Digestive System
Sabrina M. Timperman, DVM
OUTLINE
INTRODUCTION, 380
BASIC STRUCTURE OF THE GASTROINTESTINAL
TRACT, 381
REGULATION OF GASTROINTESTINAL FUNCTION, 381
ORAL CAVITY, PHARYNX, AND ESOPHAGUS, 382 TEETH, 383
Tooth Surfaces, 384
Tooth Structure, 384
Deciduous Teeth (Baby Teeth), 385
Heterodont Dentition, 385
Dental Formula, 386
TONGUE, 387
SALIVARY GLANDS, 387
TEMPOROMANDIBULAR JOINT, 389
PHARYNX, 390
ESOPHAGUS, 390
DIGESTION IN THE ORAL CAVITY AND PHARYNX, 390
SWALLOWING/DEGLUTITION, 391
ABDOMINAL CAVITY, 392
STOMACH, 392
Basic Structure and Overall Function, 392
MONOGASTRIC STOMACH AND DIGESTION, 393
Stimulation of Secretions, 394
Monogastric Stomach Motility, 394
Control of Gastric Motility, 395
Gastric Emptying, 395
Digestion in the Stomach, 396
RUMINANT STOMACH AND DIGESTION, 396
Forestomachs and Abomasum, 397
Motility of the Ruminant Stomach, 398
Reticulorumen Ecosystem, 400
Carbohydrate Digestion in Ruminants, 401
Lipid Digestion in Ruminants, 401
Protein Digestion in Ruminants, 401
Glucose Production in Ruminants, 402
SMALL INTESTINE AND ASSOCIATED
STRUCTURES, 402
Basic Structure and Function, 402
SECRETIONS OF THE SMALL INTESTINE, 404
PANCREAS, 404
LIVER, BILE DUCT, AND GALLBLADDER, 405
Bile Formation and Bilirubin Excretion, 406
Nutrient Processing in the Liver, 408
SMALL INTESTINAL MOTILITY, 410
Regulation of Small Intestinal Motility, 410
DIGESTION IN THE SMALL INTESTINE, 410
Carbohydrate Digestion, 410
Protein Digestion, 410
Absorption of Monosaccharides, Dipeptides, Tripeptides, and Amino Acids, 411
Lipid Digestion and Absorption, 412
THE LARGE INTESTINE, 412
Basic Structure and Function, 412
MOTILITY OF THE LARGE INTESTINE, 413
REGULATION OF THE LARGE INTESTINE
MOTILITY, 414
DIGESTION AND ABSORPTION IN THE LARGE INTESTINE, 414
EMPTYING OF THE RECTUM, 415
LEARNING OBJECTIVES
When you have completed this chapter you will be able to:
1.
List the functions of the digestive system.2. Describe the epithelial and muscle layers of the gastrointestinal tract.
3. Explain the process of peristalsis and segmentation.
4. List the structures of the oral cavity.
5. Name the types of tooth found in carnivores and herbivores.
6. Describe the structure of teeth.
7. Differentiate between mechanical and chemical digestion.
8. List the structures of the monogastric stomach and describe the function of each area.
9. Explain the effects of amylase, lipase, gastrin, pepsin, pepsinogen, prostaglandins, mucin, bicarbonate, secretin, cholecystokinin, proteases, and hydrogen and chloride ions on the gastrointestinal system.
10. Describe the structure and functions of the rumen, reticulum, omasum, and abomasums.
11. Differentiate between fermentative and nonfermentative digestion.
12. List the segments of the small intestine and describe the structure of the small intestinal mucosa.
VOCABULARY FUNDAMENTALS
Aborally ahb-ohr-ahl-e
Acetylcholine ah-set-ehl-ko-len
Amino acid ah-me-no ah-sihd
Amylolytic bacteria ahm-ah-lo-liht-ihck bahck-teer-e-ah Apical a-pihck-ahl
Aradicular hypsodont a-rah-dihck-yoo-lar hihp-suh-dohnt Bilirubin glucuronide bihl-e-ru-bihn gloo-kyuhr-uh-nιd Brachyodont brah-ke-o-dohnt
Brush border bruhsh bohr-dar
Buccal cavity buhck-ahl kahv-ih-te
Canine ka-nιn
Cardia kahr-de-ah
Carnassial teeth karh-nas-e-ahl teth
Carnivore kahr-nah-vohr
Cecum sek-uhm
Cellulolytic sehl-u-lo-liht-ihck
Chemical kehm-ih-kuhl
Chief cell chef sehl
Cholecystokinin ko-leh-sihs-tuh-kι-nihn Chyme kιm
Chymotrypsinogen kι-mo-trihp-sihn-o-jen Colon ko-luhn
Coronal kuh-ro-nahl
Crown kroun
Deciduous teeth (baby or milk teeth) de-sihd-u-uhs teth (ba-be or mihlk teth)
Deglutition (swallowing) de-gloo-tihsh-uhn (swahl-lo-ihng)
Dental formala dehn-tahl fohr-myoo-lah
Dentin dehn-tihn
Duodenum doo-o-den-uhm
Emesis ehm-eh-sihs
Enamel e-nahm-ahl
Endocrine ehn-do-krihn
Endopeptidase ehn-do-pehp-teh-daz
Enteric ehn-tear-ihck
Enturic nervous system ehn-tear-ihck nar-vuhs sihs-tehm Enterochromaffin-like cell (ECL-cell) ehn-teh-ro-kro- mah-fihn-lιk seh) (ECL-sehl)
Enterohepatic circulation ehn-teh-ro-heh-paht-ihck sar-kyoo-la-shuhn
Enteropeptidase ehn-teh-ro-pehp-teh-daz
Eructation e-ruhck-ta-shuhn
Eustachian tube (auditory tube) yoo-sta-osohbehn t (ahw-dih-tohr-eotob)
Exocrine ehcks-o-krihn
Exopeptidase ehcks-o-pehp-teh-daz
Fermentation far-mehn-ta-shuhn
Forestomach fohr-stuhm-uhck
Fundus fuhn-duhs
G cell jee sehl
13.
List the segments of the large intestine and describe the general functions of each segment.14. Describe carbohydrate, protein, and fat digestion.
Gastric gahs-trihck
Gastrin gahs-trihn
Gluconeogenesis gtoo-ko-ne-o-jehn-eh-sihs
Glycogenolysis glι-ko-jehn-ohl-eh-sihs
Hepatocyte heh-paht-o-sιt
Herbivore har-bah-vohr
Heterodont teeth heht-ar-o-dohnt teth
Hydrolysis hι-drohl-uh-sihs
Hypsodont hihp-s-o-dohnt
Ileum ihl-e-uhm
Incisor ihn-sι-zar
Jejunum jeh-joo-nuhm
Labial surface la-be-ahl suhr-fihs
Lactase lahck-taz
Maltase mahl-taz
Mastication mahst-eh-ka-shuhn
Mechanical digestion meh-kahn-ih-kahl dι-jehst-shuhn Mesentery mehs-ehn-tear-e
Microvilli mι-kro-vihl-lι
Molar mo-lar
Monogastric animal mohn-o-gahs-trihck ahn-uh-muhl
Monoglyceride mohn-o-glihs-ar-rιd
Monosaccharide mohn-o-sahck-ah-rιd
Mucosa myoo-ko-sah
kMucus nec cell myoo-hkcukhs ne sehl
Myenteric plexus (Auberbach’s plexus) mι-ehn-tear-ihck plehck-suhs (awb-ar-bohckz plehck-suhs)
Neck nehck
Nonprotein nitrogen compound (NPN compound) nohn—pro-ten nι-truh-jehn kohm-pohwnd
(NPN kohm-pohwnd)
Omnivore ohm-nah-vohr
Palate pahl-iht
Parietal cell pah-rι-eh-tahl sehl
Pepsin pehp-sihn
Peptidase pehp-teh-daz
Peristalsis pear-ih-stahl-sihs
Plication plι-ka-shuhn
Polysaccharide pohl-e-sahck-uh-rιd
Prehension pre-hehn-shuhn
Premolar pre-mo-lar
Procarboxypeptidase pro-kahrb-ohck-se-pehp-tιd-az
Proelastase pro-e-lahs-taz
Proenzyme (zymogen) pro-ehn-zιm (zι-mo-jen)
Proteolytic enzyme pro-te-o-liht-ihck ehn-zιm
Pyloric antrum pι-lohr-ihck ahn-truhm
Pyloric sphincter pι-lohr-ihck sfihnk-tar
Radicular hypsodont rah-dihck-yuh-lar hihp-o-dohnt
Reticulorumen reh-tihck-u-lo-r u -mehn
Reticulum reh-tihck-u-luhm
Root root
Rumen ru-mehn
Ruminant ru-mehn-ahnt
Rumination ru-meh-na-shuhn
Saliva sah-lι-vah
Salivary gland sahl-eh-vear-e glahnd
Secretin seh-kret-ihn
Segmentation sehg-mehn-ta-shuhn
Submucosa suhb-myoo-ko-sah
Submucosal plexus (Meissner’s plexus) suhb-myoo-ko- sahl plehck-suhs (mιz-narz plehck-suhs)
Sucrase soo-kras
Temporomandibular joint tehm-pohr-o-mahn-dihb-u- lahr joynt
Triadan system trι-ah-dehn sihs-tehm
Trypsin trihp-sehn
Urobilinogen yar-o-bih-lihn-o-jehn
Villi vihl-lι
Volatile fatty acid (short chain fatty acid) vohl-uh-tihl faht-e ah-sihd (shohrt chan faht-e ah-sihd)
INTRODUCTION
All animals need to provide energy and nutrients to their body's cells and tissues to perform daily activities, ensure the cells' ability to do work, and promote growth and normal development.
In the animal kingdom, often these needs are met by breaking down and absorbing nutrients that the animal has consumed. The process of extracting nutrients and creating energy from food begins when an animal consumes a meal.What an animal eats, however, varies among species. Herbivores eat plants. In herbivores, such as horses and cattle, the process of converting consumed plant material into usable nutrients and energy is heavily dependent on microbial fermentation chambers within the animals' gastrointestinal (GI) tract. Carnivores eat meat. In carnivores, such as the cat, the gastrointestinal tract itself is responsible for converting consumed meals into nutrients and energy without the aid of a microbial fermentation chamber. Dogs have similar dental anatomy to cats. Both have sharp, pointed canine teeth for tearing flesh and sharp premolars and molars, which can be used for cutting meat, but dogs are no longer strictly carnivores. They are now omnivores because they have evolved to utilize both plant material and meat. Omnivores, such as humans and pigs, eat a combination of plant materials and meats.
Although domestic animals share many similarities, there are distinct anatomic differences depending on what they consume. Ruminants such as cattle, sheep, and goats are herbivores. They have large microbial fermentation chambers where the plant materials are partially broken down before the food reaches the true stomach. Nonruminant herbivores, such as horses, are called hindgut fermenters. They have an extremely well developed and expansive fermentation chamber (the cecum) at the junction of the small and large intestines that allows microbes to help break down plant materials (Figure 16-1). In contrast, carnivores have an inconspicuous and small cecum because microbes play an insignificant role in breaking down their food.
Digestion is a process that begins inside the gastrointestinal tract but, interestingly, anything within the gastrointestinal tract can be considered still outside the body.
During embryonic development, the animal starts off as a flat sheet of cells that folds in and forms a tube. The lumen of the tube is what will become the lumen of the gastrointestinal tract.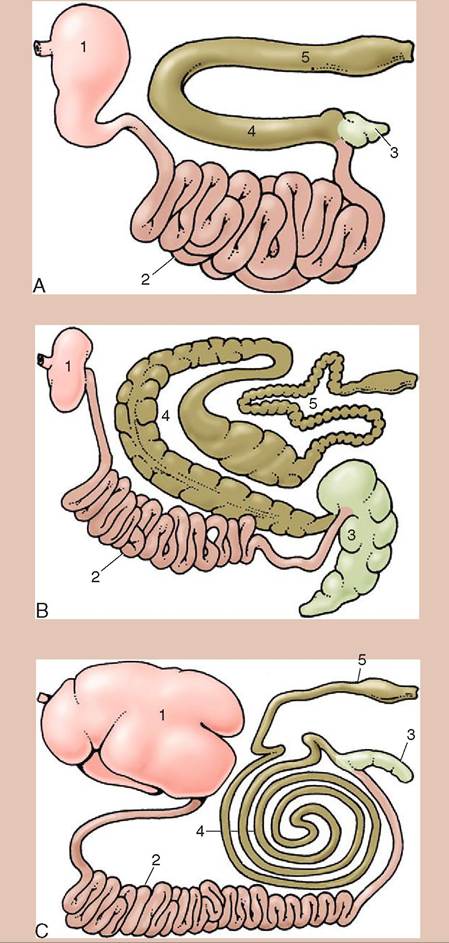
FIGURE 16-1 Gastrointestinal tracts A, of the dog, B, of the horse, and C, of cattle laid out in one plane. 1, Stomach; 2, small intestine; 3, cecum; 4, ascending colon; 5, descending colon. (From Dyce KM, Sack WO, Wenseng CJG: Textbook of veterinary anatomy, ed 4, St Louis, 2010, Saunders.)
Initially the tube is closed at both ends but soon the oral and anal openings will develop so that the gastrointestinal tract essentially becomes a continuous tube with openings at either end. The breakdown of nutrient macromolecules into their more basic parts occurs within the lumen of this tube. Even after they have been broken down into basic parts, the smaller nutrients do not enter into the body until they are absorbed across the intestinal tract wall.
Digestion refers to the part of the process in which larger molecules are broken down into their smaller component parts. This breakdown process occurs in two very different ways, mechanical digestion and chemical digestion. Mechanical digestion refers to the gastrointestinal tract movements, which physically break food up into its smaller parts. In chemical digestion, a chemical reaction breaks the bonds holding macromolecules together, resulting in the production of smaller molecules. When they are small enough, molecules are able to be absorbed across the intestinal membrane and enter the body.
BASIC STRUCTURE OF THE GASTROINTESTINAL TRACT
The gastrointestinal (GI) tract runs from the oral cavity to the anus and includes structures such as the oral cavity, esophagus, stomach, small intestine, and large intestine. When referring to the stomach, the term gastric is used and when referring to the intestines, the term enteric is used.
Most of the GI tract wall consists of four layers of tissue, from the lumen outward the mucosa, the submucosa, the muscular layers (circular and longitudinal), and the serosa (Figure 16-2). Within these layers there are some variations, depending on which part of the GI tract you are looking at.
The mucosa or innermost layer consists of its own three parts, the epithelium, the lamina propria, and the muscularis mucosae. The epithelium, which lines the lumen, is made up of different types of epithelial tissue based on its location. The epithelial tissues near the mouth and anus consist of layers of stratified squamous epithelium. The stratified cells make up many layers, which enable them to provide protection. The rest of the epithelium of the gastrointestinal tract consists of simple columnar epithelium. These epithelial cells are connected to one another with tight junctions, helping to protect the animal by creating a barrier that prevents harmful or unwanted substances from entering the body.
The second layer of the mucosa, the lamina propria, is made up of loose areolar connective tissue. This layer contains blood and lymph vessel, and glands. The third layer of the mucosa is the muscularis mucosae, a thin layer of smooth muscle, which helps form the mucosa into folds that help to increase the surface area of the lining of the stomach and intestines. The increased surface area provides a larger area for absorption of nutrients into the body.
The submucosa, which is not part of the mucosa but lies underneath it, consists of dense connective tissue.
The muscular layer is the third layer and in most parts of GI tract consists of two smooth muscle layers, an inner circular and an outer longitudinal one. In the oral cavity, pharynx, and in some species the esophagus, skeletal muscle is also present. There is also an external anal sphincter that is made of skeletal muscle, which assists animals in controlling the timing of their defecation.
The last layer making up the gastrointestinal tract wall is the serosa or adventitia, depending on whether that part of the GI tract is suspended from the body cavity (serosa) or surrounded by other tissue (adventitia). This layer is made up of loose connective tissue.
REGULATION OF GASTROINTESTINAL FUNCTION
The gastrointestinal tract is regulated by two different control systems. The first system involves a combination of the central nervous system and the endocrine system. The second system is unique to the gastrointestinal tract and consists of an enteric or intrinsic nervous system with an intrinsic endocrine/paracrine component. The enteric nervous system is commonly known as the “brain of the gut” and consists of different receptors, sensory neurons, interneurons, and motor neurons. The enteric nervous system controls both motor and secretory functions of the gastrointestinal tract and contains its own “pacemaker” cells. It is influenced by the autonomic nervous system, which can alter its degree of activity. The parasympathetic branch of the autonomic nervous system usually enhances digestive processes, whereas the sympathetic branch usually inhibits digestion. Afferent neurons travel to the central nervous system (CNS) from different types of receptor in the gut that monitor changes in GI tract tension as well as monitoring the chemical conditions of the GI tract. These neurons are associated with the autonomic nervous system and provide this sensory information to the CNS.
Two plexuses make up the enteric nervous system, the submucosal plexus (Meissner’s plexus) and the myenteric plexus (Auerbach’s plexus). The nerve fibers of both plexuses run the length of the GI tract. The submucosal plexus is located in the submucosa and controls secretions and blood flow in the GI tract. The myenteric plexus runs between the circular and longitudinal layers of smooth muscle, and is important in controlling movements of the GI tract through local reflexes.
The GI tract also contains intrinsic endocrine and paracrine systems, which have a regulatory function rather than a digestive one. For example, the endocrine hormone cholecystokinin inhibits gastric emptying and the endocrine hormone gastrin stimulates stomach motility. Endocrine
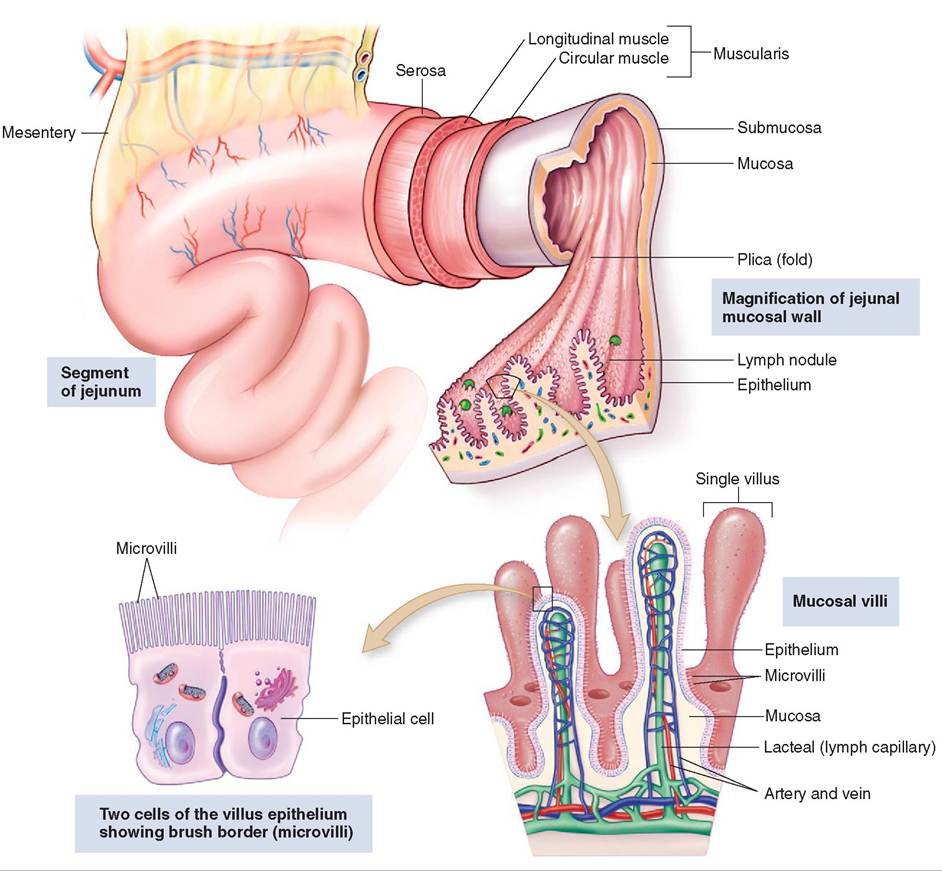
FIGURE 16-2 Small intestine. Note that the folds of mucosa are covered with villi and that each villus is covered with epithelium, which increases the surface area for absorption of food. (From Thibodeau G, Patton KT: Structure and function of the body, ed 14, St Louis, 2012 Mosby.)
cells secrete hormones directly into the bloodstream and have their effect at a distant site. Paracrine cells secrete substances into the interstitial fluid, which then travel by diffusion and affect nearby cells.
_______________________
TEST YOURSELF 16-1
1. What is the primary diet of a carnivore, an omnivore, and an herbivore?
2. What are two species of animal that require microbial fermentation to digest their food?
3. What is the purpose of the stratified squamous epithelium that lines much of the GI tract?
4. How many layers of muscle make up the muscular layer of the wall of the intestines?
5. What are the two nerve plexuses that make up the intrinsic enteric nervous system?
ORAL CAVITY, PHARYNX,
AND ESOPHAGUS
The entrance to the gastrointestinal tract is the mouth or oral cavity, also known as the buccal cavity. It contains the teeth, tongue, and everything else required to ingest food (Figure 16-3). The oral cavity consist of two parts: (1) the vestibule is the space between the outer surface of the teeth and the surrounding lips and cheeks and (2) the oral cavity proper is the space bordered by the inner surface of the teeth laterally and rostrally and by the hard and soft palate dorsally.
The oral fissure is the opening into the oral cavity. The lips mark the boundary of the oral fissure. There are both long, tactile hairs and regular hairs at the margins of the lips. Depending on the species, the lips can be very maneuverable
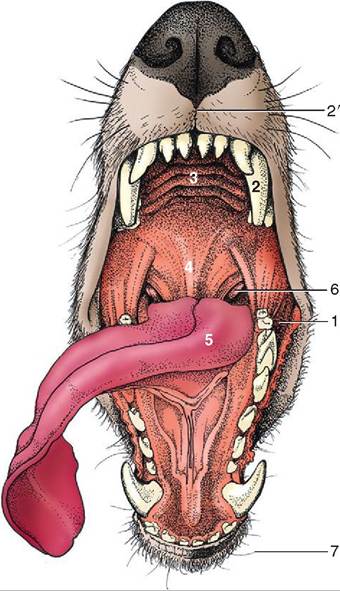
FIGURE 16-3 General view of the oral cavity of the dog. 1, Vestibule; 2, canine tooth; 2', philtrum; 3, hard palate; 4, soft palate; 5, tongue; 6, palatine tonsil; 7, tactile hairs. (From Dyce KM, Sack WO, Wensing CJG: Textbook of veterinary anatomy, ed 4, St Louis, 2010, Saunders.)
and be used to assist the animal in prehension, which is the process of bringing food into the oral cavity. The cheeks eorm the lateral walls of the vestibule. The lips and cheeks adree ma up of an outer layer of haired skin, a middle layer uf muscles and fibroelastic tissue, and an inner layer of mucosa that lines the vestibule and oral cavity. The middle muscular layer consists of the muscles of mastication (chewing), which contribute to the biting strength of the mouth. ⅞e philtrum is the cleft that divides the two halves pf the upper lip. In some species the philtrum is deeper and rmoomrienepnt, such as in carnivores, whereas other
avenimals ha a shallow inconspicuous philtrum that is hard to see, such as in horses.
The palate,chtiwsch a as the dorsal border of the oral cavity (roof of the mouth), consists of two distinct parts, the hard palate and the soft palate. The more rostral part is the hard palate, which is made up of the palatine, maxillary, and incisive bones that are covered by a mucous membrane. Several elevations cross the hard palate trans- eveatrisnelgy, cr ridges. The soft palate is the caudal part of the palate. It is made up of muscle and connective tissue and divides the pharynx (throat) into an oropharynx, twheer lo area that connects with the mouth, and the nasopharynx, the upper area that leads into the nasal pas- shaegeway. T soft palate is raised to close off the nasal
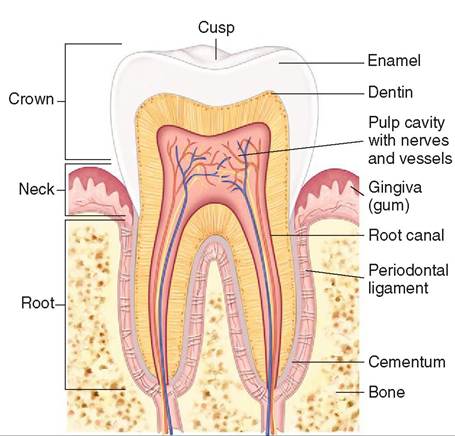
FIGURE 16-4 Longitudinal section through a tooth. A molar is sectioned to show its bony socket and details of its three main parts: crown, neck, and root. Enamel (over the crown) and cementum (over the neck and root) surround the dentin layer. The pulp contains nerves and blood vessels. (From Thibodeau G, Patton K: Structure and function of the body, ed 14, St Louis, 2012, Mosby.)
rpeavsesnagtien, gp food from entering the nasal passage-
uwraiyn,gd swallowing.
TEETH
Within the oral cavity you will find the teeth and tongue. The teeth are embedded in the upper maxilla bone and lower omnaen. dible b They are found in sockets or cavities called alveoli and are held in place by the periodontal ligament. eTeeth ar important in assisting the animal with the mechanical breakdown of food. Food is broken into smaller pieces ebayrtihneg,t cutting, and crushing action of the teeth that uccurs during the process of mastication (chewing).
The crown is the part of the tooth that projects above the gingiva (gums). ⅞e root is embedded in the alveoli below
CLINICAL APPLICATION
Tooth Resorption
deline odontoclastic resorptive lesions were first discovered in the necks of teeth, which explains why these lesions were initially known as “neck lesions.” Other species can also acquire similar lesions, so the name has been changed from feline odontoclastic resorptive lesion to tooth resorption. In this condition, tooth resorption occurs to form erosions, ewhich ar then covered with calculus or gingival tissue. Some afterted animals will show signs of pain and discom- eort, resulting in changes in behavior or appetite, whereas others show few symptoms. The level of treatment ranges fnriotomrimngo with minimal treatment to multiple tooth
extractions.
the gingiva. The tip of the root of a tooth is called the apex (plural: apices) where the blood vessels and nerves enter the tooth. The area where the crown and the root meet is called the neck (Figure 16-4).
TOOTH SURFACES
When performing a dental cleaning it is important to know what the different surfaces of the teeth are called in order to document properly where a lesion has occurred on the tooth. The outer surfaces, facing toward the cheeks and lips, are buccal and labial respectively. The inner surfaces, facing the tongue and soft palate, are lingual and palatal (Figure 16-5). In the space between the teeth, the edge of the tooth facing the midline or center of the dental arch is the mesial surface. The edge facing away from the center of the dental arch is called the distal surface. Both the mesial and distal surfaces refer to the part of the tooth that is touching or nearly touching an adjacent tooth. The occlusal or masticatory surfaces are the surfaces on the upper and lower teeth that come together when the mouth is closed. When referring to something toward the crown of a tooth the term coronal is used, whereas something toward the root is apical.
TOOTH STRUCTURE
An understanding of the anatomy of the tooth is also crucial in any dental procedure. The crown is covered by a thin layer of white hard material called enamel. Enamel is the hardest substance in the body. Under the enamel is the dentin. Dentin forms the bulk of the tooth and is as hard as bone, but not nearly as hard as enamel. The dentin surrounds an inner area called the pulp cavity that contains the blood supply and nerves which supply the tooth (see Figure 16-4).
The type or classification of teeth an animal has varies depending on its species. Carnivores, humans, and pigs (except for their tusks) have teeth that are classified as brachyodont teeth. They have relatively small crowns and well developed roots. Ruminant incisors are also brachyodont teeth. These teeth do not continually grow because the apices of their roots are open for only a finite period of time.
A horse's incisors and cheek teeth, a boar's canine teeth (tusks), ruminant cheek teeth, and some of the teeth of rodents and lagomorphs are classified as hypsodont teeth (Figure 16-6). These teeth grow continuously during most of the life of the animal because of a large reserve of crown beneath the gingiva. Hypsodont teeth can be further divided into two different types of tooth, radicular hypsodont and aradicular hypsodont. All the cheek teeth of the horse are radicular hypsodont, meaning that the apices of their roots remain open for a significant part of the horse's life, leading to continued growth. They do eventually close and stop growing. The wear on the teeth is offset by their continued eruption until growth ceases. In horses, points and hooks can develop on the teeth owing to uneven wear on the occlusal or masticatory surfaces. These sharp elongated ridges must then be filed down through a process called floating the teeth to create a level occlusal surface.
Lagomorphs and some rodents have incisor and cheek teeth that are classified as aradicular hypsodont teeth. These
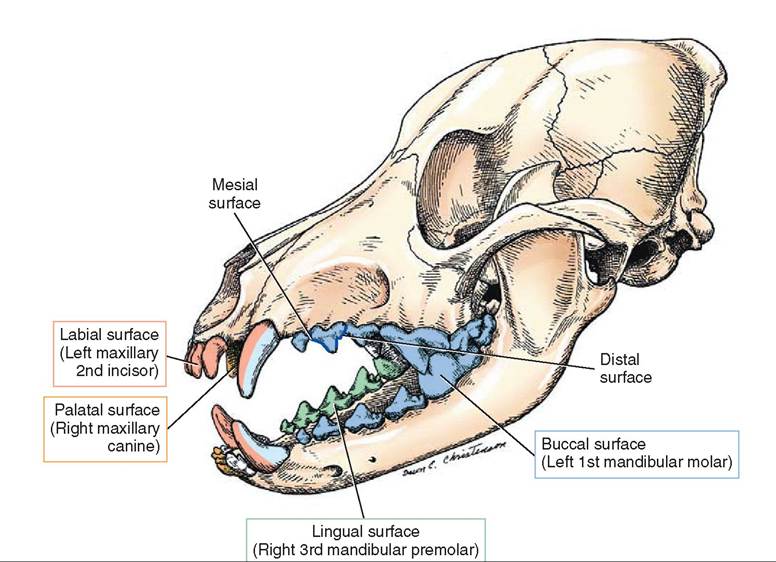
FIGURE 16-5 Tooth surfaces. (Redrawn from Christenson DE: Veterinary medical terminology, ed 2, St Louis, 2008, Saunders.)
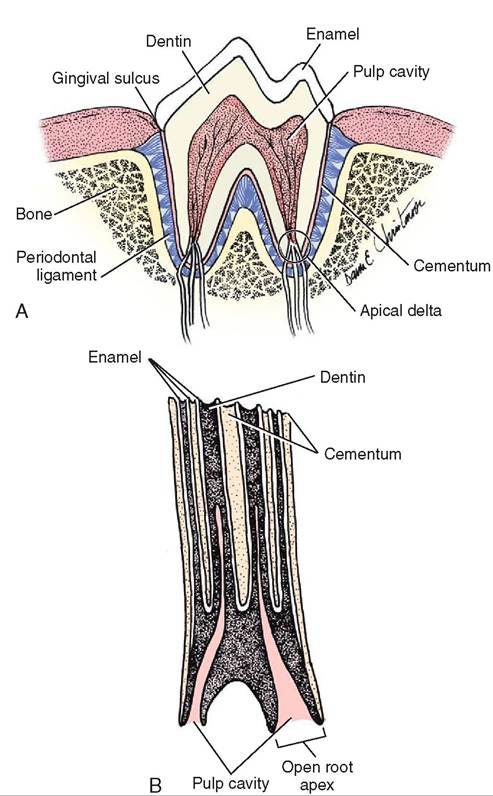
FIGURE 16-6 Comparative tooth anatomy. A, Carnivorous tooth (premolar; brachyodont tooth). B, Herbivorous tooth (premolar; hypsodont tooth). (From Christenson DE: Veterinary medical terminology, ed 2, St Louis, 2008, Saunders.)
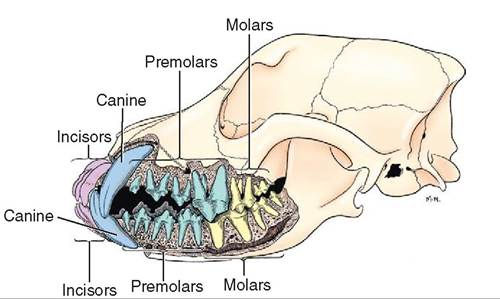
FIGURE 16-7 Jaws and teeth of an adult dog. Lateral view of the jaws, sculpted to show tooth roots. (From Evans H, de Lahunta A: Miller's anatomy of the dog, ed 4, St Louis, 2013, Saunders.)
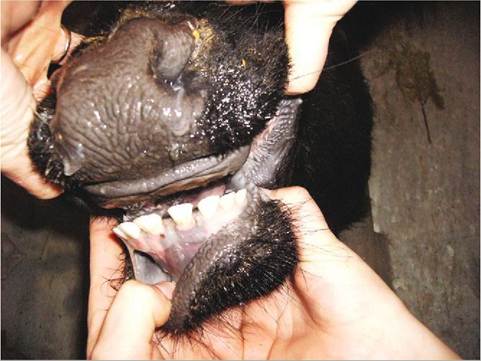
FIGURE 16-8 Dental pad and mandibular teeth of a cow. (From Holtgrew-Bohling K: Large animal clinical procedures for veterinary technicians, ed 2, St Louis, 2012, Mosby.)
teeth lack a true root and grow continuously throughout the life of the animal. The continued growth compensates for the wear on the teeth. A pet rabbit's or rodent's diet is different from what it would be in the wild. This prevents normal wear, but the teeth continue growing. These animals also may develop points or overgrowths on their teeth, which will require floating or odontoplasty (the process of recontouring a tooth surface).
The cementum is a thin bonelike covering over the roots of brachyodont teeth and most of the entire tooth superficial to the enamel in hyposodont teeth. The periodontal membrane is made of dense fibrous connective tissue that links the cementum with the alveolar wall, anchoring the tooth into the jaw.
DECIDUOUS TEETH (BABY TEETH)
All domestic species have two sets of teeth, deciduous dentition, also called the milk teeth or baby teeth, and permanent dentition, also called the adult teeth. The deciduous teeth tend to be smaller and whiter. They are present in the jaw at birth but erupt through the gums at different times in different species. For example, in the cat the deciduous teeth erupt starting at around 3 weeks and continue to erupt for about 6 weeks. The permanent set of teeth erupts at around 12 to 24 weeks (3 to 6 months) (Table 16-1). Some animals, especially horses, can have their age determined from their dentition. This skill requires a lot of practice to be accurate.
Heterodont dentition
Heterodont dentition refers to teeth of differing shapes and sizes. Domestic animals have heterodont dentition. There are four different types of teeth and each has a different function (Figure 16-7). Incisors are found in the premaxilla or incisive bone. They are small and are often used to cut and nibble on food. In some species, such as in ruminants, the upper front incisors are missing. Instead they have a thickened region called the dental pad that
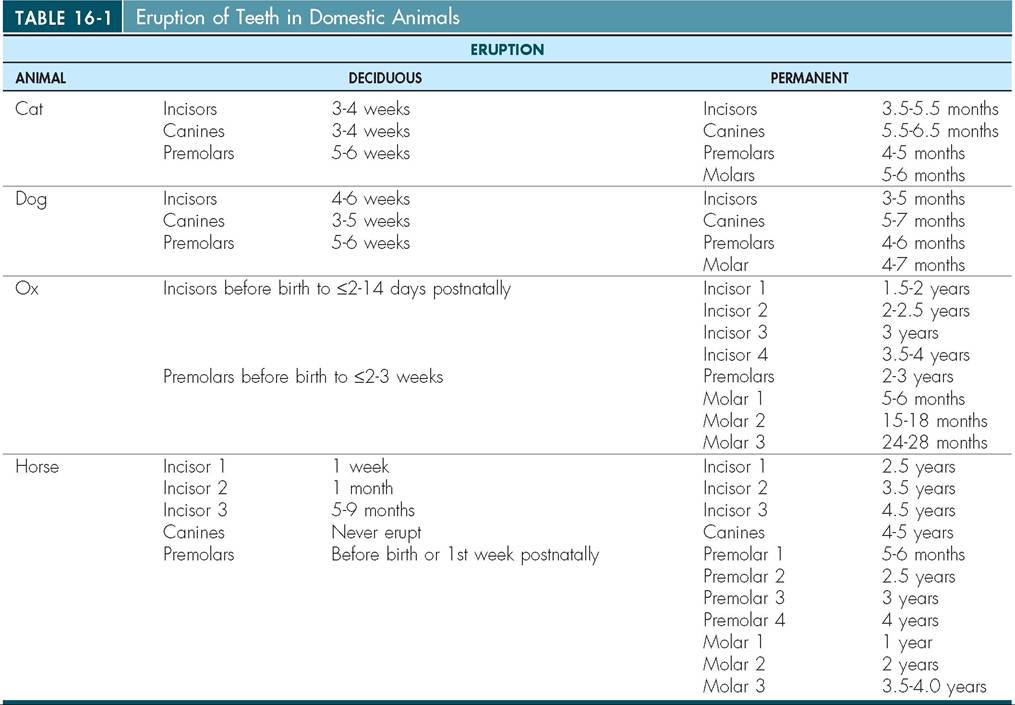
From Studdert V: Saunders comprehensive veterinary dictionary, ed 4, Philadelphia, 2012, Saunders, VitalBook file.
the lower incisors can grind and crush food upon (Figure 16-8). Other animals, such as the elephant, have tusks, which are modified first incisors. The bulk of the incisor is composed of very hard dentin, which is more commonly called ivory in the tusks of the elephant, walrus, and hippopotamus.
The canine teeth are located in the maxilla bone and the mandible. They are single teeth on each side of the jaw caudal to the incisors. These teeth are sharp and pointed and are used to tear flesh and hold prey. Ruminants do not have canine teeth. In horses, both mares and geldings may have canine teeth, but they are usually small if they are present at all. A stallion, on the other hand, can have well-developed canine teeth. In pigs, the canine teeth of a boar are exaggerated and open rooted, allowing continued growth. These teeth are also known as tusks.
The premolars and molars are called the cheek teeth and are also found in the maxilla bone and the mandible. The premolars act like shears, cutting and slicing meat from bones and grinding the food into smaller pieces. The premolars of the dog have sharp points. The horse's rudimentary upper first premolar is called the wolf tooth and is often missing or vestigial. Molars, which are only found in adult dentition, also assist in grinding and shearing. The largest cutting teeth in the jaw of the carnivore are the carnassial teeth, premolar 4 on the upper jaw and molar 1 on the mandible.
DENTAL FORMULA
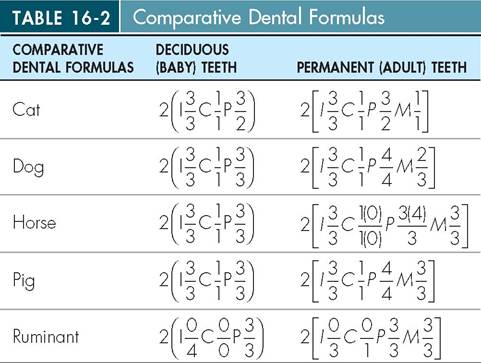
Although the types of tooth are similar, the number of each type of tooth differs with each species. The dental formula indicates how many of each type of tooth are present. The types of tooth are abbreviated as incisors (I), canines (C), premolars (P), and molars (M), and each abbreviation is followed by a number representing the number of the teeth of that type on one side of the upper and lower jaw. The dental formulas for several species are shown in (Table 16-2). Upper case letters represent the adult set of dentition; lower case letters denote the deciduous or baby set of teeth. The dental formula lists only half of the teeth, or one side of the oral cavity. In order to get the total number of teeth you would need to add all the numbers up from the maxilla and the mandible and then multiply by two.
Often it is necessary to document in a chart or make a record of a damaged or missing tooth. It is therefore crucial to also have a numeric system that can be used during charting to locate a specific tooth. The most commonly used
Adapted from Christenson DE: Veterinary medical terminology, ed 2, Philadelphia, 2008, WB Saunders Company; Noden DM, Delahunta AD: Embryology of domestic animals, Baltimore, 1985, Williams & Wilkins; Studdert V: Saunders comprehensive veterinary dictionary e-book, ed 4, St Louis, 2015, Saunders.
I, Incisor; C, canine; P, premolar; M, molar.
system is the Triadan System (Figure 16-9). Using this system, tooth numbering begins at the midline of the upper arch and is numbered as follows: right maxillary arch (100 series), left maxillary arch (200 series), left mandibular arch (300 series), and right mandibular arch (400 series). The first right maxillary incisor starting at the midline therefore is numbered 101 and the numbering continues caudally. The second incisor going away from the midline is 102, the third incisor is 103, and the canine is 104 and so forth. The deciduous teeth are numbered in the same fashion but starting in the right maxillary arch as the 500 series and moving in the same direction. Some teeth are missing in different species but the numbering continues in the same way and the number corresponding to the missing tooth is skipped. For example, right maxillary premolar 1 is missing in the cat so the numbering goes from right maxillary canine (104) to premolar 2 (106). Cats are also missing their first and second premolars on the lower mandible and the
TEST YOURSELF 16-2
1. What are the two parts of the roof of the mouth?
2. The part of the tooth that sticks above the gum line is
the___. It is covered with___.
3. What substance makes up the bulk of a tooth?
4. The inside surface of a tooth that faces the tongue is
the___ surface.
5. Where is the occlusal surface of a tooth?
6. What type of tooth will continue to grow throughout the life of an animal?
7. What are the four types of tooth that make up heter- odont dentition?
8. What numeric system is used to assign a specific number to each tooth in the mouth? numbering continues in the same way. Regardless of species, the numbering in the Triadan system is consistent so if you know that the left maxillary canine in the dog is 204, you know that the same tooth in a cat or a pig is also 204.
TONGUE
The tongue is also located in the oral cavity. It consist of both extrinsic muscles that anchor it in place and the intrinsic muscles that originate and insert on the tongue itself and make up the majority of the mass of the tongue. The fibers of the intrinsic muscles cross in multiple directions across the tongue making it flexible and maneuverable, which accounts for its fine and delicate movements. The ruminant tongue is especially flexible and is used to assist in prehension.
The tongue normally lies on the ventral surface of the oral cavity and consist of three parts, an apex, body, and root. The free unattached mobile tip of the tongue is called the apex; the body is the long and slender part that links the apex with the root. The root anchors the tongue to the hyoid bone and the sides of the mandible. The exterior of the tongue is covered by cornified stratified squamous epithelium.
Located on the dorsal surface of the tongue are different types of papilla. Some papillae have a mechanical function and assist in the grooming process and in moving the food bolus down into the pharynx. Specialized papillae contain taste buds, which allow the animal to experience a variety of different taste sensations.
The tongue not only senses taste but also has a nerve supply, which allows for sensations of pain, temperature, and touch. In addition to innervation the tongue is well supplied with blood. The superficial positioning of some of these blood vessels, especially in the dog, reflects an additional function of the tongue: thermoregulation through panting.
SALIVARY GLANDS
Salivary glands deposit saliva into the oral cavity via ducts. Saliva is extremely important in the process of digestion. It is composed mainly of water but also contains protein, electrolytes, antibodies (immunoglobulin [Ig]A), glycoproteins, other organic molecules, salivary bicarbonate and enzymes. Lysozyme, an enzyme found in the saliva, along with immunoglobulins, helps control the bacterial population in the oral cavity. Some species, mainly omnivores, as well as some avian species, have the starch-digesting enzyme amylase in their saliva. Amylase assists in the breakdown of starchy carbohydrates. Although it is secreted in the oral cavity, minimal digestion occurs there because of the short time food stays in the mouth. In pigs, rats, and humans, salivary amylase contributes significantly to the breakdown of starchy carbohydrates in the proximal stomach before the amylase is inactivated by hydrochloric acid in the stomach. Among the domestic animals, dogs, cats, and ruminants lack salivary amylase, horses produce a limited amount, whereas pigs produce the most.
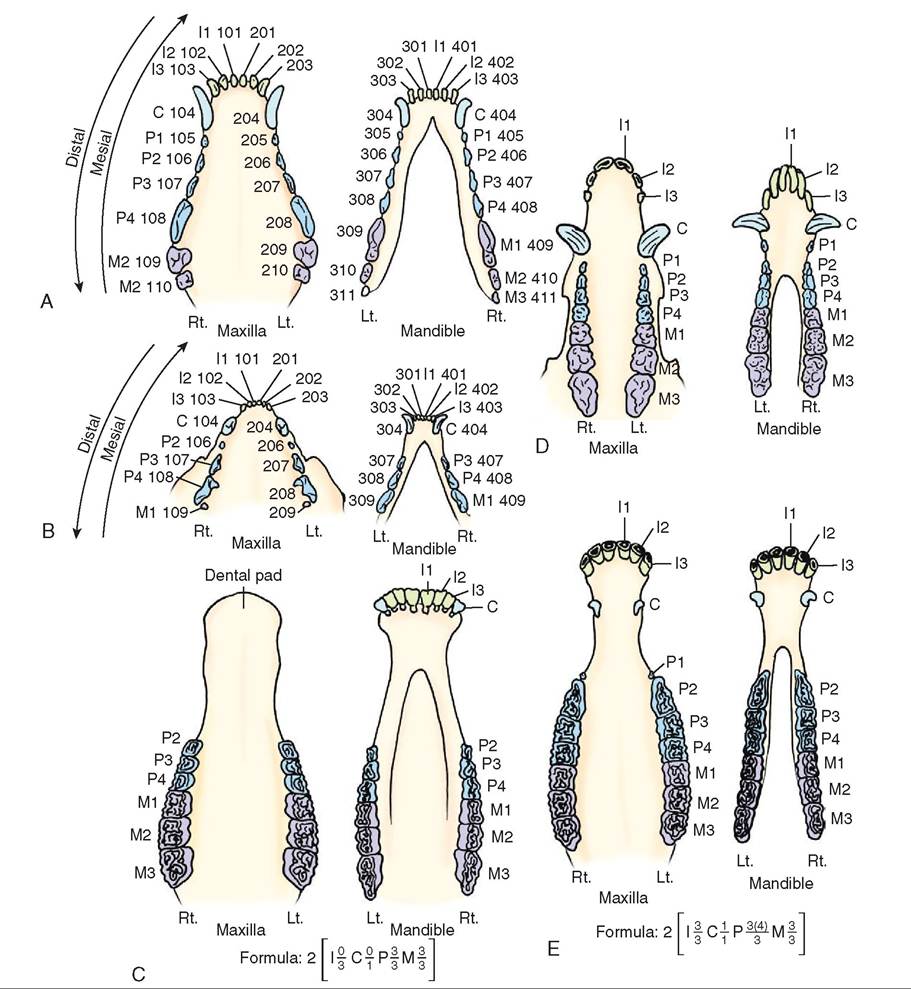
FIGURE 16-9 Comparative dental arcades. A, Canine. B, Feline. C, Bovine, ovine, and caprine. D, Porcine. E, Equine. (From Christenson DE: Veterinary medical terminology, ed 2, St Louis, 2008, Saunders.)
Saliva production varies depending on species and diet. Cattle can secrete up to 200 L of saliva per day. Ruminant saliva has a high pH (alkaline) and a high concentration of bicarbonate and other bases that are important for neutralizing the acids produced in the fermentation chambers of the forestomach.
Saliva has many functions including lubrication, antibacterial action, pH regulation, thermoregulation, and enzymatic digestion. The salivary glands are usually paired and located near the oral cavity. There are three main salivary glands, the parotid, mandibular, and sublingual glands, along with multiple smaller minor glands, such as the zygomatic (Figure 16-10). Depending on the gland, the secretion can be serous (watery), mucous (viscous), or mixed.
The parotid gland is located ventral to the ear and has a long duct extending to the oral cavity. Its secretion is serous in most species and accounts for approximately half of the total volume of saliva produced. The sublingual and the mandibular glands are located under the tongue and caudal to the angle of the jaw respectively, and both secret a mixed secretion in most species. The sublingual gland is divided into two parts in some species. In the dog there is a polystomatic part, which is more rostral and located on either side of the tongue, and a monostomatic part which is more caudal, but rostral to the mandibular salivary gland. The horse on the other hand has only the polystomatic part, whereas the ruminant has both parts, but they are in the opposite position to those of the dog, with the polystomatic part coming after the more rostral monostomatic part (Figure 16-11).
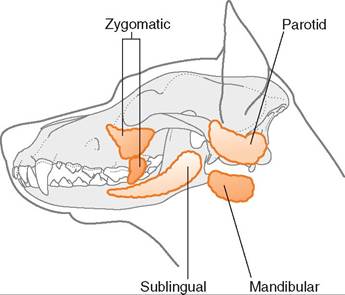
FIGURE 16-10 Salivary glands of the dog. (Adapted from Aspinall V, O'Reilly M: Introduction to veterinary anatomy and physiology, Oxford, 2004, Butterworth Heinemann. In Studdert V: Saunders comprehensive veterinary dictionary e-book, ed 4, St Louis, 2015, Saunders.)
TEMPOROMANDIBULAR JOINT
The temporomandibular joint (TMJ) is a condylar joint that forms the connection between the condylar process of the mandible (lower jaw) and the mandibular fossa of the temporal bone, which is part of the cranium. This connection occurs on both sides of the head, so there are two temporomandibular joints. The area where the two bones, the mandible and the temporal, articulate is enclosed in a joint capsule, and a thin fibrocartilagenous disc, or meniscus, divides the joint cavity into two compartments. The movements allowed by the TMJ include extension, flexion, and translation. The movement of the mandible to the side (laterally) and forward (rostrally) is called translation.
The dietary preferences of different species influence how much translation can occur. The cat, a carnivore, has a very different degree of translation from a cow, an herbivore. Many carnivores barely chew their food and instead use their teeth to rip off small chucks, which they swallow whole. Herbivores, such as a cow, have a greater degree of lateral translation. Moving the lower jaw in a rostral or lateral direction increases the grinding effect of the molars. This lateral movement can be clearly appreciated when examining a cow chewing its cud. Chewing cannot take place on both sides of the mouth at the same time because herbivores have a wider maxilla than their mandible. Because of their coarse plant-based diet, herbivores also require more extensive mastication, to mash and grind their food into smaller particles so they can be properly digested.
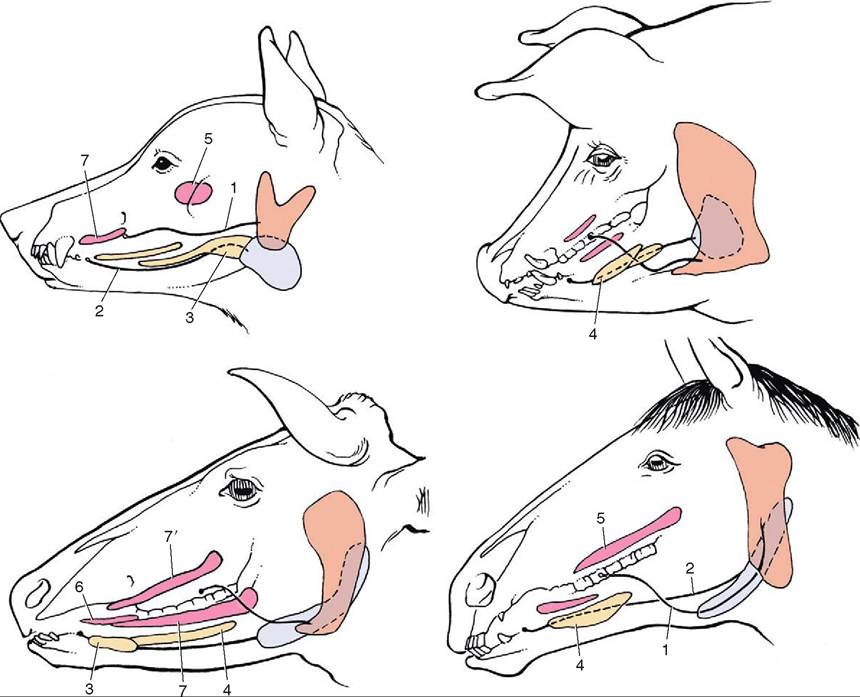
FIGURE 16-11 The major salivary glands of the dog, pig, cattle, and horse. Orange, parotid gland; blue, mandibular gland; yellow, sublingual glands; red, buccal glands. 1, Parotid duct; 2, mandibular duct; 3, compact (monostomatic) part of sublingual gland; 4, diffuse (polystomatic) part of sublingual gland; 5, dorsal buccal glands (zygomatic gland in the dog); 6, middle buccal glands; 7, ventral buccal glands; 7', middle buccal gland. (From Dyce KM, Sack WO, Wenseng CJG: Textbook of veterinary anatomy, ed 4, St Louis, 2010, Saunders.)
TEST YOURSELF 16-3
1. The majority of the tongue is made up of what type of tissue?
2. Besides water, list three substances found in saliva.
3. What are three primary salivary glands in a dog?
4. What does TMJ mean?
5. When speaking of the movement of the mandible, what is translation?
ESOPHAGUS
The esophagus is a muscular tube that connects the pharynx to the stomach. It travels dorsal to the trachea at first, but then moves more to the left as it travels down the neck. After passing through the thoracic cavity it passes through the diaphragm and enters the stomach. The inside of the esophagus is lined by a mucosa that is formed into folds, allowing for the expansion or dilation necessary when food passes through it.
The tunica muscularis consist of two layers of muscle, the inner circular layer and the outer longitudinal layer. Whether the muscle is smooth or skeletal, however, depends on the species. In dogs, horses, and ruminants, the muscle is skeletal throughout the entire length of the esophagus. In cats and primates the distal esophagus changes from skeletal to smooth muscle. Both the circular and longitudinal muscle layers are needed to move food down the esophagus.
PHARYNX
From the oral cavity food enters the pharynx (throat), which is part of both the gastrointestinal tract and the respiratory tract (Figure 16-12). It is here that food is directed into the esophagus through the act of swallowing. The pharyngeal structures are crucial in directing food into the esophagus while at the same time preventing food from entering the larynx and trachea. The epiglottis is the part of the laryngeal cartilage that covers the glottis during the act of swallowing, thus preventing the food from being aspirated into the trachea. The opening of the Eustachian tube is located in the pharynx. The Eustachian tube travels between the nasopharynx and the middle ear. It helps equalize the atmospheric pressure with the pressure in the middle ear. Diffuse areas of lymphoid tissue called tonsils that protect the animal against some diseases are also located in the pharynx.
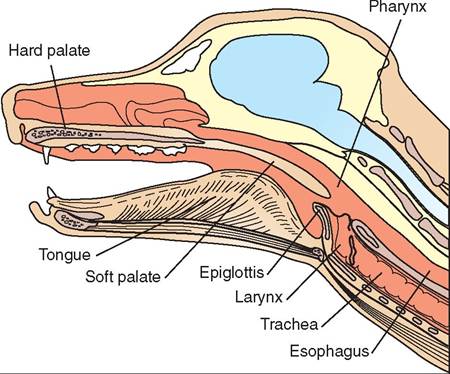
FIGURE 16-12 Longitudinal section of canine head showing the pharynx.
CLINICAL APPLICATION
Myasthenia Gravis
Myasthenia gravis interferes with normal skeletal muscle function and movement. It is an autoimmune disease in which antibodies target the receptors for acetylcholine, preventing the transmission of nerve impulses to the skeletal muscle, thereby preventing contraction. Given that the dog’s esophagus consists of skeletal muscle throughout, myasthenia gravis causes a loss of muscle tone in the esophagus, resulting in an esophageal dilation termed megaesophagus. Food is not properly moved down the esophagus and the dog presents with a history of regurgitation of undigested food. Owners often mistake regurgitation for vomiting. Regurgitation does not involve the profound muscular contractions associated with vomiting. With regurgitation, the food coming up is not digested because it has not yet reached the stomach. Animals with megaesophagus are fed in a position where the head is elevated to allow gravity to assist movement of food into the stomach. Animals with megaesophagus are more prone to aspirating food into the lungs so they must be monitored for signs of aspiration pneumonia.
There is a thickening at the gastric end of the esophagus called the cardiac sphincter that functions to prevent the highly acidic contents of the stomach from backflowing or refluxing into the esophagus and damaging its mucosa.
DIGESTION IN THE ORAL CAVITY
AND PHARYNX
In order to begin the process of digestion food must be brought into the mouth. This is prehension. Different species rely on different anatomic structures such as the teeth, tongue, and lips, as well as movement of their head and jaws, for prehension. Horses have extremely flexible lips, cows have very mobile tongues, whereas pigs use their noses to root.
After getting the food into the mouth, the animal needs to break down the food into smaller pieces that can be easily swallowed. This is accomplished by mastication, or chewing, that involves the lips, cheeks, tongue, teeth, and jaw. In addition to breaking down macromolecules, mastication serves to mix food with saliva, which acts to lubricate and soften the food.
Salivary secretion is regulated primarily by the nervous system. Afferent neurons carry nerve impulses from sensory cells within various parts of the gastrointestinal tract to the salivary center in the medulla oblongata, located in the brainstem. The brainstem then regulates the parasympathetic and sympathetic activity of the salivary glands. Both branches of the autonomic nervous system enhance the secretion of saliva. In many species parasympathetic stimulation produces more watery saliva whereas sympathetic stimulation produces a smaller amount of thick, mucoid saliva. The parasympathetic and sympathetic responses work in combination to produce the consistency and volume of saliva needed. When the animal is consuming a meal, parasympathetic stimulation predominates.
In addition to the sight and smell of food and the presence of food in the oral cavity, saliva secretion can be triggered by a variety of different stimuli, including conditioned responses. Conditioned responses are learned responses in which salivation is initiated by associating a certain unrelated factor or stimulus with the feeding process. Over time, this association strengthens until eventually the unrelated stimulus alone without the presence of food initiates an increase in salivary secretions. This is the learned response that was studied by Dr. Pavlov in his classic experiments in which he conditioned dogs to salivate at the sound of a bell.
Swallowingzdeglutition
Once the food is sufficiently macerated and mixed with saliva it forms a bolus that must now be transported from the oral cavity into the esophagus and ultimately to the stomach. Swallowing or deglutition is the process by which food from the oral cavity is transported to the stomach (in simple-stomached animals) or reticulorumen (in ruminants).
The swallowing reflex occurs in three different phases. The first phase is voluntary, which is why animals require some degree of consciousness to swallow. In an animal with decreased consciousness, the swallowing reflex center is depressed and these animals do not respond well to stimulation of receptors in the mouth and pharynx. This is important in anesthetized animals because it makes them more prone to aspirating saliva into the trachea. In stage one, the voluntary stage, the tongue pushes the bolus toward the pharynx.
Stage two, the pharyngeal stage, is an involuntary reflex that is controlled by the swallowing center in the brainstem. The food bolus stimulates pressure receptors in the pharyngeal wall, causing all openings into the pharynx, except for the esophagus, to close. Once the swallowing reflex begins it cannot be stopped. During this phase breathing stops momentarily and the epiglottis covers the glottis, preventing food from entering the trachea. A wave of muscle contraction moves across the pharynx, pushing the food bolus into the esophagus.
Stage three, the esophageal stage, is also an involuntary reflex. The presence of food in the esophagus stimulates the swallowing center to initiate peristalsis (Figure 16-13A).
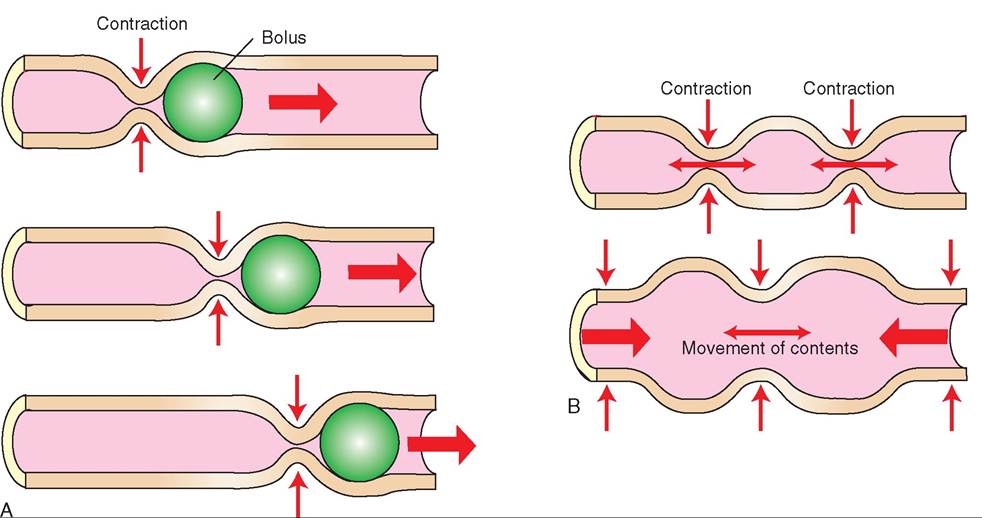
FIGURE 16-13 A, Peristalsis. B, Segmentation.
Peristalsis is the pattern of muscle contraction, involving the circular muscle layer in the esophagus (and other parts of the GI tract), that propels food through the GI tract. In the esophagus, peristaltic contractions travel caudally, pushing the food toward the stomach. Peristaltic contractions consist of a moving wave of luminal constriction cranial to the food bolus. Contraction of the circular smooth muscle causes constriction of the lumen, whereas relaxation of the circular smooth muscle allows the food to pass through the lumen.
The esophagus travels through the thoracic cavity and diaphragm, then connects with the stomach after entering the abdominal cavity. When the food bolus enters the stomach, digestion continues.
TEST YOURSELF 16-4
1. What is the structure that covers the opening of the trachea when an animal is swallowing food?
2. How many muscle layers are found in the esophagus? Which direction do the fibers run in each layer?
3. What is the proper medical term for the throat?
4. What happens during prehension? During deglutina- tion? During mastication?
5. Which of the three phase of swallowing is under conscious control?
6. What is the name of the pattern of muscular contractions and dilations that moves food forward through the esophagus and other parts of the digestive system?
Abdominal cavity
Within the abdominal cavity, the surfaces of the organs are covered by a serous membrane called the visceral peritoneum; the abdominal body wall is lined by the parietal peritoneum. Connectingperitoneum forms folds that connect the organs to the parietal peritoneum and to one another. Mesentery, omentum, and ligaments are types of connecting peritoneum. The mesentery suspends the intestines from the abdominal wall (see Figure 16-2). Many blood vessels and nerves run through the mesentery to supply different sections of the intestinal tract. The mesentery is named according to the organs it suspends. That is, the prefix “meso” is added to the organ name. For example, mesoduodenum is the mesentery that suspends the duodenum and the mesocolon is the mesentery that suspends the colon. The omentum is a double-layered connecting peritoneum that links the stomach to the abdominal wall or other organs. The smaller inner curve of the stomach is called the lesser curvature and it is connected to the first part of the duodenum and the liver by the lesser omentum. The greater curvature is the larger outer curve of the stomach and it is connected to the dorsal abdominal wall by the greater omentum (Figure 16-14). The omentum contains a large amount of fat and it is clearly seen covering the loops and coils of the rest of the intestinal tract when the abdomen is opened. The purpose of the omentum is to store fat, and assist in insulating the abdomen.
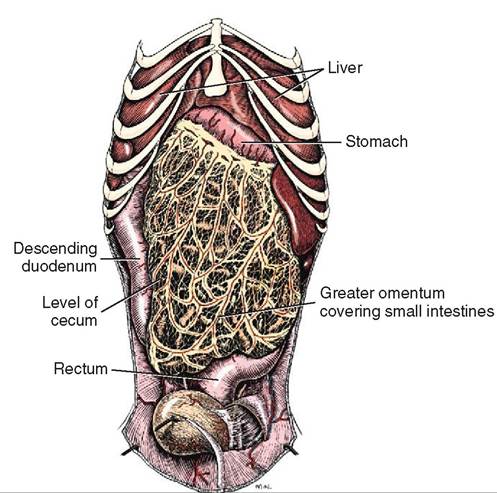
FIGURE 16-14 Greater omentum of male dog, ventral aspect. (From Evans H, de Lahunta A: Miller's anatomy of the dog, ed 4, St Louis, 2013, Saunders>)
STOMACH
BASIC STRUCTURE AND
OVERALL FUNCTION
The functions of the stomach are the storage of ingested food, mechanical and chemical breakdown of food, and production of intrinsic factor, which is required for vitamin B12 absorption in the small intestine. Given that the stomach acts as a storage compartment, animals such as dogs can consume a large meal quickly and then digest the food over a more prolonged time frame. Food that enters the stomach is subject to mechanical digestion that mixes and kneads the food, as well as to chemical digestion, which results in the disruption of chemical bonds by the action of the enzymes and acids secreted in the stomach. Mechanical digestion acts to reduce the size of the ingested particles, which increases the surface area that is available for the enzymes to do their work.
When the food is in a semiliquid state it leaves the stomach and enters the duodenum, where it is called chyme. Chyme is usually hypertonic and has a low pH due to the acidity of the stomach contents. Small amounts of chyme need to be released into the duodenum in a slow and controlled fashion to prevent large fluid shifts from occurring. The hypertonic chyme draws fluids from outside the GI tract into the lumen of the intestine by osmotic forces. If a significant volume of water moved from the vasculature into the GI tract, it could lead to a dangerous drop in blood pressure. In addition, because the chyme is acidic, it requires proper buffering to prevent damage to the duodenal mucosa.
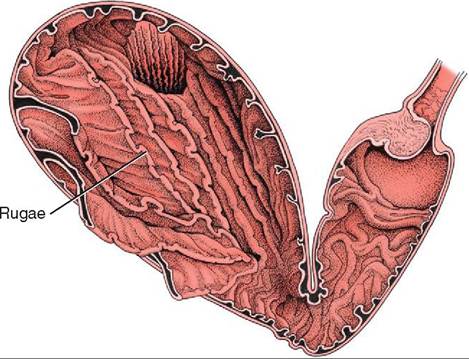
FIGURE 16-15 Gastric rugae in the abomasum. (From Dyce KM, Sack WO, Wensing CJG: Textbook of veterinary anatomy, ed 4, St Louis, 2010, Saunders.)
MONOGASTRIC STOMACH AND DIGESTION
Animals can be divided into two main groups based upon their stomach anatomy. Monogastric animals, including the dog, cat, and horse, have a single or simple stomach with one chamber. Ruminants, such as cows, goats, and sheep, have a complex stomach consisting of four chambers. The monogastric stomach is a C-shaped organ located just behind the diaphragm in the left cranial abdomen. The stomach's main blood supply is from the celiac artery, which is the first branch of the abdominal aorta. Veins leaving the stomach join the portal vein that travels to the liver. The size or volume of the stomach varies depending on how full or empty it is. Rugae are transient folds of gastric mucosa, which allow the stomach to expand when it is filled with food and increase the surface area for absorption (Figure 16-15). The gastric mucosa is made up of simple columnar epithelium containing surface mucous cells that produce a layer of mucus that helps protect the stomach from the acidity of the gastric secretions. If the surface mucous cells are not producing adequate mucus, this can lead to the development of gastric ulcers.
Depending on the species, the luminal surface of the gastric mucosa can be either glandular, nonglandular, or both. Monogastric animals such as the horse and pig have a composite stomach, which means the stomach wall contains both glandular and nonglandular tissue. The carnivore's monogastric stomach contains only glandular tissue. The equine stomach has a clear line of demarcation, called the margo plicatus, dividing the upper nonglandular half and the lower glandular portion. The pig's nonglandular region is much smaller and located where the esophagus enters the stomach.
The glandular portion of the stomach can be divided into three basic regions, called the cardia, fundus, and pylorus (Figure 16-16). In all three glandular regions gastric pits or shallow depressions dot the mucosal surface of the stomach. The gastric pits are the openings of ducts that are lined by glandular cells. The type of glandular cell depends on the region of the stomach. The secretions produced by the
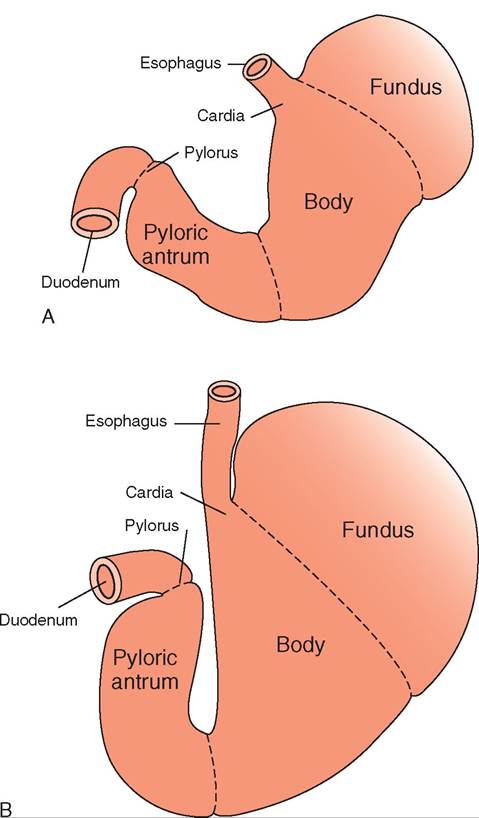
FIGURE 16-16 Gastric anatomy. A, The empty and B, the full monogastric stomach. (From Washabau RJ, Day MJ: Canine & feline gastroenterology, St Louis, 2013, Saunders.)
glandular cells move into the ducts and empty into the gastric lumen.
The cardia is the part of the stomach where the esophagus enters and is so named because of its close proximity to the heart. In this region of the stomach, mucous glands secrete a thick layer of alkaline mucus to protect the mucosa against damage from the gastric acids.
The expanded, dome-shaped, blind-ended sac that is adjacent to the cardia is called the fundus. This is the section where the rugae are prominent so the fundus can expand to store the food after a large meal is consumed. The fundus leads into the body, or corpus, which is the largest section of the stomach. It links the fundus with the distal part of the stomach, called the pylorus.
The gastric pits in the glandular regions of the fundus and the body of the stomach contain different types of glandular cell, including mucous neck cells, parietal cells, and chief cells, each producing different secretions (Figure 16-17). The parietal cells are gastric glands that secrete hydrogen and
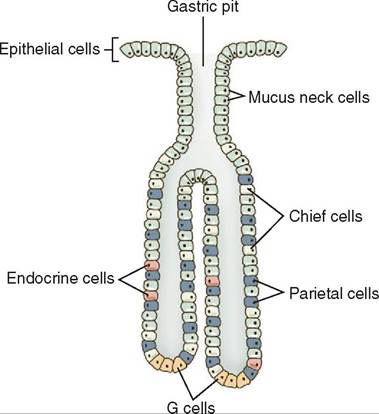
FIGURE 16-17 The gastric pit—the anatomic unit of gastric secretion. (From Washabau RJ, Day MJ: Canine & feline gastroenterology, St Louis, 2013, Saunders.)
chloride, which form hydrochloric acid (HCl) in the lumen of the stomach. Parietal cells also secrete intrinsic factor, which is necessary for absorption of vitamin B12 in the small intestine. hi the cat, the pancreas, not the parietal cells of the stomach, secretes intrinsic factor.
Cells close to the opening of the duct, called mucous neck cells, secrete a thinner, less viscous, mucus than the surface mucous Wls and are considered progenitor cells. THs means they are capable of dividing and creating new cells. The new cells can migrate either up into the mucosal surface or fouwrnther d into the gastric glands, where they can remain mucous Wls or become parietal or chief cells.
edTlhl e thir c type located in the fundic glandular region is the Wef cell, which secretes pepsinogen. Pepsinogen is an inactive precursor form of the enzyme pepsin. It is con- voerted int pepsin by the acidic environment of the stomach created by hydrochloric acid (HCl). Pepsin is a proteolytic enzyme that begins the chemical digestion of proteins. Once pepsinogen is activated to pepsin it can also activate other pepsinogen molecules. Secretion of proteolytic proenzymes such as p^^Wogen is essential in the digestive process because the constant presence of the active enzyme pepsin would lead trhoeeatkdobwn of the very cell that is making it.
lTahnedulalastr g portion of the stomach is called the
pyloric gland region. This region consists of the pyloric antrum, which is the area continuous with the body of the stomach. He stomach then narrows into the pyloric canal, terminating at the pylorus, wnich opens into the duodenum through a circular muscle called the pyloric sphincter. The sphincter helps determine the rate of gastric emptying, which is the rate at which the stomach empties chyme iunotdoetnhuemd. The glands found in the pyloric region
include mucus secreting cells and G cells. G Wls are endocrine Wls that secrete the hormone gastrin into the bloodstream.
STIMULATION OF SECRETIONS
uTbhsrteaencses, acetylcholine,astgrin, and histamine,
sctriemtiuolnaste se by glandular cells in the stomach. Histamine is secreted by enterochromaffin-like cells (ECL Wls) iansttrhice g mucosa, acetylcholine comes from cholinergic neurons, and gastrin is released by G cells. Acetylcholine stimulates both chief cells and parietal cells; gastrin and histamine mainly stimulate the parietal cells. Both acetylcholine and histamine act directly on the parietal cells to increase hydrogen and chloride ion production, whereas gastrin acts iyndirectly b causing the ECL cells to release histamine.
The cephalic phase of secretion bngms Wn an animal anticipates or is preparing to eat a meal. The enteric nervous ismysutelamteids st by a parasympathetic response, leading
to the release of acetylcholine. When acetylcholine binds to its receptors, it directly causes the parietal cells to secrete hydrogen and chloride ions and the chief cells to secrete pepsinogen into the stomach and the G cells to secret gastrin ilonotodstthreabm. Gastrin eventually travels to the pari
etal and ECL cells; it stimulates the ECL cells to release his- tcaemtyilnche.olAine can also trigger histamine release by
eElClsL. c Histamine further stimulates the parietal cells to rperoduce mo hydrogen and chloride ions.
The gastric phase of secretion Lreghis Wn food enters the stomach. Glandular cells are further stimulated by the stomach wall stretching. Formation of peptides by protein breakdown triggers long vagal reflexes to and from the brain, as well as local enteric reflexes. In addition, the acetylcholine released stimulates the secretion of histamine from the ECL caestllrsinand g from the G cells. Gastrin goes on to stimu- lvaetne e more histamine release from the ECL cells, which ieunarsntes incr HCl production. Peptides that are present frreoamkdothwenb of proteins have a direct effect on the
fraeslterainse o g from the G cells, ultimately leading to even more histamine release, with the same overall result of increasing HCl production. As a result, the pH of the stomach can H as low as 2.0.
TEST YOURSELF 16-5
1. The serous me mbrα nethot Coversthe organs of the
abdominal cavity is the____.
2. The cmn nectingneStone umtS atli sks srectomcng to the
abdominal wall is the_____.
3. Semiliquid, partially digested food that leaves the
stomach and enters the duodenum is____.
4. What argthefos r rgctionnona mo sagmarie stomach?
5. What o rotey∣oticm Oizyrsremthe utomagh oegits protein digestion?
6. What three sutionnsgrult from acetylcholine release during the cephalic phase of gastric secretion?
MONOGASTRIC STOMACH MOTILITY
uTshcelem contractions of the monogastric stomach wall coontribute t the mechanical breakdown of food particles itelhl at w b further digested in the small intestine. The
stomach releases gastric contents into the small intestine at a controlled rate. The smooth muscle that makes up the muscle layer of the stomach is an excitable tissue. Specialized smooth muscle cells located within the stomach and intes- tcitne a as pacemaker cells regulating the contraction of the gastric and intestinal smooth muscle, similar to cardiac sinoatrial (SA) node cells that control heart muscle contractions. These pacemaker cells lie at the junctions between the submucosa and the circular muscle and between the longitudinal and circular muscle extending the length of the gut. They do not have constant resting membrane potentials but instead have repetitive, spontaneous, slow fluctuations in their resting membrane potentials. When the changing rmesbtrianngeme potentials of the pacemaker cells reach
threshold they initiate action potentials that result in synchronized smooth muscle contractions. These slow fluctua- teisotinnsgin r potential vary among different parts of the gastrointestinal tract and, although they are spontaneous, tehey can b regulated by the autonomic nervous system.
tTuhaetioflnusc in resting membrane potential do not
always initiate an action potential. Only when the peak of a svleow wa crosses threshold and leads to an action potential duoscelse a m contract. The slow waves therefore cannot coanutrseacctions unless they reach threshold. Acetylcho- loimne fr parasympathetic neurons elevates the baseline rmesbtriannge me potential, resulting in the slow waves
approaching threshold more often. This stimulates an increase in action potentials and consequently more smooth murcle contractions. Norepinephrine released by sympa- tuhreotnics ne does the opposite and lowers the baseline rmesbtrianngeme potential, making it less likely to cross
threshold and therefore reducing the frequency of smooth omnutsrcalcet icons.
When threshold is reached, the opening of voltage-gated Ca2+ chnnnels and entry of calcium into the muscle cell initi- aotnetsrathcteiocn process. A greater frequency of action
potentials means more calcium enters the muscle cell, result- irneagteinr a g force of muscle contraction.
Each part of the stomach differs in its degree of move- dment base on the function it is designed to perform. When an animal eats a meal, the fundus expands and adapts to accommodate large volumes of food without significantly increasing intraluminal gastric pressure. The corpus (body) ies a larg mixing chamber, whereas the pyloric antrum acts,like a pump regulating the movement of food toward the hpiynlocrteicr.sp
eTrhisetapltic contractions that occur in the fundus and corpus are weak but get progressively stronger as food moves toward the pylorus. The peristaltic movements fragment the foood int smaller particles, while also influencing the rate of gastric emptying into the small intestine. As the peristaltic wave gets closer to the pylorus, the pressure in the pyloric liusems en r forcing a small amount of chyme to exit the shteonmach. W the peristaltic contraction reaches the pyloric sphincter, the sphincter narrows and prevents larger particles of food from leaving the stomach. Instead, they are forced back into the corpus, in a process called retropulsion. This aolrlows f more remixing and grinding into small enough particles that will allow them to pass through the pyloric sphincter.
CONTROL OF GASTRIC MOTILITY
iTlihtye mot of the stomach is under neurohumeral control, meaning that both neurotransmitters and hormones can affect the motility of the stomach and the rate of gastric enmptying. I the neural component, the nerve fibers of the vagus nerve synapse on nerve cell bodies of the gastric myen- txeursi.c ple This has a profound effect on the control of iglaitsyt.ric mot The motility of the stomach decreases when food enters it. The motility increases as the food approaches tyhloeripc region. These opposing responses are both the result of vagal (vagus nerve) stimulation. The vagal nerve efilbeearsse r a neurotransmitter other than acetylcholine tuhpaptrsesses movement. Some sources identify this neurotransmitter as vasoactive intestinal peptide (VIP). In the epgyilonric r the vagal nerve fibers release acetylcholine,
xwchitiacthoriys e and increases peristaltic contractions.
eTnhteiodnist that accompanies ingestion of a large meal tortihggers b adaptive relaxation of the fundus and corpus earnidstaplsis in the pylorus.
rTmheo nhaol control of gastric motility by gastrin is excitatory, but its effect on motility is not nearly as pronounced as its effect on secretion of gastric juices.
GASTRIC EMPTYING
faTthee r o gastric emptying is primarily controlled by the fstrength o the pyloric antral muscle contractions and to a lxetsesnetr e the degree of contraction of the pyloric sphincter. The strength of the pyloric antral muscle contractions is a result of interplay between the stimulatory effect of gastrin, secreted by G cells, and the inhibitory effect of the duode- hneunm. W a meal is consumed and the stomach stretches, cbaolth lo and central reflexes are stimulated, leading to the frcelteyalcsehoolinae and causing an increase in pyloric
aunsctlreal m contractions and more rapid emptying of the stomach. Gastrin stimulates contraction of the pyloric aonthtral smo muscle and dilates the pyloric sphincter, tehausisnigncr the rate of gastric emptying.
uTrhoehnuemeral stimulatory effects of the stomach are yalanced by the neurohumeral inhibitory effects of the small ihnetnestine. W food enters the duodenum, multiple neural reeflexes ar initiated that slow the rate of gastric emptying ibtyinignhib the strong contractions of the stomach and increasing the tone of the pyloric sphincter. These neural reflexes involve both short (local, through the enteric nervous system) and long reflex arcs. The long reflex arc consists of a signal sent to the central nervous system, and a response that results in increased sympathetic stimulation and decreased parasympathetic stimulation to the stomach. This reecsruealtssedin d gastric smooth muscle contractions.
uTrhoenanle reflexes are initiated when sensory nerve endings in the duodenal mucosa perceive duodenal distention in the presence of chyme, high concentrations of rpeptides o fat breakdown products, osmolarity of chyme (hyper- or hypo-osmotic chyme empties slower than iso- osmotic chyme), and/or low chyme pH. Not only nervous reflexes, bit also hormones, such as secretin, cholecystokinin (CCK) and gastric inhibitory peptide (GIP), released by the small intestine, play a role in delaying gastric emptying by inhibiting pyloric antral contractions and further constricting the pyloric sphincter. Release of these hormones is most strongly associated with an increase in fatty or acidic cnhteyrminege the duodenum.
CLINICAL APPLICATION
Emesis
Emesis is a protective mechanism that provides animals with the ability to remove harmful or toxic substances from the rtomach or upper intestine. Some species, such as swine, dogs, and cats, vomit easily. The vomiting reflex is controlled by the vomiting center in the medulla. The reflex begins with the animal taking a deep inspiration, followed by closure of the glottis and contraction of the abdominal muscles, but ansottr icthe g muscles. The combination of abdominal
murcle contraction and inspiration results in an increase pressure in the abdomen. This force is transferred to the fcontents o the stomach. The cardiac sphincter relaxes and food is forced into the esophagus. Antiperistalsis mo^s the food up the esophagus and into the oral cavity. Antiperistal- eis is ^rse peristalsis where the peristalsic wave moves epsatretdially dig food toward the oral cavity, as opposed to peristalsis that moves chyme toward the anus.
Horses r are Iy vomit because their cardiac sphincter is so strong and because the angle at which the esophagus enters tohme ascth causes the stomach wall to push against the
sphincter. The cardiac sphincter closes tightly when the stomach is full, making it difficult for it to open from a reverne direction. In the horse, attempting to vomit can increase the pressure in the stomach considerably, resulting in dilation and rupture of the stomach. lruepcoesaeting g monosaccharide units and may be rbranched o unbranched. Sugarsecan b simple monosaccharides (srich as glucose) or made up of two or more monosaccharide units linked together (such as the disaccharides sucrose and lactose). Cellulose is a complex carbohydrate, ewshich mak up the structural part of a plant. Mammalian dnizgyemsteivse e cannot break down cellulose.
Proteins are made up of repeating amino acid units. A protein is defined as a chain of more than 50 amino acids, ohegledthter by peptide linkages. Peptides are chains of fewer than 50 amino acids linked together. For example, odnipseispttides c of two amino acids, tripeptides consist of three amino acids, and so forth.
Luicphids, s as triglycerides, are made up of a glycerol backbone and three fatty acids. Although other lipids are found in food, the majority of the fat found in an animal's doniestiscts of triglycerides.
Imneso animals, such as omnivores, chemical digestion begins with amylase in the saliva. Amylase is an ^aτιe that breaks down starch carbohydrates. Because food is held in trvhaitleyo ca for such a short period of time, starch digestion by sali vary amylase takes place primarily in the stomach. Starch digestion is continued in the small intestine by pan- cyrlaesaet.ic am In animals that do not have amylase in their saliva, the digestion of starch does not begin until the food has reached the small intestine and pancreatic amylase is roetlheased. B luminal and membranous chemical carbohy- edsrtaitoendig then continue within the small intestine.
ePsrtoiotenin, dig on the other hand, begins in the setposminaocghe.nP released by the chief cells is activated
teyopspin b the acidic environment produced by HCl in the stomach. This begins luminal chemical protein digestion, iwllhich w continue in the small intestine. In luminal epsrtoiotenin, dig larger proteins are broken into smaller hpeaipntsid. e c
TEST YOURSELF 16-6
DIGESTION IN THE STOMACH
Mechanical digestion in the oral cavity and in the stomach ionstieaslse t increase the amount of food surface area that can be exposed to the digestive enzymes. Digestive enzymes are respoiesible for the second phase of digestion, chemical digestion. Chemical digestion is divided into two phases: luminal and membranous chemical digestion.
Luminal chemical digestion results in large macromolecules being broken down into short chain polymers through the process of hydrolysis. Hodrolysis is a chemical reaction ihnicwh a bond is broken by the insertion of a water mol- eycdurolel.ysHis is then repeated in membranous chemical
digestion, where the short chain polymers are completely borwonken d into their most basic component parts.
A meal may include carbohydrates, proteins, and lipids as ceensergy sour as well as other nutrients, such as vitamins raanlds. mine Carbohydrates are made up of repeating units called monosaccharides, such as glucose, galactose, and fructose. Starches are large carbohydrates made up of
1. Which neurotransmitter, released by sympathetic neurons, causes a reduction in the frequency of smooth muscle contractions in the stomach?
2. Wh at partofthe mo nogastric stomach increases in size to accommodate a large meal?
3. W hat nerve canelicit ypessite tyses ofgaetric movement through the release of different neurotransmitters in the myenteric plexus?
4. bst thsde eos ditioen iutheduadesumthdtcanCecrease the rate of gastric emptying.
5. What aret2e two types oS diaestioh thactake place in the stomach?
6. Repeating units of monosaccharides make up.
RUMINANT STOMACH AND DIGESTION
uTmheinrant stomach consists of four chambers. It is iomportant t note that the ruminant has only one stomach, even though it is made up of four chambers. The first three ohambers are known as the forestomach or forestomachs.
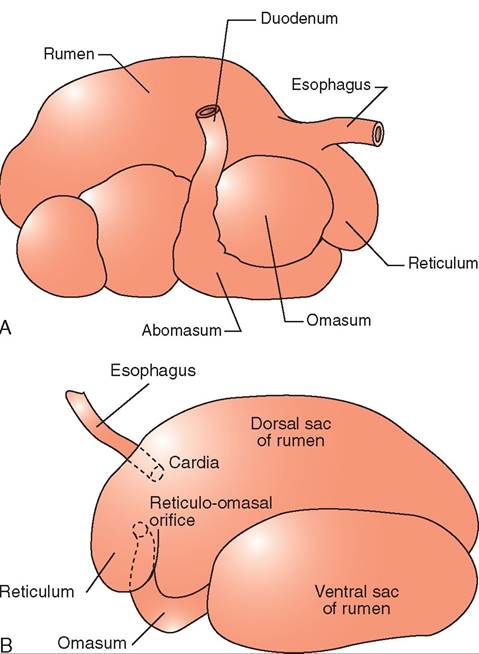
FIGURE 16-18 A, Stomach and forestomachs of a cow as seen from the right side of the cow. B, Rumen topographic anatomy as seen from the left side of the cow.
Some references refer to all three chambers as one forestomach, whereas other references refer to the three chambers as three separate forestomachs. These three chambers, the reticulum, rumen, and omasum, are nonglandular. As food leaves the forestomachs, is enters the fourth chamber or the “true” stomach, the abomasum, which is glandular (Figure 16-18).
Ruminants are herbivores, and their highly fibrous and less nutritive diet requires that they consume greater quantities of food than carnivores. To digest this food properly, ruminants also require a more complex and involved digestion process than carnivores. The forestomachs are lined by stratified squamous epithelium and are nonglandular, meaning they do not produce digestive enzymes. The rumen and reticulum are where the majority of cellulose is degraded, yet because they lack the enzymes necessary to break down the complex carbohydrates (cellulose) that make up the bulk of their diet they need another way to begin the digestive process. The rumen and reticulum contain a large number of microorganisms (bacteria, protozoa, and fungi) that are responsible for a fermentation process that yields adequate nutrition for the ruminant. During fermentation, complex carbohydrates such as cellulose and hemicellulose are broken down into a variety of different end products such as volatile fatty acids that the ruminant absorbs and uses for energy.
The abomasum functions in the same way as the monogastric simple stomach in carnivores. As mentioned previously, the forestomachs in ruminants such as the cow, sheep, goat, deer, reindeer, and moose consists of three chambers: the reticulum, rumen and omasum. There are other ruminants, however such as camels and llamas, that have only the reticulum and the rumen and lack the omasum.
To enter the forestomachs the food must pass through a powerful sphincter that is found at the junction of the esophagus and the reticulum and rumen.
Forestomachs and abomasum
Each of the forestomachs performs a different digestive function, so their anatomic features are different. The rumen is a large expansible chamber where fermentation occurs. It contains many microorganisms that assist in breaking down the carbohydrate substances that make up the structural parts of plants: cellulose, hemicellulose and pectin. The rumen occupies most of the left side of the abdominal cavity and extends from the diaphragm to the pelvis when it is full. This means that the other abdominal organs are located to the right (Figure 16-19). The rumen is commonly referred to as the “paunch” because of its large volume, which in an adult cow can reach 100 L. The ruminal mucosa contains numerous papillae, which help to increase the surface area available for absorption. Pillars (muscular folds) divide the rumen into the dorsal sac, ventral sac, and two caudal sacs.
The reticulum is located cranial to the rumen and is on the median plane, lying against the diaphragm. The reticulum is also called the “honeycomb” because its mucosa resembles a honeycomb, with its crisscross pattern (Figure 16-20). The contents of the reticulum can enter and exit the rumen fairly easily, meaning that the rumen and reticulum essentially act like one unit in which fermentation occurs. The reticulum and the rumen are indeed frequently referred to as one unit, the reticulorumen.
The reticular or esophageal groove (Figure 16-21) links the esophagus with the omasum and plays a crucial role in the young ruminant. When a young ruminant nurses, the groove folds in and essentially turns into a tube for milk to travel directly into the omasum and abomasum, bypassing the reticulum and rumen. If the groove did not close, the milk could spill into the reticulorumen, and the bacteria present could ferment the milk, producing lactic acid. This would decrease the rumen pH, causing acidosis, and inhibiting normal microbial development in the reticulorumen.
The next chamber, the omasum, is also called the many plies or book stomach because of its many leaves, which resemble pages (plies) in a book (Figure 16-22). The leaves are actually folds of mucosa that have many papillae on their surfaces. This increases the absorptive surface area in the omasum, where absorption of water and salts takes place. The omasum is a spherical compartment located on the right side of the abdominal cavity. It connects the reticulorumen to the abomasum.
The abomasum is the elongated “true stomach,” lined with glandular tissue (see Figure 16-15). It is located on the
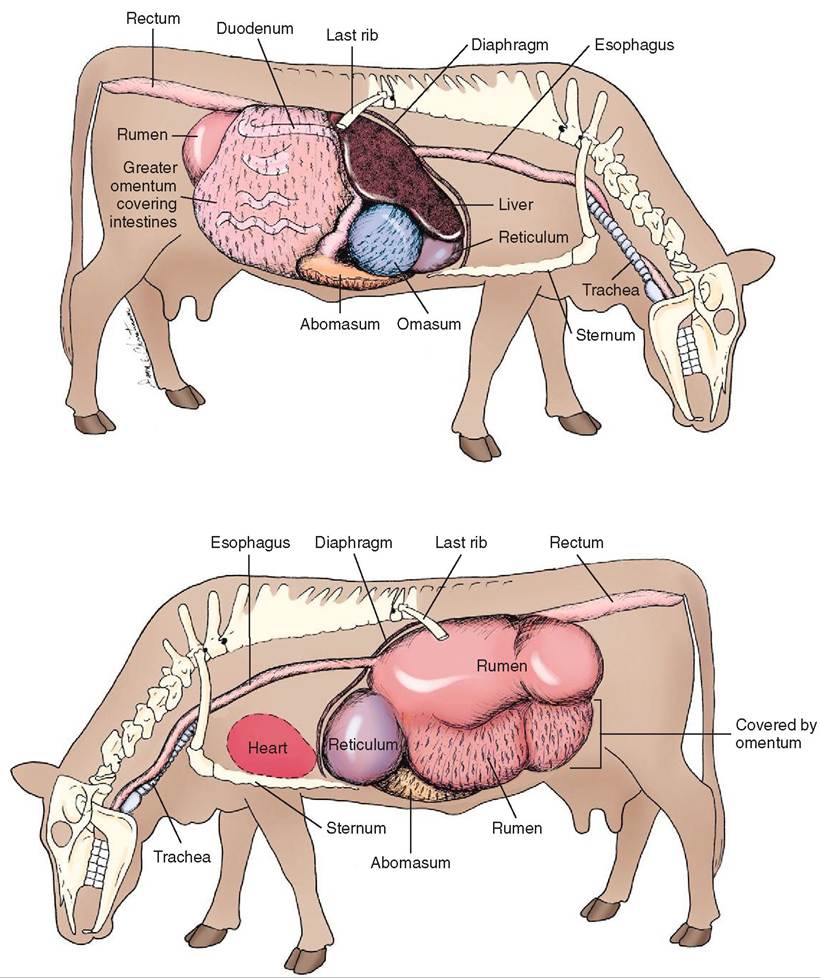
FIGURE 16-19 Bovine abdominal viscera (left and right lateral views). (From Christenson DE: Veterinary medical terminology, ed 2, St Louis, 2008, Saunders.)
right side of the abdomen, in contact with the ventral abdominal wall. The abomasum functions much like the simple stomach in the monogastric animal, except that in ruminants the abomasum does not act like a storage compartment. ⅞e flow of ingesta into the abomasum is continuous. A the abomasum expands it inhibits contractions of the reticιιlornmeιι, limiting the amount of ingesta that can enter the abomasum and keeping the abomasum's volume relatively constant.
Abomasal emptying is controlled in a similar fashion to tnhoegamstoric stomach. The abomasum contains numer- loaunsdgs, which secrete substances such as pepsinogen, and hydrogen and chloride ions, just like the monogastric stomach. hi the young ruminant the enzyme rennin that causes milk protein coagulation is also released. Milk coagulation prolongs the amount of time milk proteins stay in the abomasum, allowing more time for pepsin to break down trhoetepins.
MOTILITY OF THE RUMINANT STOMACH
Digestion in the ruminant begins like that in a nonruminant. oood is consumed and the process of mastication breaks loaordger f particles up into smaller parts, mixing them with saliva to create a rounded mass called a bolus that the
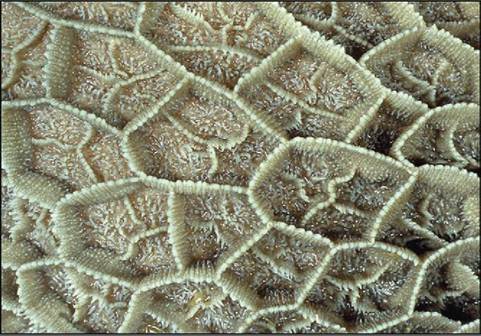
FIGURE 16-20 The mucous membrane lining of the reticulum. Note the honeycomb appearance. (From Dyce KM, Sack WO, Wenseng CJG: Textbook of veterinary anatomy, ed 4, St Louis, 2010, Saunders.)
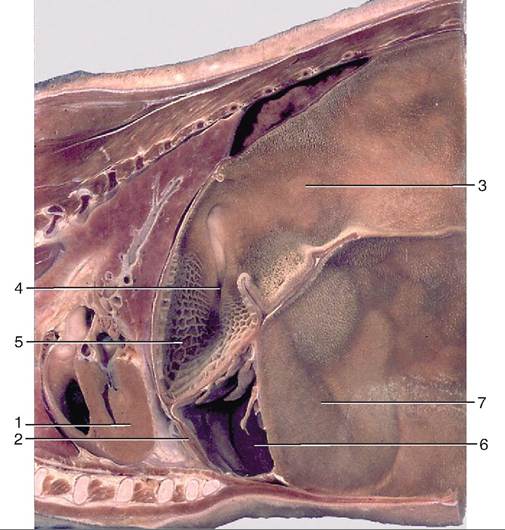
FIGURE 16-21 Paramedian section of part of the trunk of a goat. 1, heart; 2, diaphragm; 3, dorsal sac of rumen; 4, esophageal groove; 5, reticulum; 6, abomasum; 7, ventral sac of rumen. (From Dyce KM, Sack WO, Wensing CJG: Textbook of veterinary anatomy, ed 4, St Louis, 2010, Saunders.)
ruminant swallows. An adult cow can secrete as much as 200 L of saliva per day (the adult human that produces only 1 to 2 L per day). Ruminant saliva is alkaline and contains bicarbonate and phosphate. The bicarbonate helps to neutralize the acids produced by fermentation in the reticuloru- men; the high phosphate content is necessary for both buffering and microbial growth.
The reticulorumen undergoes three types of contraction. The primary contractions or mixing contractions help ensure adequate movement of the contents back and forth between
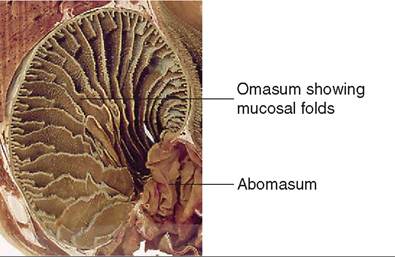
FIGURE 16-22 The mucous membrane lining of the omasum. (From Dyce KM, Sack WO, Wenseng CJG: Textbook of veterinary anatomy, ed 4, St Louis, 2010, Saunders.)
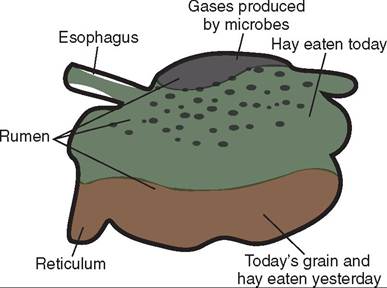
FIGURE 16-23 Layers of material in the rumen. Grain and yesterday's hay are the heaviest materials, so they have fallen to the bottom of the rumen and into the reticulum. (From Colvile J, Oien S: Clinical veterinary language, St Louis, 2014, Mosby.)
the reticulum and the rumen. This serves to separate particles on the basis of size: the heavier, larger particles will fall into the lower portions of the reticulorumen and the smaller, lighter particles move to the top of the rumen to be carried into the omasum (Figure 16-23). The primary contractions also provide the force necessary to move the liquid contents from the reticulorumen into the omasum.
The second type of contraction in the reticulorumen is associated with the process of rumination or chewing the cud. During rumination, the ingesta moves up from the reticulorumen into the oral cavity, where additional saliva and continued chewing help to break down the coarse plant materials further.
The rumination process involves four steps. The first step is regurgitation. During regurgitation the reticulum contracts, bringing material from the dorsal reticulum to the cardiac sphincter. Next the cardiac sphincter relaxes, followed immediately by the animal taking a breath against a closed glottis. This causes negative pressure in the thorax, which pulls open the esophagus and the cardiac sphincter. At the same time, the rumen contracts, increasing pressure within the rumen. Pressure in the rumen is now higher than in the esophagus so the food bolus passes into the esophagus. The bolus is then moved toward the oral cavity by an antiperistaltic wave of contraction (reverse peristalsis) of the esophageal smooth muscle.
Th∣e process of rumination then proceeds with the next three steps: re-insalivation, re-mastication, and re-swallowing. During re-insalivation and re-mastication the ruminant adds more saliva and chews the regurgitated bolus much more thoroughly and completely than the first time it was in its mo^L The number of chews is based on the diet; ruminants that eat fiber-rich forage ruminate over a longer ifpmereiod o t than those fed a low fiber diet.
Rumination is completed when the animal re-swallows the bolus. Re entire time spent ruminating also varies with the type ot' diet that is consumed. For example, animals consuming a mostly hay diet ruminate for approximately 8 ehrours p day. Rumination occurs in a number of distinct rpeeardiods sp out over the entire day.
The third type of reticulorumen contraction is respon- soirble f the release of gases, such as carbon dioxide and rmoedtuhcaende, p during fermentation. This process is
called eructation and is how ruminants belch. These contractions are known as secondary contractions. CaAon dioxide is produced as the microorganisms break down carbohydrates and amino acids. Methane-producing bacteria then use the carbon dioxide to produce methane. Both gases start to accumulate in the reticulorumen during the fermen- traotcieosns p and need to be released, or the ruminant could develop a con d i tion known as bloat (acute rumenal tympani). When bloat occurs the accumulating gases build up, causing the reo'cιιlorumeιι to expand. The expansion compresses the thoracic and abdominal organs and compromises blood flow to abdominal organs. It also interferes with lung function by rpeuststuinreg p on the diaphragm. Tissue hypoxia (inadequate tissue oxygenation) and death of the ruminant can follow if the bloat is not treated.
The amount of gas produced by fermentation in a high yielding dairy cow over a 24-hour period can be as high as 4000 L. These gases must be removed through eructation yefore they can build up. Eructation occurs about once a minute.
Eructation begins with secondary contractions in the caudal blind sac of the rumen. The contractions move crani- anlgly alo the dorsal sac. They force air forward toward the cranial part of the reticulorumen. As this is occurring, the folds ot' the rumen wall keep the contents away from the fopening o the esophagus. The cardiac sphincter and lower elsaoxphagus r and air is able to enter the relaxed esopha
gus. The gas is then propelled up the esophagus by antiperi- sotnatltriacctcions. Rather than producing a burp, most of taehscteegd is dir into the lungs during inspiration and then eoxupgehlled thr expiration.
Although the enteric nervous system plays a part in regulating the motility of the ruminant digestive system, longer roemflexes fr the central nervous system play an even more iomleportant r in coordination of normal motility. There is eaan ar in the brainstem that is responsible for control of reticuloruminal motility. The area is innervated by vagal ravffeerent ne fibers that receive input from intraluminal sceantesdors lo in the reticulorumen. These sensors monitor conditions such as pH, osmolarity, and expansion of the iwthaillns w the reticulorumen. The brainstem sends efferent soiuggnhals thr the vagus nerve to the smooth muscle of tehtiecurlorumen, controlling its contractions. Because the rvvaegus ne carries both afferent and efferent fibers, these reflexes are often referred to as vagovagal reflexes. Tine brain- setceemiveaslso r nerve impulses from other parts of the buochdy, s as the olfactory organ and taste buds in the oral cavity, that can also affect the reflex.
TEST YOURSELF 16-7
1. List the Iourchambersthat make Uptheruminant stomach.
2. Vahich oftbubauh chaaιbatsis Uhelargastfeumentation chambat aed oe which sida of tha aeimal would you look to saa Whathar it was bloatad?
3. Which cheobe a is known as the many plies?
4. W hut io tbeim2ortanchoftbs asopoagaal groove?
5. Explaie tha diffataeca batwaae auuctatioe aed uumieatioe.
6. What dwes a cow drn whaws hec hewa her cud?
RETICULORUMEN ECOSYSTEM
oTbhiealmicr species that inhabit the reticulorumen consist of a wide variety of bacteria, protozoa, and fungi that help cormeaptleexa c ecosystem within the reticulorumen. The reticulorumen must maintain the proper balance among the ydpifefesrent t of bacteria, protozoa, and fungi to promote tehrme efntation process. This delicate balance is largely cyontrolled b diet and how much food the ruminant con- sourmes. F example, when there is an increase in grain fcontent o the diet, an imbalance is created in the reticuloru- hmeen. T microbes that break down starch flourish whereas microbes that break down the complex carbohydrates are diminished.
uWmhiennantht e r consumes its food, a small amount
of oxygen is ingested. This oxygen is quickly used up by some of the microorganisms, creating a low oxygen environment in the reticulorumen. This low oxygen environment allows facultative anaerobic bacteria and protozoa, which are crucial players in the fermentation process, to thrive.
uTmhebenr of microorganisms inhabiting the reticulo- rumen is large and the interaction among them is complex, buumtianrant is not born with this ecosystem already estab- lhisehned. W a ruminant is born, its gastrointestinal tract is eursiulea.lly st The microorganisms that will one day be so corucial t its survival must be acquired. The young ruminant attains these different microbes by ingesting them from the environment. The interactions that occur between mother aonudngy, such as during grooming, nursing, or cleaning, chieliltpatfea this transfer of microorganisms. By the time tohuengy animal is weaned, its microbial environment is
established and contains a large variety of bacteria, protozoa, ia.nd fung
eTrheentdiff types of bacterium are classified on the fbhatsis o w nutrients they metabolize and the substances
they produce. For example, amylolytic bacteria break down starch and soluble carbohydrates into volatile fatty acids (VF)V). Cellulolytic biaCTeria break down components of cell walls, including cellulose, hemicelluloses, and pectin, into VVA. ^e protozoa in the reticulorumen are unicellular, anaerobic, and mostly ciliates. In terms of size, ciliates are much larger than most bacteria. The importance of cil- iates in the digestion process of the ruminant is unclear but they do produce volatile fatty acids, carbon dioxide, lactate, and hydrogen from different varieties of plant. The ruminant can live without protozoa in its gastrointestinal trurotatcotzoba p are thought to play a role in slowing
down the digestion starches and proteins that are rapidly fermented. The number of fungi that make up the final fportion o the microbial population is small but they are tohought t play a crucial role in the breakdown of the eplllant c wall.
CARBOHYDRATE DIGESTION
IN RUMINANTS
The ruminant’s natural diet consists mostly of grasses or roughage and some grains. The grasses or roughage are com- fopmospedlexo c carbohydrates such as cellulose, hemicelluloses, and pectin, which makeup the structural components of the plant and provide the main nutrients for the rumi- riant. Grains, also known as concentrates, are primarily composed of carbohydrates that are nonstructural. These nonstructural carbohydrates include energy stored in the form of starches, fructosans (hydrolyzed to fructose), and suigmarpsl.e s Flthough most mammals produce enzymes tehat ar capable of breaking down starches and soluble rseugars, the are no known mammalian enzymes that can borwenak d complex carbohydrates. In order to survive on trhoeuisr fib diet, ruminants have evolved a symbiotic rela- ittiohnship w the microorganisms living in their digestive shyestems. T microorganisms use the nutrients the ruminant coornsumes f their own growth and development. The microorganisms then produce their own end or waste products that ruminants can use for their own growth and development. In addition the ruminant can digest the microorganisms themselves as a source of protein.
Ientictuhleo rrumen, cellulase enzymes bound to the
fseullruflaocleyst ioc c bacteria break down complex carbo-
ohyudgrhates thr hydrolysis, releasing specific monosaccharides (simple sugars) such as glucose and less complex polysaccharides (ITiiilhpIe sugars linked together). Starches aunbdle sol sugars are also broken down into monosaccha- roildyesascacnhdarpides through hydrolysis by the amylo-
lcytteirciab. a These initial steps do not take place in the
microorganisms and the saccharides released are not imme- vdaiailtaeblylea to the ruminant. Instead, the saccharides are aybsorbed b the microorganisms and further metabolized to spyruvate. F this process is occurring nicotinamide adenine dinucleotide (NAD) is reduced to NAD hydrogen (NADH) ond two molecules of adenosine triphosphate (ATP) are produced. Because the reticulorumen has an anaerobic environment, the pyruvate is converted into volatile fatty acids (VFAs) Hich as acetic acid (acetate), propionic acid (propionate), and butyric acid (butyrate).
When an animal consumes a diet high in starch, the total production of VFAs is increased, the proportion of propionate is increased, and the VFAs are produced faster than twhen a die is high in fiber. This illustrates what a profound eofemfdepctosfitionc has on VFA production.
TAhs e VF that are produced in the forestomachs are absorbed into the blood. Most of the VFAs are absorbed fertoicmulotrhuemern, some from the omasum and
abomasum, and the last small bit of absorption is completed lin the smal intestine. The VFAs are the main source of eonr ergy f the ruminant, functioning in the same role as golrucose f monogastric animals.
LIPID DIGESTION IN RUMINANTS
Lipids constitute a small portion of the diet of ruminants. iDthiets w too many lipids result in decreased appetite, reduced motility of the reticulorumen, and decreased fermentation of cellulose. Lipids can be found in grasses and plants in the form of triglycerides, glycolipids, and a small famount o free fatty acids.
Microorganisms in the reticulorumen rapidly hydrolyze triglycerides. During this process free fatty acids are sepa- roamted fr the glycerol backbone of the triglyceride. The glycerol then goes through the process of fermentation, pro- dAus,cing VF whereas most of the fatty acids go through the fypdrorocegsesnoatihon. The hydrogenated fatty acids leave
tohme ascth and are absorbed in the small intestine. The
absorbed fatty acids are ultimately used by the ruminant to pnreorgdyuce e or are stored.
PROTEIN DIGESTION IN RUMINANTS
Iunmtihneanr t, ingested food travels through the reticulo-
rumen and then moves into the omasum and abomasum. This means the fermentation process occurs before enzy- emstaitoicn dig and that the nutrients in the food are first aovailable t the microbes living in the reticulorumen. A majority of the protein in the ruminant's diet is actually degraded and used by the microorganisms to meet their own metabolic needs. Peptidases relrcised by rumen bacteria borwenak d proteins to form small peptide chains. These semptaidllep chains are then absorbed by the microorganisms and further hydrolyzed inside the cell, culminating in trhoed upction of amino acids. Some of the amino acids are
yused b the bacteria to create microbial proteins. If the caimdsino a are not used to make microbial proteins, they techaenrb fur broken by the microbes, which deaminate the caimdsino a into ammonium. Ammonium is ultimately used to produce VFAs, which are required by some microbes as growth factors.
Breakdown of peptides by extracellular peptidase is only yone wa in which microbial amino acids are produced, ihcorowbeevser. M can make amino acids from ammonium. Iunrnt, ammonium can be produced by microbes not only teharmoiungahtidon of amino acids, but also by conversion
of non-protein nitrogen (NPN) compounds. Tliese NPN compounds may be endogenous, such as urea, or found in the diet as nitrates. Ruminants ingest nitrates because they are used as plant fertilizers. In fact, NPN compounds are sometimes added to a ruminant diet because this is cheaper than adding more expensive dietary protein. The microorganisms in the reticulorumen are then required to synthesize the protein needed.
In the ruminant, proteins synthesized by microbes are transported to the small intestine where they are broken down by proteolytic enzymes in a similar fashion to that in the carnivore small intestine. Bypass proteins, which are proteins that pass through the forestomachs unaltered, are also degraded enzymatically in the small intestine. The end products of protein breakdown are amino acids, which are absorbed across the intestinal mucosa into the bloodstream and transported to the liver. In the liver, the absorbed amino acids can be metabolized to make proteins. As the amino acids are metabolized, urea is produced as a waste product. The liver also produces urea from endogenous proteins as well as from ammonium absorbed from the rumen. In monogastric animals the kidneys are responsible for excreting urea but in the ruminant the urea travels back to the rumen and ultimately the saliva, where it is available to be used by the microbes once again to make microbial proteins.
GLUCOSE PRODUCTION IN RUMINANTS
In the ruminant, fermentation in the reticulorumen occurs before enzymatic digestion in the small intestine. The ruminal microbes process most carbohydrates and proteins before they are exposed to the intestinal digestive enzymes. In fact, almost no ingested carbohydrates make it to the small intestine. This prevents their exposure to intestinal digestive enzymes and subsequent breakdown into monosaccharides. As a result, the ruminant is constantly faced with a potential glucose deficiency.
Although the ruminant receives most of its energy from metabolizing the VFAs produced by microbes, a source of glucose is still required by some tissues. To meet this need, the ruminant has developed a very effective method of producing and conserving glucose. Almost all of the glucose the ruminant needs is made through gluconeogenesis, production of glucose by the liver from noncarbohydrate sources. In the case of the ruminant, the noncarbohydrate source is the VFA propionate.
Propionate, a waste product of microbial metabolism, is absorbed from the rumen into the blood and travels directly to the liver where it is used to make glucose. Propionate never enters the systemic circulation. Most butyric acid, another VFA, is absorbed and metabolized by the liver into beta-hydroxybutyrate, a ketone. This molecule is easily metabolized by most tissues in the body but cannot be used to make glucose. Acetate, which is the most abundant VFA in the circulation, is absorbed and passes through the liver mostly unchanged, but it also cannot be used to make glucose. Both acetate and butyric acid are used in the mammary gland to synthesize fatty acids.
Propionate is not the only substance to undergo gluconeogenesis. Amino acids that are absorbed from the intestinal tract can also be used to make glucose through gluconeogenesis.
Producing glucose more efficiently is only one of the strategies that the ruminant uses to ensure adequate glucose levels. The other strategy involves conserving glucose more effectively. In the ruminant, glucose is not used to make fatty acids. Fatty acids are made in adipose tissue from acetate rather than in the liver as in other species. In the ruminant this frees up glucose to ensure a supply of glucose will be available for those tissues that require it.
TEST YOURSELF 16-8
1. The main source of energy in ruminants is the production of.
2. In the reticulorumen complex carbohydrates are broken
down to___ or___.
3. What are the three most important volatile fatty acids produced by conversion from pyruvate?
4. Lipids are found in grasses and plants in the form of (three compounds)?
5. Proteins in ruminant diets are broken down extracellu-
lary by peptidase into____.
6. Urea produced as a by-product of amino acid metabolism in the ruminant liver goes to the for reuse.
7. What is gluconeogenesis and where in the ruminant does it mostly take place?
SMALL INTESTINE AND ASSOCIATED STRUCTURES
BASIC STRUCTURE AND FUNCTION
The small intestine extends from the pyloric sphincter of the stomach to the beginning of the large intestine. It is essentially a tube that carries the chyme away from the stomach and deposits it in the large intestine. The small intestine consists of three distinct parts: duodenum, jejunum, and ileum. In those animals that do not rely on large amounts of fermentation to supply their need for energy and nutrients, the small intestine is the principal site for digestion and absorption.
The intestines are suspended from the body wall by the mesentery. The duodenum is the first portion of the small intestine. It receives the chyme as it exits the stomach through the pyloric sphincter. From the pyloric sphincter, the duodenum extends in a somewhat cranial direction for a short distance before turning caudally and descending on the right side of the abdomen (descending duodenum) toward the right kidney. The duodenum then turns medially and moves in a cranial direction again for a short distance (ascending duodenum), before ending at the duodenojejunal flexure where it attaches to the jejunum (Figure 16-24).
The pancreas runs right alongside the descending duodenum and next to the greater curvature of the stomach (Figure 16-25). It consists of an endocrine portion, which
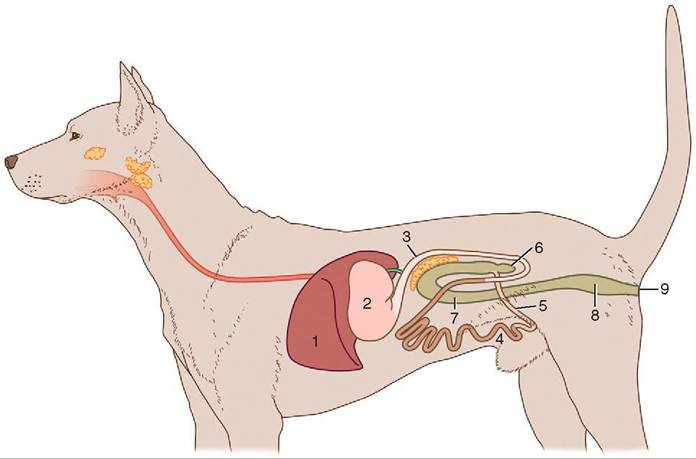
FIGURE 16-24 Schematic representation of the intestines in a dog. 1, Liver; 2, stomach; 3, duodenum; 4, jejunum; 5, ileum; 6, cecum; 7, colon; 8, rectum; 9, anus.
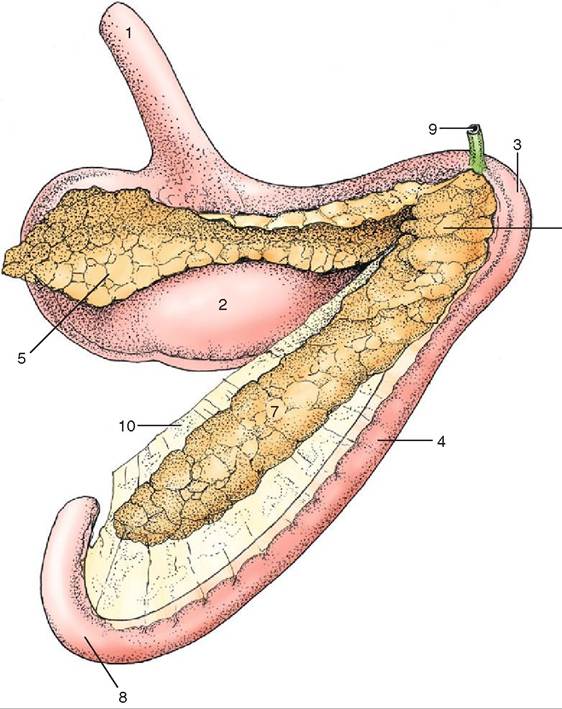
FIGURE 16-25 The pancreas of the dog (caudal view). 1, Esophagus; 2, stomach; 3, cranial flexure of duodenum; 4, descending duodenum; 5, left lobe of pancreas; 6, body of pancreas; 7, right lobe of pancreas; 8, caudal flexure of duodenum; 9, bile duct; 10, mesoduodenum. (From Dyce KM, Sack WO, Wenseng CJG: Textbook of veterinary anatomy, ed 4, St Louis, 2010, Saunders.)
produces various hormones and deposits them directly into the bloodstream, and an exocrine portion, which produces digestive proenzymes that are deposited through a duct into the duodenum. More on this organ later.
The jejunum continues the intestinal tract and is the longest part of the small intestine, forming a mass of many coils and loops that occupy the ventral abdominal cavity. The bulk of chemical digestion and absorption occurs in the jejunum. Its mesentery is long and allows the jejunum to have a great range of motion.
The small intestine ends with the ileum, which is indistinguishable grossly from the jejunum. Peyer’s patches, which are aggregates of lymphoid tissue, are found throughout the small intestine and are especially prominent in the ileum. These structures help protect the animal from disease by controlling local populations of bacteria, functioning in antibody production, and aiding in filtration of fluids. The ileum empties into the large intestine at the cecum (ileocecal junction) in the horse, at the colon (ileocolic junction) in the dog and cat, and at the cecum and colon (ileocecocolic junction) in the ruminant and pig.
To assist the small intestine in digestion and absorption, several features help increase its surface area. First, the small intestine is long and consists of many loops and coils. Second, the mucosa lining the small intestine is thrown into folds or plications. These plicae are permanent structures of the small intestine, in contrast to gastric rugae (the folds of the gastric mucosa), which disappear when the stomach fills. Third, the intestinal mucosa is made of simple columnar epithelial cells that have fingerlike projections called villi, which in turn contain smaller projections called microvilli (Figure 16-26). The villi move back and forth, helping move liquid contents into close contact with the mucosa. The microvilli form what is called the brush border (see Figure 16-2) that provides the majority of the surface area of the small intestine. Embedded in this brush border are brush-border enzymes that are responsible for membranous
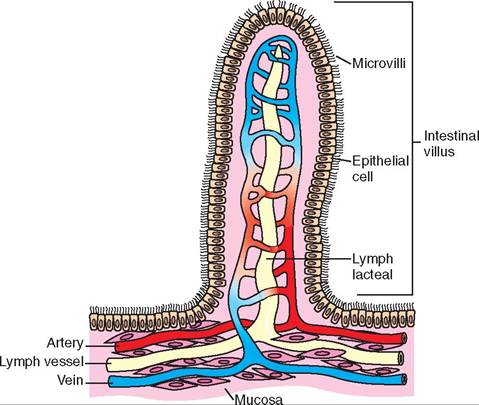
FIGURE 16-26 Microanatomy of intestinal villus.
digestion. Membranous digestion in the small intestine plays the ultimate role in the digestion of carbohydrates, proteins, and nucleotides.
Between adjacent villi are found groups of undifferentiated cells, in an area called the intestinal crypts (or crypts of Langerhans). These undifferentiated cells are the only intestinal cells undergoing cell division and they replace those further up on the tips of the villi as they are lost due to wear. An extensive network of blood capillaries is found inside the villi. These capillaries collect some of the nutrients absorbed from the gastrointestinal tract and transport them to the liver. Lymphatic capillaries called lacteals also are found in the villi (see Figure 16-26). Lipids are too large to enter the blood capillaries and must be transported in the lacteal. The lacteal carries absorbed lipids and fat-soluble substances to the thoracic duct, which empties into the venous blood of the vena cava.
SECRETIONS OF THE SMALL INTESTINE
Many secretions are released from the small intestine that contribute to the digestive process. The duodenal mucosa, for example, secretes two very important hormones, cholecystokinin (CCK) and secretin. The stimulus for secretion of CCK is the presence of chyme with a high amino acid concentration, high fatty acid concentration, or a low pH entering the small intestine. The role of CCK is multifaceted. First, it inhibits gastric emptying. This allows the chyme to exit the stomach at a controlled rate, allowing more time to neutralize the acidic chyme, which in turn protects the small intestine. Second, CCK causes increased secretion of pancreatic digestive proenzymes as well as bicarbonate (HCO3-) secretion. Third, it is the main trigger for gallbladder contraction. Finally, CCK stimulates the secretion of enteropeptidase from the duodenal mucosa. Enteropeptidase is responsible for converting one of the pancreatic proenzymes, trypsinogen, into its active form, trypsin.
Secretin, another hormone released by the duodenal mucosa, decreases the HCl production in the stomach, and increases pancreatic and biliary bicarbonate secretions. The stimulus for secretin release is the presence of chyme with a low pH or high fatty acid concentration. The digestive enzymes, which are released from the pancreas, function optimally when the pH is more alkaline than the acidic chyme as it exits the stomach. The release of bicarbonate from the pancreas and liver helps neutralize the acids leaving the stomach, ensuring that the pancreatic digestive enzymes are most effective.
Pancreas
The pancreas, as mentioned earlier, has both endocrine and exocrine functions. The endocrine part of the pancreas consists of the pancreatic islets (formally known as islets of Langerhans). These pancreatic islets contain several different cell types and each cell type produces its own hormone. For example, beta cells secret insulin, which is needed to transport glucose into most cells, thereby lowering the glucose levels in the blood. Alpha cells produce glucagon, which has the opposite effect, raising blood glucose levels by stimulating processes such as gluconeogenesis.
The exocrine portion of the pancreas consists of groups of acini. An acinus is made up of a single layer of columnar epithelial cells, resembling a cluster of grapes, surrounding a lumen. The cells of the acini release their secretions into the lumen, which merge with other lumina to form ducts that eventually converge into the pancreatic duct. The pancreatic duct empties into the duodenum. Some species have one pancreatic duct whereas others have two. In humans, sheep, and goats, the pancreatic duct combines with the common bile duct just before it enters the duodenum.
The secretions of the exocrine pancreas contain bicarbonate and pancreatic digestive enzymes. Pancreatic cells have surface receptors that are stimulated by acetylcholine, cholecystokinin, and secretin. During the cephalic and gastric phases of digestion, anticipation of food and food stretching the stomach causes an increase in pancreatic secretions. This is brought about mainly by an increase in vagal activity (acetylcholine release). During the intestinal phase of digestion, both a neural and an endocrine stimulus increase pancreatic secretions. Distention of the duodenum results in vagal stimulation of the pancreatic cells (neural stimulation), whereas chyme entering into the duodenum stimulates the release of secretin and cholecystokinin (CCK) (hormonal stimulation).
Enzymes derived from the exocrine pancreas are vital to the digestion of proteins, carbohydrates, lipids, and nucleic acids. Lipases break down lipids into two free fatty acids and a monoglyceride. Pancreatic amylase breaks down starches primarily into maltose (two glucose molecules linked together). Nucleases break down nucleic acids into nucleotides. Finally, proteolytic enzymes, also called proteases, break down proteins into amino acids.
All of the proteolytic enzymes are secreted by the pancreas in an inactive form called a proenzyme (or zymogen). These proenzymes need to be activated in the lumen of the intestine, just like pepsinogen, the proenzyme released by the chief cells in the stomach. The pancreatic proenzymes are trypsinogen, chymotrypsinogen, proelastase, and procarboxypeptidase A and B. They are activated as follows: CCK stimulates duodenal mucosal cells to secrete enteropeptidase. In the duodenal lumen, enteropeptidase then activates trypsinogen into trypsin, its active form. Trypsin is then responsible for activating the other proenzymes released by the pancreas. For example, chymotrysinogen is activated into chymotrypsin, proelastase is activated into elastase, and procarboxypeptidases A and B are activated into carboxypeptidases A and B. Trypsin can also activate additional trypsinogen to trypsin.
LIVER, BILE DUCT, AND GALLBLADDER
The liver is located near the stomach, immediately caudal to the diaphragm (see Figure 16-24). Its secretions are essential
TEST YOURSELF 16-9
1. List, in order as they leave the stomach, the three sections of the small intestine.
2. What is the brush border in relation to the small intestine, and what is its function?
3. What characteristics of chyme are necessary to stimulate the release of cholecystokinin (CCK)?
4. Which hormone, released in the duodenum, is responsible for decreased hydrochloric acid production in the stomach?
5. What substance, released by the pancreas and liver into the duodenum, helps neutralize the acidic chyme leaving the stomach?
6. What are two substances secreted by the exocrine portion of the pancreas?
for digestion and absorption of nutrients. The liver has a multitude of other functions not directly related to digestion. These functions include synthesizing nutrients and regulating their release into the bloodstream, excreting toxic substances, both those originating within the body and those that have been brought into the body, and producing most of the plasma proteins, cholesterol, and many of the blood coagulation factors.
The liver is the largest digestive gland in the body and, like the pancreas, is considered an extramural gland, meaning it is outside the lumen of the gastrointestinal tract. It has two surfaces, a diaphragmatic surface that is the convex surface in contact with the diaphragm, and a visceral surface in contact with the abdominal organs. The falciform ligament attaches the liver to the diaphragm, whereas the right kidney, stomach, duodenum, colon, and jejunum leave impressions on the visceral surface. The mammalian liver consists of lobes. For example, carnivore livers have six lobes: left lateral, left medial, quadrate, right medial, right lateral, and caudate lobes. The caudate lobe has two parts: a papillary process which is on the left of the median plane lying in the lesser curvature of the stomach and a caudate process, which is more caudal on the right side of the liver, enclosing the cranial pole of the right kidney in the right dorsal abdomen. The gallbladder is located between the quadrate and the right medial lobe. Microscopically each lobe of the liver is made up of many hepatic lobules.
The liver is strategically placed between the vessels draining the intestine, and associated structures, and the general circulation. This means that the liver is able to process the blood leaving the gastrointestinal tract, preventing harmful substances, such as toxins, from entering the general circulation. The liver receives blood from two sources, the hepatic portal vein from the gastrointestinal tract and the hepatic artery, a branch of the celiac artery. The celiac artery branches directly off the abdominal aorta and supplies oxygenated blood to the liver tissue. Both the hepatic artery and the hepatic portal vein enter the liver near the periphery of the lobules, in areas known as triads, so called because the bile ducts also are found there in addition to the hepatic artery and hepatic portal vein. The hepatic artery and the hepatic portal vein travel from the periphery of the lobule toward its interior, coming together and emptying their blood into sinusoids, which are the large capillaries of the liver. The endothelial cells making up the walls of the sinusoids have large fenestrations or pores in them so that they are much more “leaky” then normal blood capillaries.
Adjacent to the sinusoids are hepatocytes, which come into close contact with the blood. The pores in the walls of the sinusoid endothelial cells allow large molecules, such as proteins, made by the hepatocytes to enter the blood easily.
Attached to the inner surface of the sinusoids are macrophages called Kupffer cells, which engulf foreign substances. The blood from the sinusoids enters the central vein, which is located at the center of each lobule, and ultimately exits the liver, returning back to the general circulation via the hepatic vein. Located between the individual hepatocytes are bile ductules called bile canaliculi. Bile is excreted from the hepatocytes into these canaliculi and travels in the opposite direction from the blood, away from the central vein, emptying into larger bile ductules, and ultimately into the bile duct at the triad. Bile is necessary for lipid digestion in the intestines (Figure 16-27).
Bile exits the liver and travels through the hepatic ducts, emptying into the common bile duct. The cystic duct also comes off the common bile duct. It connects the gallbladder, if the species has one, to the common bile duct. The common bile duct can empty directly into the duodenum, but in some species it combines with the pancreatic duct before entering the duodenum at the major papillae. There is a sphincter, called the sphincter of Oddi, which controls the entrance of the bile duct into the duodenum. When the sphincter of Oddi is closed, bile backs up into the gallbladder (Figure 16-28).
The gallbladder concentrates and stores bile until it is needed. The cat and dog usually eat meals once or twice a day so fat digestion does not occur continuously. In these animals, the gallbladder stores bile until the animal eats a meal. When fats are consumed, a large quantity of bile enters the duodenum. The gallbladder also provides the liver with a storage container so it can continue to excrete waste products through bile production even during periods when the animal isn't eating.
When chyme with a high fat or peptide concentration enters the small intestine, cholecystokinin (CCK) is released from the duodenal mucosa. CCK is the major stimulus for contraction of the gallbladder and relaxation of the sphincter of Oddi. This allows bile to enter the duodenum.
Given that ruminants have a more continuous digestive process, their gallbladders have a short retention time and do not concentrate the bile as much as that of carnivores. Horses do not have gallbladders. Because they also have a more continuous digestive process that requires a more continuous secretion of bile into the duodenum, bile storage isn't needed.
Bile is composed primarily of bile salts, phospholipids, cholesterol, and bile pigments. Hepatocytes form bile acids from cholesterol. Bile acids contain a water-soluble (hydrophilic) side and a lipid-soluble (hydrophobic) side. Bile acids tend to aggregate and position themselves with their hydrophobic parts facing inward, away from water, and their hydrophilic parts pointing out, toward water. This configuration is called a micelle. In the digestive process, these micelles in the intestine are important because they emulsify lipids, allowing the lipids to come close to the intestinal mucosa so they can be absorbed.
BILE FORMATION AND BILIRUBIN EXCRETION
Bile production begins with bile salts (conjugated bile acids) being secreted into the bile canaliculi. The bile salts draw water out of the hepatocytes by osmosis, making bile a liquid. The composition of the bile is altered as it passes through the canaliculi by both absorptive and secretory processes; that is, water and ions are moved into and out of the canaliculi. One of the secretory processes involves secretin, which stimulates duct cells to secrete bicarbonate, which is added to the bile. The hepatic bicarbonate, along with the pancreatic bicarbonate, helps to neutralize the acid chyme entering into the duodenum from the stomach.
When the bile is released into the intestine, it emulsifies fats. The bile salts themselves are not reabsorbed until they reach the ileum. After they are reabsorbed in the ileum, they enter the hepatic portal vein and travel back up to the liver. The liver is responsible for reabsorbing almost all of the bile salts out of the hepatic portal vein and recycling them back into the bile. If the liver is functioning properly, only insignificant amounts of bile salts will bypass the liver and end up in the systemic circulation. This pathway for bile, from the liver to the intestine to the hepatic portal vein back to the liver, is called enterohepatic circulation.
The amount of bile salts that recirculates from the intestine to the liver influences how much bile is synthesized by the hepatocytes. As long as enterohepatic circulation of bile continues, bile synthesis will continue. When the stimulus for CCK secretion is gone, CCK secretion ceases and the sphincter of Oddi closes, diverting the bile back into the gallbladder. This means that the reabsorption of bile salts from the ileum back to the liver will diminish. Because the return of bile salts to the liver is the stimulus for continued bile acid production, this results in diminished bile acid synthesis.
In addition to their role in digestion and absorption of fats, bile acids are important in the excretion of organic compounds. Bilirubin, one of the breakdown products of hemoglobin, is an organic compound that is eliminated through the bile. Red blood cells, like all cells, have a finite life span and must be removed from circulation when that life span comes to an end. The majority of senescent (dead or dying) red blood cells are broken down through extravas- cular hemolysis (outside the vascular system). They are removed from circulation by the mononuclear phagocytic system. The mononuclear phagocytic system consists of macrophages found in the liver, spleen, bone marrow, lungs,
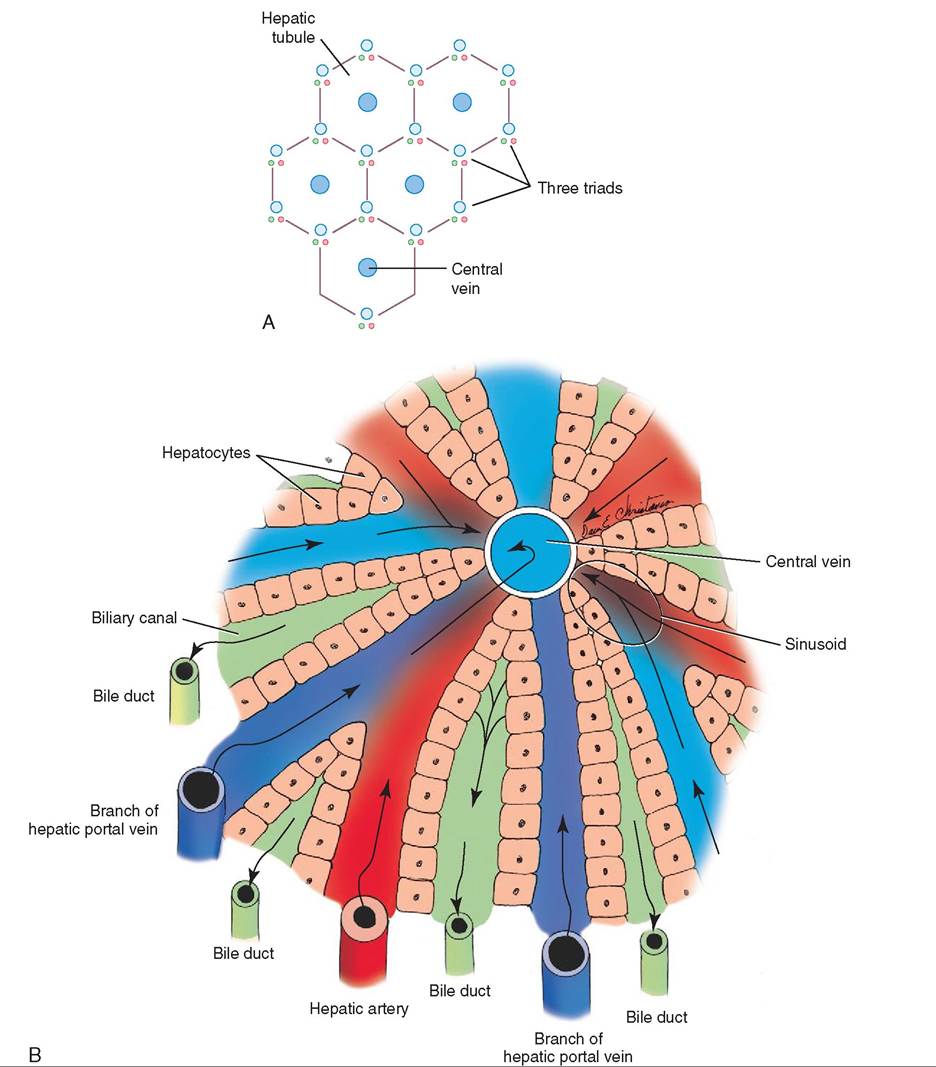
FIGURE 16-27 A, Anatomy of a hepatic lobe. B, Anatomy of a hepatic lobule. (B, From Christenson DE: Veterinary medical terminology, ed 2, St Louis, 2008, Saunders.)
and l^nph nodes. These macrophages phagocytize or engulf senescent red blood cells, rupturing them and releasing hemoglobin. This process is called hemolysis.
In the macrophage, hemoglobin is broken down into its component parts, globin and heme. Globin is degraded into amino acids that are reutilized by the animal. The iron is soemparated fr heme and is either stored in macrophages or transferred into plasma, where it combines with plasma proteins, becoming transferrin. Transferrin circulates to the bone mam>w and the iron is reused to make new hemoglobin.
Heme, which is a red pigment, is converted to biliverdin, a green pigment. Biliverdin is then converted into free or unconjugated bilirubin, ayellow to orange pigment. Uncon- iugated biIirnbin is released into the bloodstream, where it ictohmbines w the plasma protein albumin and travels to the liver. A this point the unconjugated bilirubin is not water soluble. The liver absorbs the unconjugated bilirubin. In the
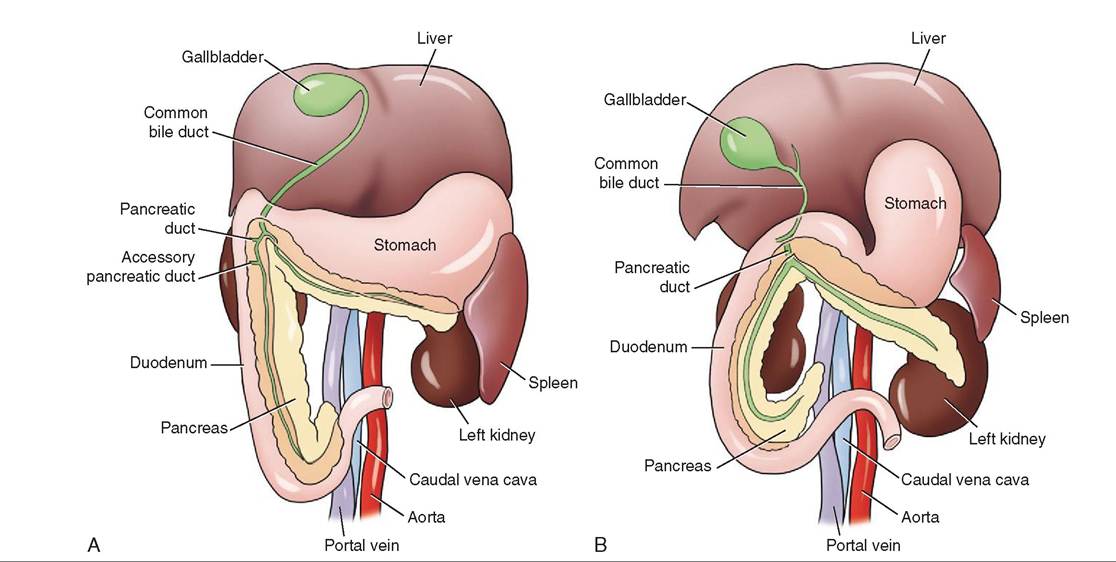
FIGURE 16-28 Schematic view of the bile duct and pancreatic duct emptying into the duodenum of A, the dog and
B, the cat. (From Washabau RJ, Day MJ: Canine & feline gastroenterology, St Louis, 2013, Saunders.)
hepatocyte, bilirubin is conjugated (joined) to glucuronic acid to form bilirubin glucuronide or conjugated bilirubin, which is water soluble. The hepatocyte releases the conjugated bilirubin into the bile, and when bile enters the small intestine conjugated bilirubin is transported along with it. In the small intestine, glucuronide is removed from conjugated bilirubin by bacterial enzymes. Fecal bacteria reduce the bilirubin to urobilinogen. Several different things can then happen to urobilinogen. Most urobilinogen is oxidized to urobilin and stercobilin and excreted from the body in feces. Urobilin is a yellow to orange pigment, depending on its degree of oxidation, whereas stercobilin is a brownish pigment. Together they give feces its characteristic color. The rest of the urobilinogen is reabsorbed into the enterohepatic circulation. From there it has two possible fates. Most of the urobilinogen is taken up by the liver and re-excreted into the bile, ultimately re-entering the enterohepatic circulation. Some of the urobilinogen in the hepatic portal vein bypasses the liver and enters the general circulation. The urobilinogen then travels to the kidney, where it is converted into urobilin and excreted from the body in urine, giving urine its characteristic color.
If this process is disrupted at any point, the level of bilirubin in the blood becomes elevated. As a result, the animal may become icteric (jaundiced), a condition in which the tissues turn a yellow or orange color. Disruptions can occur anywhere in the pathway, including before the liver (prehe- patic), within the liver (hepatic), and after the liver (posthe- patic) (Figure 16-29). If too many red blood cells are hemolyzed in the circulation, the amount of bilirubin can overwhelm the normally functioning liver and lead to prehepatic bilirubinemia. Bilirubinemia is an abnormal increase of bilirubin in the blood. If the liver is not functioning normally and the ability of hepatocytes to conjugated bilirubin is reduced, this can result in hepatic bilirubinemia. Cholestasis refers to a condition that prevents or slows the flow of bile through the bile ducts, causing a buildup of substances that would normally be excreted through the bile. If there is an obstruction of the bile duct (extraheptic cholestasis) or some impairment of bile flow within the canaliculi (intrahepatic cholestasis), this would results in posthepatic bilirubinemia.
TEST YOURSELF 16-10
1. What three structures are found in the triad located at the periphery of a hepatic lobule?
2. What is the function of Kupffer cells?
3. Where does the common bile duct enter the small intestine?
4. What common species of domestic animal does not have a gallbladder?
5. What is enterohepatic circulation?
6. Unconjugated bilirubin in blood is not water soluble. How does it become water soluble?
NUTRIENT PROCESSING IN THE LIVER
The liver plays a vital role in keeping blood glucose levels normal. In the nonruminant animal, carbohydrate digestion results in the production of monosaccharides, such as glucose, fructose, and galactose. Glucose is the primary monosaccharide absorbed from the small intestine, entering the hepatic
Systemic circulation
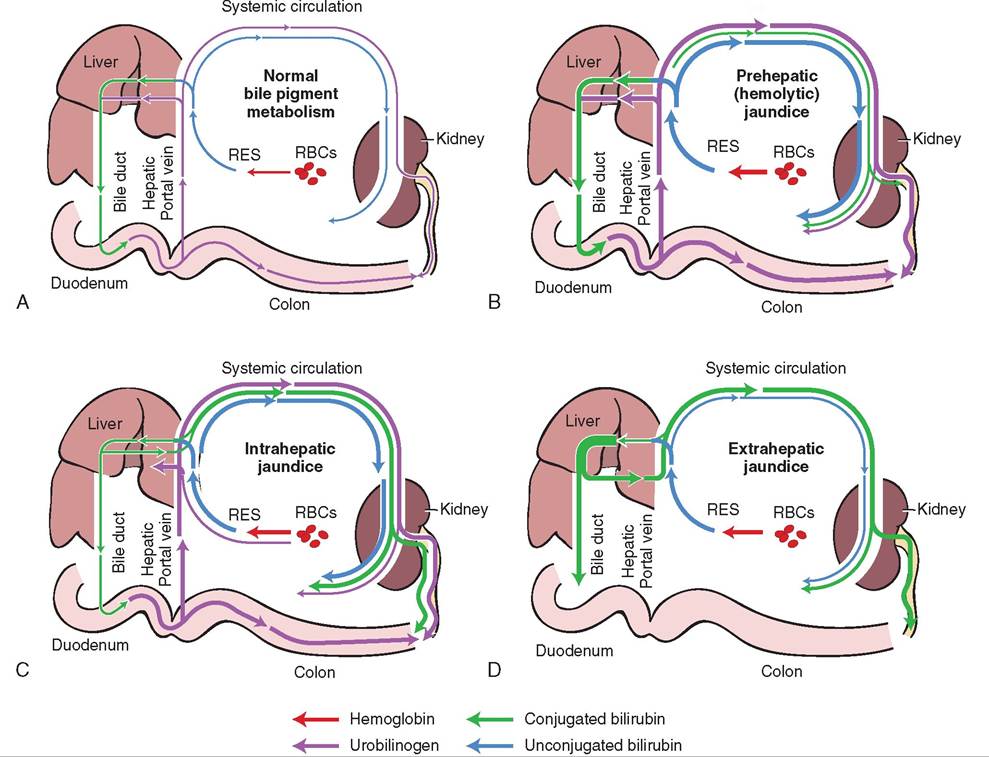
FIGURE 16-29 Diagrams comparing the formation, excretion, and enterohepatic circulation of bilirubin and other bile pigments. A, The normal animal; B, prehepatic jau∏cice associated with hemolysis and overproduction of bilirubin; C, intrahepatic jaundied associated with hepatocellular disease; D, posthepatic jaundied associated with extrahepatic bile buct oostruction. RBCs, dod bl∞d cells; RES, retusuk^(^r^do⅛l^lfcιl or mononuclear phagocytic system. (Courtesy Dr. Susan Johnson, The Ohio State University. In Washabau RJ, Day M: Canine & feline gastroenterology, St Louis, 201 3, Saunders.)
portal Wn and arriving at the liver. In the liver, glucose can be metebolized to produce energy in a number of forms, including ATIj. Conversion of glucose to energy occurs through the process of glycolysis. In addition to glucose, two other Hiondsaccharides that are absorbed in the nonrumi- uant (fructose and galactose) can be converted to glucose by the liver and, therefore, can be used as energy sources.
In the liver, as well as in skeletal muscle, if there is an excess amount of glucose, it can be stored as glycogen. Wfen blood glucose levels are high, some excess glucose can be tyaken up b adipose cells and converted to triglycerides. In tnhreunmoinant the liver can turn glucose into triglycerides when glucose levels remain high, even after glucose has been stored by the skeletal muscle and adipose tissue. In the rumi- rneant, the is less glucose available so this does not occur.
Glycogen, the major storage form of glucose, can be broken down to glucose monomers through a process called glycogenolysis wfen the animal needs to use glucose as an energy source. The process of glycogenolysis can occur when the animal is in the postabsorptive state (after absorption has occurred, between meals) and glucose is no longer so readily available.
During the postabsorptive state, when the availability of glucose is reduced, gluconeogenesis can also occur in the liver amnedtismoes the kidney. Gluconeogenesis is the synthesis loufcogse from noncarbohydrate sources, providing the animal with a source of glucose when glucose levels decline. The substrates used in this glucose-making process are pyruvate (formed from amino acids during protein breakdown), lactate, and glycerol.
When glucose is scarce, tissues can utilize lipids as an ceen.ergy sour Triglycerides in adipose tissue can be broken doown int glycerol and fatty acids by the enzyme lipase. Glycerol can then be converted into glucose in the liver by gluconeogenesis, whereas the fatty acids are oxidized in ivsasruieosus t as a source of energy. This process can be helpful Wn the animal's glucose level is low between meals. In eevere starvation, however, fatty acid mobilization from adipose tissue can increase tremendously, overwhelming the oxidative capability of the liver to convert fatty acids to energy. Instead the fatty acids are converted into ketones. Some tissues can use ketones as an alternative energy source, but too many ketones can ultimately cause a metabolic acidosis, called ketosis. It is important to note that, in nonruminants, ketones come only from the liver, as a consequence of the mobilization of fats, but in ruminants the ketone betahydroxybutyrate is also produced during normal digestion from butyrate, one of the VFAs.
Another function of the liver is protein production. The liver synthesizes nearly all the plasma proteins, including albumin and blood clotting proteins. Albumin is a blood protein that plays a vital role in keeping fluids in the vascular system. Because of its large size, albumin does not easily pass through most capillary walls. It exerts osmotic pressure, preventing water from moving out of the capillaries into the extravascular space. As a result, it helps to maintain the balance of fluids between the blood, interstitial spaces, and cells. If that balance is upset, abnormal fluid accumulations in body tissues can occur.
The liver also can convert amino acids into ketoacids. Ketoacids can be used by the liver for energy production, or they can be converted to glucose or fatty acids, which can be used in lipid synthesis. When amino acids are degraded in the liver, ammonia, a toxin, is formed. The hepatocytes convert ammonia to urea, which is later excreted primarily by the kidney in nonruminants. In ruminants, some urea re-enters the salivary glands and the rumen. Once it is in the rumen, microorganisms can use it again to synthesize proteins.
SMALL INTESTINAL MOTILITY
The two primary movements in the small intestine are peristalsis and segmentation. As described in the section on esophageal motility, peristalsis is a moving wave of luminal constriction preceded by an area of luminal distention (see Figure 16-13A). Peristalsis in the small intestine propels the intestinal contents toward the large intestine. Segmentation is a different type of movement carried out in the small intestine (see Figure 16-13B). The presence of chyme stretches the intestinal wall, triggering the ring of circular muscle to contract; as one set of contractions relaxes, another area of circular muscle contracts in a different area. These random, localized contractions of the circular muscle layer help mix the digestive contents and move the digestive fluid closer to the epithelial surface of the intestine, increasing its contact with the mucosal enzymes and absorptive surfaces. Segmentation is a mixing action only and does not move the chyme toward the large intestine.
REGULATION OF SMALL INTESTINAL MOTILITY
The pacemaker cells within the small intestinal wall are similar to those in the stomach and function as described in the section on gastric motility.
DIGESTION IN THE SMALL INTESTINE
CARBOHYDRATE DIGESTION
The small intestine is where the majority of carbohydrate chemical digestion occurs. Starches are one of the few carbohydrates broken down in the luminal phase. In luminal carbohydrate digestion, the enzyme alpha-amylase, which is secreted by the pancreas, breaks starch down into smaller chains of glucose molecules. These include the disaccharide maltose (two glucose molecules), trisaccharides (three glucose molecules), and oligosaccharides (three to ten glucose molecules).
The second phase of chemical digestion is the membranous phase. Embedded in the brush border are the specific enzymes for each type of polysaccharide. The polysaccharides must therefore come into contact with the intestinal epithelium to be broken down by the brush border enzymes. The enzymes are named according to the polysaccharides they break down. For example, the enzyme maltase breaks down the disaccharide maltose. The enzyme isomaltase breaks down the disaccharide isomaltose. Remember that starch is made up solely of repeating glucose units; luminal digestion of starch will yield only glucose molecules.
Sugars, such as sucrose and lactose, are not broken down in the luminal phase and instead are degraded in the membranous phase by the enzymes embedded in the brush border. The membranous enzyme sucrase breaks down sucrose into a glucose molecule and a fructose molecule, whereas lactase breaks down lactose into a glucose molecule and galactose molecule. Some adult mammals do not produce the enzyme lactase and therefore cannot digest lactose. Ultimately, enzymatic digestion of carbohydrates yields monosaccharides, which are small enough to be absorbed through the intestinal wall (Figure 16-30).
PROTEIN DIGESTION
Luminal protein digestion, which began in the stomach with pepsin, must be completed in the small intestine. The pancreas releases proenzymes that are activated in the lumen of the small intestine. Two types of enzyme participate in luminal digestion, the endopeptidases and the exopeptidases. Endopeptidases include pepsin as well as the pancreatic enzymes trypsin, chymotrypsin, and elastase. These enzymes break proteins at internal points along the polypeptide chains, resulting in smaller peptide chains. They do not release free amino acids. Exopeptidases, on the other hand, break proteins at the ends of the polypeptide chains, releasing free amino acids. Carboxypeptidases A and B are examples of exopeptidases. Protein luminal digestion by exopeptidases yields free amino acids.
The enzymes responsible for membranous protein digestion are also embedded in the brush border, and the peptides must come into contact with the epithelial surface for membranous digestion to occur. Peptide-digesting enzymes, or peptidases, hydrolyze the small peptide products of luminal phase protein digestion, yielding free amino acids. Some
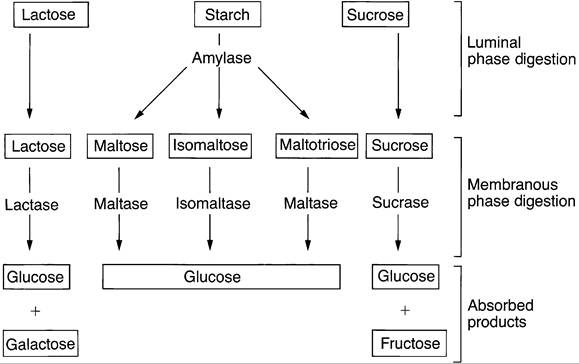
FIGURE 16-30 Carbohydrate digestion. (From Cunningham JG: Textbook of veterinary physiology, ed 4, St Louis, 2007, Saunders.)
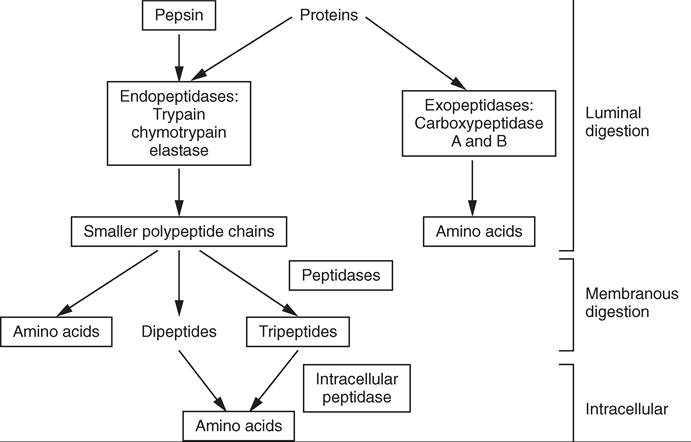
FIGURE 16-31 Protein digestion. (From Cunningham JG: Textbook of veterinary physiology, ed 4, St Louis, 2007, Saunders.)
peptide chains are incompletely hydrolyzed and form dipeptides and tripeptides. These dipeptides and tripeptides are small enough to be absorbed into enterocytes (intestinal wall cells). Within the enterocytes are intracellular peptidases that complete hydrolysis, producing free amino acids that are available for passage into the blood. Only free amino acids are small enough to be absorbed into the blood (Figure 16-31).
ABSORPTION OF MONOSACCHARIDES, DIPEPTIDES, TRIPEPTIDES, AND
AMINO ACIDS
Absorption across the intestinal mucosa can occur in a multitude of ways, including both passive and active processes. Although some molecules are transported passively by diffusion down their concentration gradients, others must use an active transport process, using energy to enter the body. Some substances rely on multiple methods. For example, electrolytes are absorbed by attaching to one of several transport molecules and use a variety of mechanisms responsible for completing their absorption. Other molecules are transported by antiports that allow one molecule to be exchanged for another, such as the sodium/hydrogen exchange. Glucose and amino acids often have to rely on secondary active transport for absorption. Glucose relies on the establishment of a sodium gradient to help move it into the enterocyte against its concentration gradient. When glucose levels are high within the enterocyte and lower in the lumen of the GI tract, an unfavorable concentration gradient is formed preventing glucose from passively entering into the enterocyte. The body cannot allow this glucose to be lost in the feces. A sodium-potassium pump found on the enterocyte near the blood side of the cell pumps three sodium atoms out and two potassium atoms in, helping to create a lower concentration of sodium inside the cell. This creates a concentration gradient that favors the movement of sodium from the lumen of the gastrointestinal tract into the enterocyte. As the sodium is moving into the cell, glucose moves by cotransport into the cell along with the sodium, against the glucose concentration gradient. This is an example of secondary active transport and both glucose and amino acids can be brought into the enterocyte through this process. To complete the process, glucose must then move through the cell wall into interstitial space by facilitated diffusion. Facilitated diffusion is a passive transport process, meaning it does not require ATP, but uses a carrier molecule to move glucose out of the cell. The glucose completes its absorption process by entering the capillaries from the lateral space by simple diffusion.
LIPID DIGESTION AND ABSORPTION
Fats and fat-soluble vitamins are also digested in and absorbed from the small intestine. Triglycerides are the primary dietary lipids; other lipids include cholesterols, phospholipids, and esters. Lipids are not water soluble and cannot dissolve in the watery medium of the gastrointestinal fluids. In order to digest and absorb fats, a four-step process must occur, comprising emulsification, hydrolysis, micelle formation, and absorption.
The first step, emulsification, begins in the stomach, where lipids are warmed to body temperature and thoroughly mixed. This process helps break lipids into smaller droplets. Emulsification is then completed in the small intestine by the detergent action of bile salts and phospholipids. If bile salts were not available, the smaller fat droplets would coalesce back into larger fat droplets in the intestine, which does not have the intense mixing action of the stomach. Bile salts coat the lipid droplets, causing them to repel one another and remain as small individual droplets. Decreasing the size of lipid droplets increases the surface area available for the hydrolytic pancreatic enzymes, lipase and colipase, to break down each triglyceride into a monoglyceride and two fatty acids. The monoglyceride and the two fatty acids are not yet soluble in water, which again would cause a problem if it were not for the bile salts forming micelles. The monoglycerides and fatty acids form a union with the bile salts and phospholipids in a mass, where the water-soluble bile salts and phospholipids are on the outside and the waterinsoluble monoglyceride and fatty acids are on the inside, completely surrounded by the bile salts and phospholipids. It looks as though the entire molecule is water-soluble, not just the outside surface. The micelles essentially act like tiny ferries, transporting the fatty acids and monoglyceride to the enterocyte surface. At this point the monoglyceride and fatty acids diffuse into the cell and the bile salts do not. This keeps the bile salts available to transfer other monogylcerides and fatty acids. Inside the enterocyte, the monoglyceride and the two fatty acids are reassembled into a triglyceride and packaged into chylomicrons. Chylomicrons are spherical structures that are made up of triglycerides, cholesterol, phospholipids, and proteins. The Golgi apparatus packages the chylomicrons into vesicles, which are transported to the cell membrane, from which they are released into the interstitial fluid by exocytosis. Chylomicrons cannot enter the capillaries because of their large size; instead they are picked up by the lymphatic system and transported to the systemic circulation without passing through the liver.
TEST YOURSELF 16-11
1. The major storage form of glucose is.
2. How are triglycerides in adipose tissue converted into glucose for energy?
3. Which plasma protein produced in the liver plays a crucial role in fluid movement between plasma and interstitial fluid?
4. Which movement in the small intestine propels intestinal content toward the rectum?
5. In the luminal phase of the chemical digestion of starches, which enzyme, secreted by the pancreas, is needed?
6. In the membranous phase of chemical digestion of sugars, where are the necessary enzymes located?
7. What is an exopeptidase?
8. Both glucose and amino acids are brought into a cell through secondary active transport? What does this mean?
THE LARGE INTESTINE
BASIC STRUCTURE AND FUNCTION
The end of the small intestine, the ileum, leads into the large intestine, which consists of the cecum, colon, rectum, and anus. The large intestine overall has a wider diameter then the small intestine, hence the name. The amount of microbial fermentation that occurs within the large intestine contributes to how extensive and complex its anatomy is. Whether the ileum opens into the colon, cecum, or both depends on the species.
The cecum is a blind diverticulum at the beginning of the colon. It varies from being small and inconspicuous in the carnivore, where little fermentation is required, to a voluminous and expansive cecum in equine species that rely heavily on hindgut microbial fermentation. In the horse, the comma- shaped cecum is so expansive that it takes up a large portion of the abdomen on the right side of the animal (Figure 16-32). It consists of a base, the main body, and an apex (point of the blind diverticulum). The cecum in the ruminant is a large blind tube that extends caudally.
The colon can be divided into three parts, ascending, transverse and descending, with the most variation between species appearing in the ascending colon (Figure 16-33). In the carnivore, the ascending colon is short and straight,
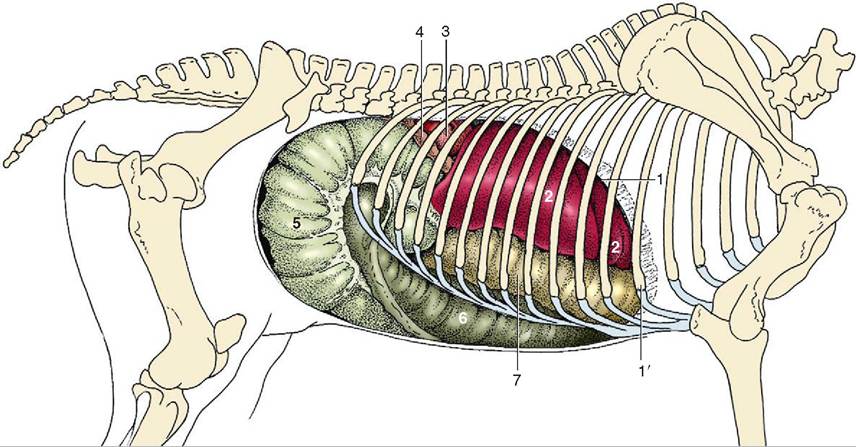
FIGURE 16-32 Visceral projections on the right abdominal wall (including the diaphragm) of a horse. 1, Cut edge of the diaphragm; 1', rib #6; 2, liver; 3, right kidney; 4, descending duodenum; 5, body of cecum; 6, right ventral colon; 7, right dorsal colon. (From Dyce KM, Sack WO, Wensing CJG: Textbook of veterinary anatomy, ed 4, St Louis, 2010, Saunders, VitalBook file.)
extending cranially and leading into the transverse colon. The transverse colon goes from the right side of the abdomen to the left, often running between the stomach and the small intestine, and terminates at the longest part of tohloen,c the descending colon. The descending colon follows the left side of the abdomen before going medially teocttuhme.r
oTlhoen c in the pig and ruminant consists of the same
three parts. However, the ascending colon forms a coil or spiral, which is quite different from the carnivore's short and straight ascending colon. The ruminant's ascending colon in feact can b further subdivided into a proximal loop, forming an S shape; a spiral loop, which coils toward the center of the mesentery and then coils back up in the reverse direction; and a distal loop, which connects the ascending with the transverse colon. In the pig, the ascending colon also forms a spiral arrangement but instead of being in a flat plane, it iesdshap more like a cone. In both animals, the ascending cnolon the leads into the transverse colon, which crosses ifogrohmt r t left, similar to that of the carnivore, leading iensctoentdhiendg colon.
rTshee ho has the largest and most complicated colon of the domestic species and it is often referred to as the great choelon. T horse's ascending colon actually forms a double horseshoe-shaped loop, with one loop (the dorsal colon) lying on top of the other (the ventral colon). The bottom of the horseshoe is toward the front of the animal and the turn between the two occurs at the pelvic inlet. The colon narrows significantly at the transition from ventral to dorsal colon, raenfdo rtehe transport can be delayed in this area.
During segmentation in the colon and cecum of the horse,aonthd pig b the longitudinal muscle, which runs in bands
called teniae, and the circular muscle contract, forming bulging saclike structures called sacculations. Tfiese saccula- tions are also called haustra. Certain areas contract then rnelax, the different areas contract and relax in another loca- tchiceounla. tTionssa are semipermanent and may even be
voisstible p mortem. Sacculations prolong the time during ownhtiecnhtsthe c stay in the large intestine by creating eoxlutrma ev. This allows more time for absorption and
microbial digestion.
MOTILITY OF THE LARGE INTESTINE
eTshtienainl t contents (ingesta) travel slowly in the large
intestine, especially in animals such as the horse and the reaqbubiriet that r extensive hindgut fermentation to provide ennergy. I the large intestine, the ingesta must be thoroughly omixed t allow contact with the absorptive surface of tohloenc, where absorption of water and ions occurs. Dif- fyepresnt t of motility pattern occur within the large intes-
ittihne w the function of mixing, retaining, and propelling ingesta toward the anus. Although they vary among species, itlhitey mot patterns in general consist of four different types: seamentation, peristalsis, antiperistalsis, and mass movement.
Segmentation is the most common type of movement in tehceumc and colon. Segmental contractions move the ingesta in a slow back and forth movement, increasing the eontact between the ingesta and the epithelial surface. Peristalsis in the large intestine is similar to peristalsis in other parts of the GI tract, moving ingesta toward the anus.
Antiperistalsis occurs in the opposite direction, toward the stomach. The movement helps slow down the transit
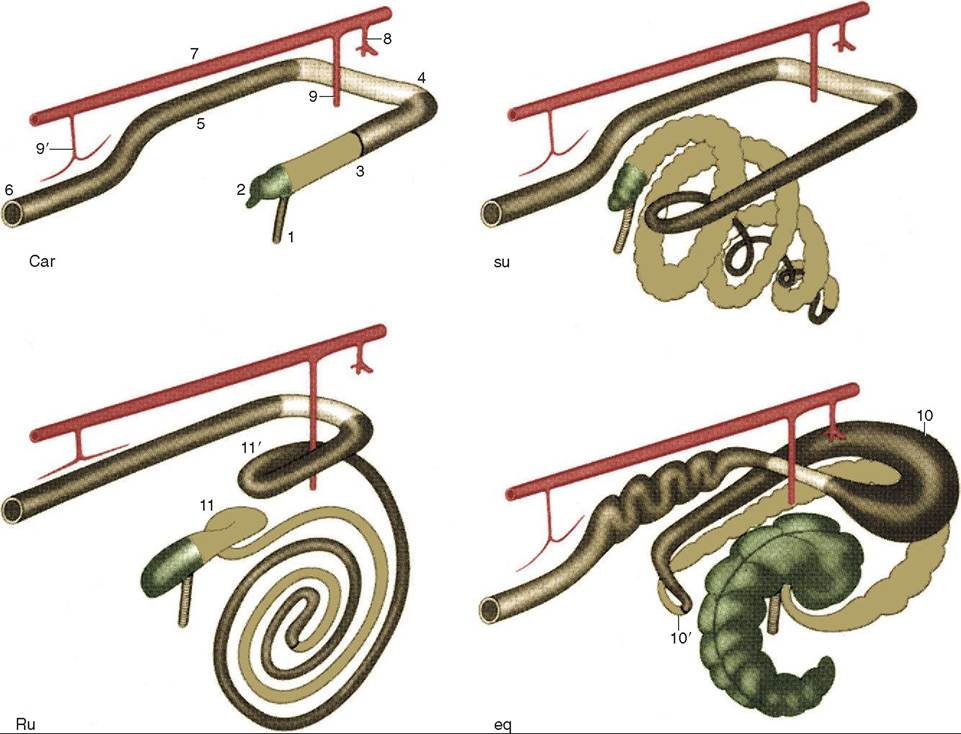
FIGURE 16-33 Schematic drawing of the large intestine of the domestic mammals: carnivores (Car), the pig (su), ruminants (Ru), and the horse (eq). Cranial is to the upper right. 1, Ileum; 2, cecum; 3, ascending colon; 4, transverse colon; 5, descending colon; 6, rectum and anus; 7, aorta; 8, celiac artery; 9, 9', cranial and caudal mesenteric arteries; 10, 10', dorsal diaphragmatic and pelvic flexures of ascending colon; 11, 11', proximal and distal loops of ascending colon. (From Dyce KM, Sack WO, Wenseng CJG: Textbook of veterinary anatomy, ed 4, St Louis, 2010, Saunders.)
of the ingesta, allowing more time for reabsorption to occur. The hairpin turn between the ventral colon and the dorsal colon is narrow, and functions in a very similar way to the pylorus of the stomach. In the horse, this area is often where the antiperistalsis movements originate. Antiperistalsis in this area also prevents larger food molecules that rhquire further digestion from passing through, allowing rmaller, more liquid, ingesta to continue its journey into the odloornsa. l c
The last type of movement seen in the large intestine is called mass movement. It consists of a period of intense propulsive a ct ivi ty, moving the contents of the large intestine toward the rectum. Mass movement contractions occur owtihthin b the colon and the cecum.
REGULATION OF LARGE
INTESTINE MOTILITY
alow waves are initiated by large intestinal pacemaker cells that have the same fluctuating membrane potential as seen in aortthser p of the gastrointestinal tract. The pacemaker cells can be found throughout the large intestine; they have alternating periods of activity and rest, allowing different hreas of the large intestine to be the active pacemakers. Depolarizing waves from the various pacemaker cells can be sent thoeward t stomach, creating antiperistaltic movements, or thoeward t rectum, resulting in peristaltic movements. The autonomic nervous system influences the activity of the pacemaker cells in the large intestine, just as in the small intestine.
DIGESTION AND ABSORPTION
IN THE LARGE INTESTINE
eTrwyo v important functions of the large intestine are absorption of water and ions, and completion of carbohy- rdortaetien o p microbial digestion and absorption. The forestomachs in the ruminant are the primary location of microbial digestion, whereas in nonruminant hindgut fer- rmbeivnotirnesg, he such as horses and rabbits, microbial digestion occurs mainly in the cecum and colon. Although the physical location of fermentation is different, the substrates being broken down in the cecum are similar to those fermented in the rumen. These substrates are mainly the structural and nonstructural carbohydrates found in plants, aemlslew as so proteins.
The horse’s fermentation center, being positioned after the stomach where enzymatic digestion occurs, influences how food is processed in the cecum and colon. Carbohydrates are exposed to enzymes in the small intestine, making them more susceptible to breakdown by microbes in the oecum and colon. Some simple carbohydrates may be broken ydown b the enzymes and acids in the stomach, but in comparison to the carnivore, the enzymatic digestion process that occurs in the horse is far less efficient. Additionally, the ohnigchenctration of cell wall or structural elements found ionodplant f may act to protect the simple nonstructural components from exposure to digestive enzyme, thus pre- ivrenting the degradation.
Soimilar t the ruminant, the horse, through the process of fermentation, produces VFAs that are absorbed from the cecum and colon and used as an energy source. There also are local glands in the wall of the large intestine that produce bicarbonate that helps to neutralize acids produced during tehrme fentation process.
Proteins from the food the microbes would have used for trhoewirthg and metabolism have already been broken down and absorbed by the horse during enzymatic digestion, oprior t the ingesta entering the cecum and colon. This presents the microbes in the cecum and colon with a potential enfiitcrioegnecny.d To solve this problem, extensive amounts
of urea are transferred from the blood to the large intestine.
The microbes use the urea, along with the small amount of protein that has escaped enzymatic digestion, to make amino raocitdeisnasn. d p
EMPTYING OF THE RECTUM
When ⅛τιe (ingesta) passes through the large intestine fmuch o the water is absorbed, leaving a semisolid material ceacellse.d f The consistency of normal feces varies among species from semiliquid “cow pies” to small, hard sheep pellets. ^^en feces are Vmedorted to the rectum sensory receptors are stimulated. This initiates the defecation reflex that causes the colon and rectum to contract, and relaxes the inner anal sphincter muscle, a smooth muscle. The animal tehrceenivpes the need to defecate. Some species such as the cat and the dog can be trained to close the voluntary outer anal sphincter, a ^reletal muscle, and delay defecation. Other aunchimals s as ruminants, horses, and birds seem unable to lowearn ho t control this sphincter voluntarily.
TEST YOURSELF 16-12
1. List thefourpαrts ofthe Iargeintestine.
2. Which species of lotgr onimol is o hindgut fhtmrntht?
o. Namea pommoo sancies ohanloaI hhathas a short ascending colon?
4. Nomea eemmon Sfncies cCaniaιal hhet fas a spiral oscending colon sloped like o cone?
5. W hot isthe purpossoftSesacculations?
6. What ate tnefohrmovemested ssoci atedwite the large intestine?
7. HPowafo feeds formed?