CRYPTOSPORIDIUM
Cryptosporidium is a genus of enteric protozoans within the protist phylum Apicomplexa.25'26 At least 10 species within the genus (C. parvum, C. muris, C. wrairi, C.
felis, C. meleagridis, C. baileyi, C. serpentis, C. nasorum, C. saurophilum, and C. andersoni) are currently recognized on the basis of differences in host specificity and oocyst morphology.27'28 Immunocompromised individuals, such as patients with acquired immunodeficiency syndrome (AIDS), can be infected with most of the species, including C. parvum, C. felis, C. muris, and C. meleagridis.27-29 C. parvum, the most common species that infects AIDS patients, contains eight chromosomes with a 10.4-Mb haploid genome, of which a complete map was recently completed and two distinct genotypes—human type 1 and bovine type 2—were identified to infect humans.30,31 Humans are infected by ingesting cryptosporidium oocysts. After being ingested, oocysts excyst in the gastrointestinal tract, releasing infective sporozoites that, in turn, invade intestinal epithelial cells. Soon after attachment, cryptosporidium induces protrusion of the host cell membrane to cover the parasite and resides in an intracellular but extracytoplasmic parasitophorous vacuole. In this parasitophorous vacuole, the parasite undergoes further intracellular development through asexual as well as sexual cycles, which eventually leads to formation of new oocysts.27,32,33 Although most oocysts formed are thick-walled and are excreted from the host in fecal material, some oocysts are thin-walled and can excyst within the same host, which leads to a new cycle of development.27Epidemiology of Cryptosporidial Infection in Humans
Human cryptosporidium infection is distributed in urban and rural populations in both developed and developing countries and on all six continents.
Oocyst excretion rates for the general population vary from 1 to 3% in industrialized countries and can reach 10% in developing nations.33 Cryptospo- ridial infection accounts for 2.2% (0.26 to 22%) and 6.1% (1.4 to 40.9%) of diarrhea in immunocompetent persons in industrialized and developing countries, respectively.34 Infection is more common in immunocompromised subjects, especially those with AIDS.33 In the pre-HAART (highly active antiretroviral therapy) era, 3 to 4% of AIDS patients in developed countries and as many as 50% of AIDS patients in the developing world were infected. Since the introduction of HAART, the incidence of cryptosporidial infection has fallen in the developed world.32,35-37 However, it is still a clinically significant problem in AIDS patients who do not have access to HAART.32Cryptosporidial Clinical Manifestations
The intestinal tract is the primary site of cryptosporidiosis. The duration and severity of clinical symptoms of intestinal cryptosporidiosis depend largely on the immune status of the infected individual. Cryptosporidiosis in otherwise healthy individuals is usually a self-limiting illness with a median duration of 9 to 15 days.38 Although infection can be asymptomatic, the most common clinical manifestation is profuse watery diarrhea containing mucus but rarely blood or leukocytes. Other symptoms include nausea, vomiting, cramp-like abdominal pain, and mild fever.38 In AIDS patients, a wide spectrum of disease symptoms are present, from asymptomatic shedding of cryptosporidial oocysts to a fulminant cholera-like illness, which result in significant morbidity and mortality. The diarrhea in AIDS patients is usually watery, and stool frequency can be as much as 10 times a day with a mean volume of 1 liter; these patients can experience a 10% drop in body weight and usually develop severe malabsorption. Most never clear the infection and ultimately have a shorter survival life than AIDS patients without cryptosporidiosis.37,39,40 Several studies have shown a significant relationship between the immune status of AIDS patients and the severity of the diarrheal disease.
Four clinical patterns of disease have been identified: asymptomatic carriage, transient infection (diarrhea lasts for less than 2 months followed by a complete remission of symptoms and loss of cryptosporidium from fecal specimens), chronic infection (diarrhea lasts for 2 months or more with persistence of the parasite in the stool and biopsy specimens), and fulminant infection (patients pass at least 2 l of watery stool daily).38 Chronic infection is the most common presentation (59.7%), followed by transient (28.7%), fulminant (7.8%), and asymptomatic (3.9%) infection. Fulminant infection occurs only in patients with CD4+ T cell counts of or Fas-sensitive Jurkat cells undergo apoptosis via a Fas receptor/Fas ligand-dependent pathway when co-cultured with infected biliary epithelia. Additionally, caspases, a family of intracellular cysteine proteases and downstream effectors of the Fas receptor,52 were implicated in the molecular machinery responsible for apoptosis.1 Furthermore, blocking apoptosis with the caspase inhibitor increased the percentage of infected cells, suggesting that infected cells may attempt to eliminate the parasite through apoptosis.47Nuclear factor-κ B (NF-κB) activation was shown to prevent apoptosis of TNF-α-stimulated cells in several experimental models.53 Early studies demonstrated that C. parvum infection of intestinal epithelial cells is paralleled by the activation of NF-κB target genes.54-56 More recently, McCole et al.49 indicated that C. parvum-induced apoptosis was limited by the concomitant activation of NF-κB. Using an in vitro model of biliary cryptosporidiosis, we found that nuclear translocation of the NF-κB family of proteins (p65 and p50) was observed in cells exposed to C. parvum, and activation of NF-κB was found in infected cells but not in bystander uninfected cells. In addition, inhibition of NF-κB activation resulted in apoptosis in infected cells and significantly enhanced C.
parvum-induced apoptosis in bystander uninfected cells. These observations support the concept that although C. parvum induces host cell apoptosis in bystander uninfected host cells, which may limit spread of the infection, it directly activates the NF-κB∕I(κ)B system in infected biliary epithelia, thus protecting infected cells from death and facilitating parasite survival and propagation.Synergistic Effects of C. parvum Infection with HIV-I-Associated Soluble Factors to Induce Epithelial Cell Apoptosis
There is, as yet, no evidence to support a role for direct infection of the biliary epithelium by the HIV virus. Previous studies have shown that cholangiocytes do not express CD4, CXCR4/5, and galactosylceramide, the receptors for HIV direct infection or injury.57 No direct HIV-1 infection of cholangiocytes in humans has ever been reported. As mentioned above, AIDS-cholangiopathy is now recognized as a result of opportunistic infections within the biliary tree, and in most series, cryptosporidium is the single most common identifiable pathogen. Moreover, immunocompromised patients, because of profound immunosuppressive therapy, do not appear to be affected by biliary cryptosporidiosis; thus, HIV-infected patients appear to be uniquely susceptible. However, in our lab and in others, cholangiocytes were successfully infected with C. parvum in vitro,58,59 suggesting that HIV infection is not a prerequisite for C. parvum infection of cholangiocytes. Therefore, synergistic pathologic effects in response to dual infection may exist.
Recent studies indicate that HIV infection of T cells can promote release of a number of HIV-1- related biologically active peptides (e.g., TAT, gp120, Nef, and Vpr).60-62 Of particular importance is the discovery that HIV-1 TAT protein, a 16 kDa peptide essential for viral replication, is released in a biologically active soluble form and can be taken up by many cell types by an unknown mechanism.63 Furthermore, TAT was reported to induce cytokine release and to mediate apoptosis in T cells, monocytes, and endothelial cells.64-68 TAT was reported to induce apoptosis in many cell types.
It was reported that TAT stimulates matrix metalloprotease activity in monocytes in vitro.69 TAT does not seem to upregulate FasL directly but rather synergies with other signaling pathways (e.g., CD4 dependent) to regulate FasL expression.69 Most recent studies indicate that TAT upregulates caspase-8 in epithelial cells, a downstream enzyme system responsible for Fas- associated apoptosis,70 and thus makes cells more susceptible to Fas/FasL-associated apoptosis. Therefore, TAT could serve two functions to influence Fas/FasL-mediated apoptosis: to increase susceptibility via caspase-8 upregulation and to enhance FasL production at the transcriptional level. We found that when cholangiocytes were exposed to C. parvum in the presence of recombinant biologically active HIV-1 TAT protein at a concentration (100 ng/ml) comparable to serum levels in AIDS patients,71 a significant increase of cholangiocyte apoptosis was found.58 In contrast, nochange of the infection rate of C. parvum to Cholangiocytes was found in the presence of TAT. Thus, it seems that HIV-1 will influence C. parvum-induced cholangiocyte apoptosis with C. parvum via TAT by modulation of the Fas/FasL apoptotic pathway and thus will act synergistically to cause AIDS cholangiopathy.
Diagnosis
Definitive diagnosis of cryptosporidium infection requires microscopic detection of the parasite in tissue or in body fluids, because cryptosporidiosis does not produce a specific clinical syndrome. However, clinical, endoscopic, immunologic, and molecular techniques all have a place in the diagnosis and clinical assessment of human cryptosporidiosis.38 The identification of oocysts in stool with the aid of special stains, such as the modified acid-fast stain, or with the assistance of immunofluorescent labeled antibodies is the approach most commonly used in diagnostic laboratories. Polymerase chain reaction (PCR)-based techniques are available as research tests.
Serology is of limited value, because many healthy individuals already have antibodies to C. parvum. Endoscopic appearances in the intestines are usually normal, although in cases of severe partial villous atrophy, there may be a diminution in the fold pattern in the duodenum.When biliary disease is suspected, ultrasonography is the best initial diagnostic test. However, the most sensitive test to use to diagnose biliary tract disease in HIV-infected patients is endoscopic retrograde cholangiopancreatography (ERCP).
Therapy
The treatment of cryptosporidiosis is far from satisfactory. There is no antimicrobial chemotherapeutic agent that will reliably eradicate the organism. A number of agents seem to have some activity against the organism, although they are usually suppressive and not curative. Paromomycin, azithromycin, and most recently, nitazoxanide are commonly used.72,73 However, as these antipara- sitic drugs are, at best, modestly effective, preliminary clinical results using these agents are not uniform and well-designed placebo-controlled studies, including assessment of drug combinations, are needed to confirm which, if any, agents are effective.73-76
Because the clinical course of cryptosporidiosis in immunocompromised individuals depends largely upon the immune status of the host, treatment options for these patients should aim to resolve the immune system function and to lead to resolution of clinical symptoms. The best treatment for cryptosporidiosis in AIDS patients is HAART.32,35'37,74 It can partially restore host immune function and resolve cryptosporidium infection. Some nucleoside antivirals have a direct effect on growth of C. parvum in vitro.77 Therapy of biliary cryptosporidiosis in AIDS patients is dependent on HAART, whereas endoscopic sphincterotomy may provide symptomatic relief for patients with abdominal pain or cholangitis associated with papillary stenosis, as it facilitates drainage and decompression of the biliary system and thus improves quality of life.36,78 Patients with diffuse intra- and extrahepatic sclerosing cholangitis alone have few specific treatment options.
epstein-barr virus
Epstein-Barr virus (EBV) is a human Gammaherpesvirinae that infects B lymphocytes. EBV is ubiquitous, and up to 95% of the adult population has been infected with it. Most infections are acquired through the orthopharynx, usually during childhood, and do not seem to cause any disease. However, when immunocompromised, EBV can favor the initiation of a number of malignancies. The EBV genome is linear and composed of double-stranded DNA of more than 170,000 base pairs. The DNA is contained in an icosahedral capsid, surrounded by a loosely packed tegument and finally by an outer envelope coated with glycoprotein spikes. When it is introduced to a host, infection occurs when the EBV gp350∕220 binds to the B lymphocyte surface receptor CD 20.79 After infection, either the viral genome is incorporated into the host cell’s chromosomal DNA or the viral genome is circularized and exists as an episome.80
EBV Clinical Manifestations
The majority of human infections with EBV are asymptomatic. However, in different contexts, EBV has been implicated in the pathogenesis of Hodgkin’s lymphoma (HL), Burkitt’s lymphoma (BL), posttransplant proliferative disorder (PTLD), primary effusion lymphoma (PEL), and nasopharyngeal carcinoma (NPC). HIV patients and other immunocompromised individuals have a much greater susceptibility to EBV-related infections, because the T lymphocytes are not able to restrict the virus to a latent pool of B lymphocytes.79 In most circumstances, lymphomas that result from EBV infection are restricted to the B lymphocytes, but there are cases in which the EBV- associated malignancies occur in T lymphocytes or NK cells.81
EBV infection can either exist in the lytic form or develop into one of three latency phases. Most frequently, the latency seen in vitro is type III, which is that characterized by expression of nuclear antigens EBNA-1, EBNA-2, EBNA-3A, EBNA-3B, EBNA-3C, and EBNA-LP and membrane proteins LMP1, LMP2A, and LMP2B.79 In type-I latency, most frequently seen in BL, only EBNA1 and LMP2A are observed, and in type-II latency (nasopharyngeal cancers), EBNA1, LMP1, and LMP2 are expressed.79 It is commonly believed that the different types of latency observed can be attributed to different viral/host interactions and are explained by the utilization of different promoters (Table 24.1).
Burkitt's Lymphoma
BL accounts for 30% of all EBV-related lymphomas. It was the first characterized and most frequently continues to be observed as lymphoid malignancies located on the jaw and neck.82 Almost all cases of endemic BL are infected with EBV, but only 20% of sporadic BL cases are positive for EBV.79 Both types of BL feature a chromosomal translocation of either t(8,14), t(2,8), or t(8,22), which causes the c-myc oncogene to relocate near the immunoglobin gene of chromosome 2, 14, or 22.83 The effect of this translocation is to abrogate the biological functions of c-myc, which is implicated in the regulation of cell proliferation, differentiation, and apoptosis.
Lymphoproliferative Disorders
The family of lymphoproliferative disorders (LPDs) begins with polyclonal B cell hyperplasia, which may progress to monoclonal lymphoma.84 The LPDs are observed most frequently in primary immunodeficient patients83 but more frequently are being observed in posttransplant patients as immunosuppressant drugs become more effective.85 A risk factor for LPDs may be the development of EBV viremia, which has led to recommendations to monitor EBV viremia and reduce immunosuppressant
Table 24.1
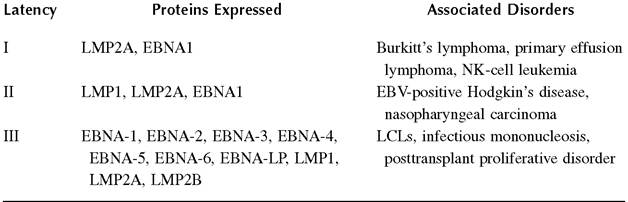
treatments if plasma viral copy numbers begin increasing rapidly.86 Primary effusion lymphoma (PEL) is a type of LPD that occurs in AIDS patients and is characterized by liquid growth in the body cavity, without a solid-mass tumor.87 It is almost always seen in patients with an EBV infection of type-I latency who are usually coinfected with Karposi’s sarcoma-associated herpesvirus. EBV may also be associated with T cell lymphomas88 and NK-cell leukemia.
EBV Proteins That Modulate Apoptosis
LMP1
Latent membrane protein 1 (LMPl) has a dramatic influence upon infected cells, because it radically affects the molecular signaling that occurs through its interactions with TRAFs, TRADD, JAK3, activation markers, and antiapoptotic molecules and has a cumulative effect of reducing the susceptibility of a cell to apoptosis. LMP1 is an integral membrane protein of 386 amino acids that is encoded by the BNLF1 gene. LMP1 is composed of three regions, each with differing functions. The short amino terminus (17 amino acids) is responsible for tethering the protein to the membrane in the proper orientation, and it is also the site of ubiquination. Six hydrophobic transmembrane domains (200 amino acids total) promote oligomerization centered around the protein rafts in the membrane.89 The carboxy terminus varies in length depending on the number of repeats of an 11 amino acid segment but averages approximately 200 amino acids.90 Within the carboxy terminus are three C-terminal activation regions (CTAR) regions that are associated with the protein’s antiapoptotic effects (detailed below). LMP1 has a typical half-life of 2 to 5 h within the cell, dependent on cell type, after which it is cleaved at Leu 242 and releases the carboxy cytosolic tail from the transmembrane region. Additionally, LMP1 is subject to phosphorylation within the C- terminal domain at Ser 313 and Thr 324.91
Wang et al. first identified LMP1 as possessing oncogenic (and antiapoptotic) properties when they transfected it into NIH3T3 and Rat-1 cell lines and observed a resulting increase in contact inhibition and anchorage independent growth.92 Furthermore, after LMP1-expressing cells were introduced into nude mice, it induced the growth of lymphoid tumors.92 LMP1 has four functions that favor its oncogenic properties: transfection with LMP1 can induce DNA synthesis, similar to the DNA synthesis seen during B cell activation; LMP1 has the ability to upregulate the production of antiapoptotic molecules; LMP1 interferes with the cell cycle of progression; and evidence of LMP1 control of matrix metalloproteins and IL-8 suggests that it may play a role in angiogenesis and metastasis.89
In addition to its transforming and toxic properties, LMP1 disrupts the signal transduction pathway of B and T cells. LMP1 expression induces an upregulation in the cell adhesion molecules ICAM-1, LFA1, and LFA3. It also increases activity of the CD71 transferrin receptor; activation markers CD21, CD23, and CD40; and antiapoptotic molecules Bcl-2 and A20.93-95 However, this upregulation is seen only in the latent infections of EBV. The lytic form of LMP1, which lacks the 128 amino acids on the amino terminus, does not display the transactivating or antiapoptotic activity94 and it does not impact DNA synthesis or the cell size:volume ratio.96
Through the presence of two NF-κB response elements located on the A20 promoter, LMP1 is able to enhance expression and activity of A20 (a zinc finger protein that confers resistance to the TNF). Transient transfection of Jurkat T cells with LMP1 increases CAT activity of an A20 promoter CAT construct, whereas mutating the B-binding sites of the A20 promoter negates any increase in CAT activity, indicating that the activation of A20 is mediated by NF-κB.97 Similarly, the ability of LMP1 to transactivate HIV replication is eliminated when the HIV LTR NF-κB binding site is deleted.98 It has been suggested that LMP1 influences NF-κB-dependent activation by phosphorylating and degrading the IB inhibitor of NF-κB, which then allows the NF-κB to translocate to the nucleus.99 Additionally, the ability of LMP1 to activate NF-κB may account for the property of LMP-1 to upregulate ICAM1, a cellular adhesion molecule.
LMP1 is similar in function, although not in amino acid homology, to members of the TNF receptors, especially TNFR1 and CD40,100 because it is able to bind to many of the same proteins as do TNFRs. These interactions occur primarily through the CTARs located on the carboxy terminus of LMP1.
CTAR1
LMP1 is able to interact with the TNF receptor-associated factor (TRAF) proteins. These six proteins (TRAF1-6) are involved with signal transduction and interact with the cytoplasmic tail of TNF receptors. LMP1 was shown to interact with TRAF1, TRAF2, and TRAF3, and the first 44 amino acids of the carboxy terminus are mandatory for that binding to occur. The interaction of TRAF1/TRAF2 heterodimers with the LMP1 TRAF domain results in cellular activation, ultimately resulting in B phosphorylation and nuclear translocation of NF-B.
Stimulation of the TRAFs initiates a signal cascade that results in the activation of not only the NF-B pathway but also the mitogen-activated protein kinase (MAPK) pathway, the c-Jun N- terminal kinase (JNK) pathway, and the PI3K pathway, all of which repress antiapoptotic signaling within the cell.100 The decreased rates of apoptosis promote an environment in which tumor development may be favorable. For example, TRAF2 binds to CTAR1 and becomes activated, after which it, in turn, activates NIK. NIK activation leads to the activation of the NF-B pathway, which initiates transcription for a number of antiapoptotic molecules, including A20, Bcl-2, Bfl-1, and cIAP2.100 A similar schema and result occur for the p38∕MAPK pathway,101 and although both pathways were proven to operate independently of each other, both are initiated through TRAF2.
CTAR2
The NF-B and MAPK pathways are capable of being activated through not only CTAR1 but also CTAR2. For that to occur, TRADD interacts via its N-terminus with the C-terminus of CTAR2.100 The TRADD activating site has proven crucial for the LMP1 transformation of B lymphocytes, because LMP mutants lacking the last three amino acids are incapable of transforming B cells.102 The CTAR2 recruitment of TRADD is also responsible for the activation of the JNK pathway.103 Stimulation of the JNK pathway upregulates phosphorylation and transcriptional activity of c-Jun (a mitogenic transcription factor). This leads to the binding of TRAF2 and TRADD to CTAR2 and the formation of AP-1 complexes, which increase transcription associated with certain promoters.103 However, the requirement for CTAR2 has proven to be cell-type specific and could happen through CTAR1 if TRAF1 were present.101 TRADD can induce apoptosis by means of TRAF2-mediated NF-B activation or promote apoptosis through association with Fas-associated death domain (FADD) protein, which initiates caspase-8 clustering activation and ultimately results in apopto- sis.104 Because LMP1 lacks TNFR death domains yet interacts with TRADD, the TNFR-mediated antiapoptotic signals of NF-B are favored after LMP1 activation of TRADD.
Additionally, there is some evidence that LMP1 can act as a tumor suppressor as well. Floett- mann et al. transfected LMP1 into an EBV-negative cell and observed an inhibition of the cell cycle progression.105 More telling, EBV-negative BL transfected into LMP-1 decreases the tumor- ogenicity and clonality from control cells.106 In total, this suggests that LMP1 plays a complicated role within the progression of an EBV-infected cell to a cancerous state; however, the overall effect seems to be antiapoptotic.
EBNA1
EBNA1 is a DNA-binding protein encoded by the BKRF1 gene of EBV. It is 641 amino acids long and is the only EBV latent protein to be expressed in all three forms of viral latency.107 EBNA1 functions in the virus by dimerizing and binding DNA, where it acts as a transcriptional enhancer to assist in maintaining and replicating the viral genome,108 and EBNA1 is restrained to the nucleus by c-myc, the nuclear matrix attachment factor. When the c-myc gene is silenced or translocated, as often occurs in BL, the EBNA1 invades the cytoplasm and is not able to maintain the EBV genome.109 Within the infected B cell, EBNA1 prevents cytotoxic T cell responses through a glycine-alanine rich region that interferes with the major histocompatibility complex (MHC), thus inhibiting the EBNA1 peptide antigen processing that would normally occur within an infected cell. Additionally, EBNA1 has been observed to have oncogenic properties, as 20% of transgenic mice that expressed EBNA1 developed B cell lymphoma.110 Moreover, EBNA1 is expressed in more than 95% of endemic BL cases, but in sporadic BL, the instance of EBNA1 is less than 20%.
EBNA2
Similar to EBNA1, EBNA2 is a transcription factor that is coded for by the BYRF1 gene. Interestingly, it is the first protein to be expressed after infection of B lymphocytes111 and is critical to the immortalization of B cells.112 It exists in two different forms (EBNA2A and EBNA2B), and both forms are required to immortalize cells.113 Rickinson et al. showed, by comparisons of complementary deficient strains, that EBNA2B is much less efficient at transforming B cells than EBNA2A.114 The genetic variation between the two strains occurs in the middle of the DNA and most significantly features a 42-base pair deletion in EBNA2B.113 Within the viral genome, EBNA2 is a transactivator of LMP1 and LMP2A, the Cp promoter (that regulates latent EBNA1) and HIV- LTR. In addition, EBNA2 is responsible for some of the reprogramming of cell gene expression by activating a variety of promoters.115 Within the cellular genome, it transactivates CD21 and CD23 along with c-fgr, another proto-oncogene.111
EBNA3, EBNA4, EBNA5, EBNA6
EBNA3, EBNA4, and EBNA6 (also called EBNA3A, EBNA3B, and EBNA3C) have been shown to act to arrest the infected B lymphocyte’s growth and division cycle by disrupting the G2/M checkpoint.116 After a cell becomes infected, EBNA2 and EBNA5 are expressed within the first 32 h, which corresponds with the progression from G0 to G1. As the cell moves into S phase, an increase in the levels of EBNA3, EBNA4, and EBNA6 can be observed.116 EBV modifies the activities of the tumor suppressor gene pRb by modulating the signaling pathways that affect it and thus obliterating its checkpoint function.117 EBNA5, in combination with EBNA2, has been shown to upregulate the activity of cyclin D2 and stimulate the resting B cell into G1 phase. The G1 checkpoint is then circumvented by EBNA6 as it disrupts the cyclin∕CDK-pRB-E2F signaling pathway,118 and EBNA6 is also thought to disrupt the G2/M progression.
EBNA-LP
When a B lymphoctye becomes infected with EBV, EBNA2 and EBNA-leader protein (LP) are the first two viral proteins to be expressed.119 EBNA-LP is a phosphoprotein that is the first protein encoded on the multicistronic mRNA that codes for all the EBNAs. It is located in both the cytoplasm and nucleus (where it associates with the nuclear matrix) and has been shown to stimulate EBNA2-mediated transcriptional activation of genes such as LMP1.120 It is not essential for viral survival, but it has been shown to bind to a number of molecules involved in viral life cycle, cellular mechanics, antiapoptotic molecules Bcl-2, p53, and pRb, and HS1-associated protein HAX-1.119 HAX-1 has been implicated in the regulation of apoptosis. The antiapoptotic characteristic of HAX-1 has been attributed to the Bcl-2 homologous domains BH1 and BH2 that are known to be the apoptosis regulatory sites in Bcl-2.121 Consequently, EBNA-LP modifies the net cellular susceptibility toward apoptosis.120,121 These results imply that as soon as a cell becomes infected with EBV, the viral proteins are enhancing antiapoptotic and protransformation signals.
EBER
EBERs (Epstein-Barr encoded RNAs) are viral protein transcripts that are found in almost all infected cells. They are usually approximately 250 nt nonpolydenylated RNA polymerase III transcripts and are usually found in the nucleus.122 EBERs are thought to inhibit the activity of double-stranded RNA-activated protein kinase and thus provide resistance to the host antiviral defense of interferon-α induced apoptosis. However, they are not required for viral maintenance or replication, and they are not required for the immortalization of B cells. There is also some speculation that EBERs enhance tumorigenicity.122
Diagnosis and Therapy
Diagnosis of EBV-associated malignancies requires tissue diagnosis, often with molecular techniques, to confirm the presence of EBV-encoded transcripts.123 After being diagnosed, EBV-asso- ciated malignancies are often amenable to standard anti-neuroplastic therapies of chemotherapy and radiation. However, in such cases, reversal of underlying immunosuppression is an important adjunct therapy. When applied to the therapy of HIV-associated EBV malignancies, immune reformation associated with HAART can independently reduce EBV viral replication124 as well as improve survival from such malignancies.125
More on the topic CRYPTOSPORIDIUM:
- Cryptosporidium cuniculus Infection: Cryptosporidiosis
- Diarrhea Cryptosporidium
- Cryptosporidium spp. Infection: Cryptosporidiosis
- Cryptosporidium spp. Infections
- Cryptosporidium spp. Infection: Cryptosporidiosis
- PARASITIC DISEASES
- BIBLIOGRAPHY FOR PARASITIC DISEASES
- Bibliography for parasitic diseases
- CONTENTS
- Protozoal Infections
- CONCLUSIONS
- PARASITIC DISEASES