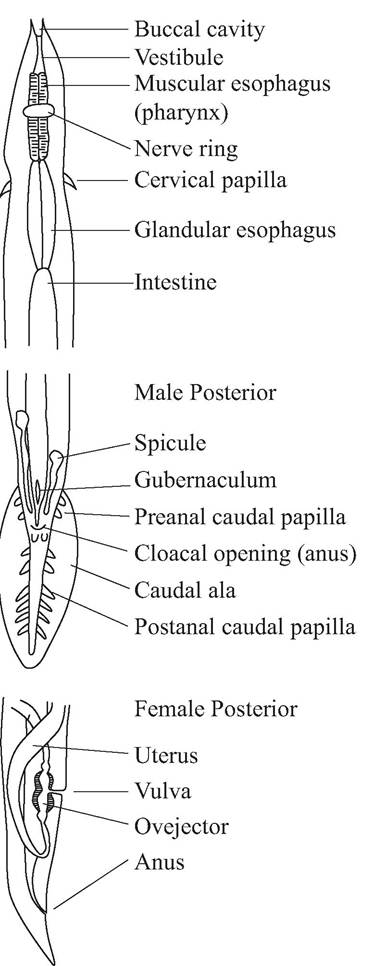
FIGURE 3.1 Generalized anatomy of male and female nematodes (McDonald, 1974; Courtesy of Malcolm McDonald and the U. S. Fish and Wildlife Service).
pressure. With rare exceptions they have separate sexes (dioecious).
Nematodes are not segmented, and have a complete digestive tract, including both a mouth and anus (Roberts and Janovy 2000). This is in contrast to cestodes and acantho- cephala, which have no intestinal tracts and absorb food through their cuticles; trematodes have bilateral blind ceca.
A buccal capsule lined by a cuticle is present in some nematodes. The esophagus of nematodes is glandular and muscular, and lined by a cuticle. There are several types of esophagi among nematodes. Filariform esophagi have parallel muscular walls and are the most common. Strongyliform esophagi are bulbous. Rhabditi- form esophagi are bulbous, with an indentation on the bulb to allow a nerve to cross over.
Nematodes can be distinguished from other pseudocoelomate groups by the possession of spicules among males and a ventral excretory pore (Roberts and Janovy 2000).
Taxonomy typically is based on the copulatory bursa with associated spicules and rays in males (Bush et al. 2001), the location of the vulvar opening in females, esophageal shape, the anterior adornments (e.g., buccal capsule or lips), and characteristics of cuticular bosses and ridges (Fig. 3.1).Although there is some variation in taxonomic schemes proposed for nematodes (Adamson 1987; Anderson 1992, 2000; Roberts and Janovy 2000), we use the work of Anderson (2000), which is based on the Commonwealth Institute of Helminthology (CIH) keys (Anderson et al. 1974-1983), as the foundation for a classification (App. 1: Table ι). However, biochemical, immunological, and molecular techniques increasingly are providing additional and alternative approaches for studying parasite systematics (McManus and Bowles 1996, Andrews and Chilton 1999).
Nematodes have more species that cause mortality among wildlife than any other helminth group. Approximately 16,000 nematode species are described, and about 40,000 species are estimated to exist (Anderson 2000). About one-third of all nematode genera described occur as parasites of vertebrates (Anderson 1984) and many others are important as insect or plant pathogens (Bush et al. 2001). However, many nematodes are free-living and play important roles as decomposers in recycling nutrients, or in food web relationships by feeding on bacteria and other microorganisms
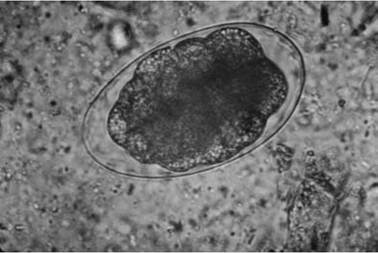
FIGURE 3.2 Ovum of Ostertagia spp., an abomasal nematode of cervids.
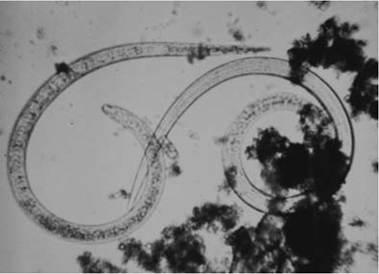
FIGURE 3.3 First stage (Li) larvae of Trichinella spp. being digested out of bear muscle (Courtesy of R. M. Wood and California Department of Public Health).
(Bush et al. 2001). In one assessment, about 24,000 parasitic species of nematodes were estimated to exist (Poulin and Morand 2004). A number of pathogenic nematodes are described for both birds (Wehr 1971) and mammals (Samuel et al.
2001). Among waterfowl (Family Anatidae) alone, over 200 species of nematodes in more than 50 genera have been reported (McDonald 1974); of these, about 20 species are reported to be pathogenic (Wobeser 1997).Nematodes have several distinct life history stages, including an egg (Fig. 3.2), four larval stages (Fig. 3.3), and adults of separate sexes (Fig. 3.4). The larval and adult stages are separated by four molts (Bird and Bird 1991).
After adult nematodes mate, the eggs develop first stage larvae (L1), which successively molt into second stage larvae (L2), third stage larvae (L3), fourth stage larvae (L4), and then mature into adults. In the great majority of nematodes parasitizing vertebrates, the L3 larvae are infective to the definitive vertebrate host (Anderson 2000). Some exceptions of stages infective to the definitive host occur with many ascarid nematodes, in which the L2 larva is infective, Trichinella (L1 is infective), and Eustrongylides (L4) (Anderson 2000). There is no reproduction among any larval stages of nematodes.
Some nematodes have a direct life cycle (monoxenous), without a need for any intermediate hosts. Other nematodes, especially among vertebrates, have an indirect life cycle (heteroxenous), with development from the L1 to the L3 requiring intermediate hosts (Anderson 2000). Some advantages of an intermediate host are that the pre-infective stages of nematodes are protected from the uncertainties of the external environment in a nutrient-rich environment and the survival of larvae can be extended in time and space; also, the intermediate host often is a desired prey item of the final host and thus increases the likelihood of the parasite reaching its definitive host (Anderson 2000). Although relatively uncommon, some nematodes have paratenic hosts in which the infective stage of a parasite persists without essential development and usually without growth in a host (Anderson 2000).
While there are some larval changes in an intermediate host, there are no larval changes in a paratenic host. Nematodes using paratenic hosts include metastrongyloid lungworms of mustelids and members of the order Spirurida (Anderson 1992).Definitive hosts can be infected in a variety of ways. Ingestion of an infective larval stage is the most common means. Ingestion can be of infected intermediate hosts, paratenic hosts, or food or water in fecally contaminated environments. Penetration of skin is another means of transmission. Some infective nematode larvae can penetrate directly through the skin of the definitive host; hematophagous arthropods also can serve to infect a definitive host with infective larvae through the skin. Less commonly, transmammary transmission can occur to nursing young; transmission also can occur by ingestion
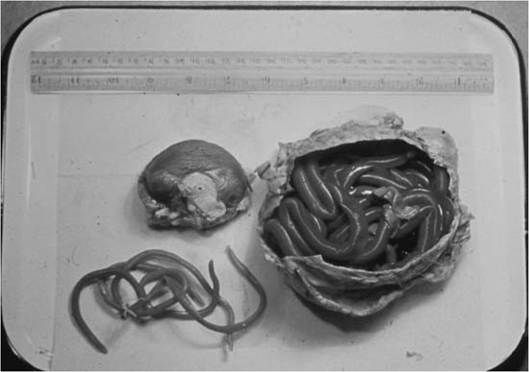
FIGURE 3.4 Adult kidney worms (Dioctophyma renale) from a coyote (Canis Iatrans). Note the size of the animal's uninfected kidney on the left (Courtesy of O. Brunetti and California Department of Fish & Wildlife).
of emesis. Transplacental transmission is an occasional mechanism as well (Anderson 1992).
In this chapter, we will illustrate some of the variation found among nematode life history strategies, with a focus on species of concern to managers. The approach initially will be by life cycle type, starting with direct life cycles and then focusing on nematodes with indirect life cycles. Coverage will include important intestinal nematodes, lung worms, and vascular system nematodes.
Direct Life Cycle Nematodes
Intestinal nematodes
More on the topic phylum Nematoda:
- phylum Acanthocephala
- INTRODUCTION TO PHYLUM Platyhelminthes
- PHYLUM PENTASTOMA
- CONTENTS
- CONTENTS
- BIBLIOGRAPHY FOR PARASITIC DISEASES
- CRYPTOSPORIDIUM
- Encephalitozoon cuniculi Infection: Encephalitozoonosis; Microsporidiosis
- INTRODUCTION TO PROTOZOA
- LEECHES (ANNELIDA: HIRUDINEA)
- GENERAL INTRODUCTION
- Boon Andrew. The Ethics and Conduct of Lawyers in England and Wales. Hart Publishing,1999. — 808 p., 1999