Glycolysis
We begin our quest to understand energy production with glucose because it is typically the most common nutrient used for acute energy production. The initial processing of glucose molecules begins in the cytoplasm of the cell.
Once inside the cell, 6-carbon glucose is converted in a series of reactions into two 3-carbon molecules of pyruvate. The 10 linked reactions responsible for this conversion are collectively called glycolysis, the Embden-Meyerhof pathway, or anaerobic respiration. It is typical to describe carbohydrate catabolism beginning with glucose, but you should remember that polymers of glucose (glycogen in animal cells) can be cleaved to provide the glucose monomers to enter the glycolysis pathway. Thus, the cell may well contain stores of glycogen that can be cleaved to supply glucose aside from the uptake of glucose across the plasma membrane. Table 3.2 provides a list of some often confusing, similar-sounding terms that have to do with carbohydrate metabolism. It may be useful for you to periodically refer to this table as you study this important topic. Figure 3.3 provides an outline of each of the steps in glycolysis. We have purposefully not given all of the chemical structural detail showing changes in carbons and functional groups as glucose is modified through glycolysis. If this is needed, it is available in any introductory biochemistry book. Our goal is for you to appreciate the highlights, the overall chemical events, and most importantly, the physiological relevance. Although each of the reactions of glycolysis is enzyme- mediated, we have focused on two glycolysis reactions and their associated enzymes along with one other reaction that is not strictly part of glycolysis. These are (1) the conversion of glucose to glucose-6- phosphate (step 1 of glycolysis, catalyzed by hexoki- nase), (2) the conversion of fructose-6-phosphate into fructose 1,6-diphosphate (step 3, catalyzed by phosphofructokinase [PFK]), and (3) the conversion of pyruvate into lactate (catalyzed by lactate dehydrogenase). Some specific reactions and molecules associated with glycolysis are provided in Figure 3.4. At this point, it is worth remembering the significance of glycolysis. This pathway allows the conversion of the 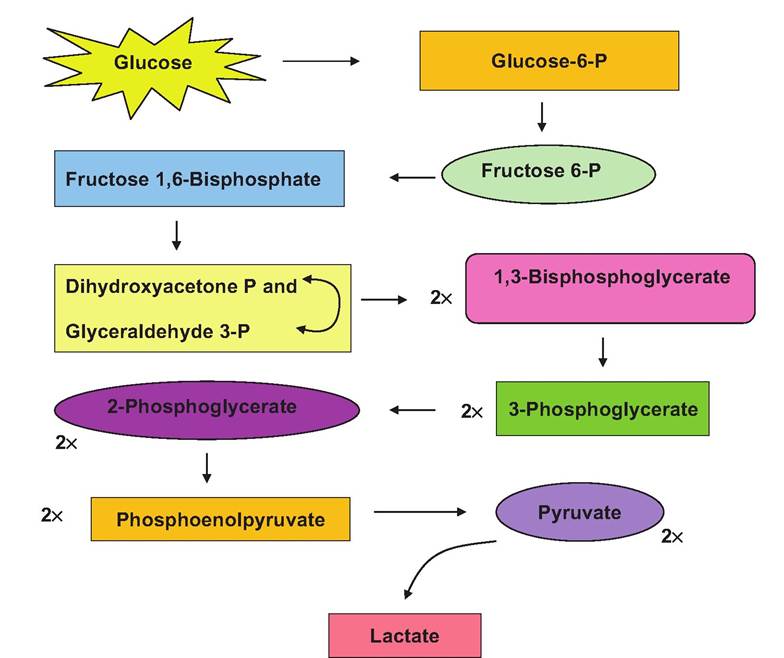
Fig. 3.3. Chemical steps and intermediates in glycolysis are illustrated.
nutrient glucose into molecules that can then be shuttled into the mitochondria for use in the process of oxidative phosphorylation. However, as we indicated earlier, oxidative phosphorylation (simply the production of ATP linked to a series of oxidationreduction reactions) requires oxygen. In addition to preparing molecules for entrance into the mitochondria, a small amount of ATP is produced during the glycolysis reactions. In contrast with mitochondrial activity, this occurs via substrate-level phosphorylation. As you will see, the amount of ATP made in this manner is very small compared with that which occurs with the complete catabolism of the glucose (glycolysis reactions + mitochondrial activity), but it is nonetheless essential. This is because production of ATP via glycolysis alone can occur in the absence of oxygen. For this reason, it is called anaerobic respiration.
Let us outline the steps of glycolysis. Typically, glucose is captured by the action of membrane transporters and passed into the cytoplasm. At this juncture, a phosphate group is added to the sixth carbon of the glucose. This then produces glucose-6-phos- phate. Somewhat ironically, even though the glycolysis ultimately leads to energy production, in the first step of glycolysis, ATP is actually used. This is because the phosphate group added to the glucose is donated from ATP as illustrated in Figure 3.4. In step 2, glucose- 6-phosphate is converted into another hexose sugar by the action of the enzyme phosphohexose isomerase, that is, fructose-6-phosphate. Step 3 again utilizes another molecule of ATP as the enzyme PFK catalyzes the addition of another phosphate group to produce fructose 1,6-diphosphate.
Step 4 is a cleavage reaction catalyzed by the enzyme aldolase, which produces two 3-carbon molecules. These are dihydroxyacetone phosphate and glyceraldehyde 3-phosphate. In step 5, the enzyme phosphotriose isomerase converts dihydroxyacetone into a second molecule of glyceraldehyde 3-phospate. There are now two identical molecules to continue through glycolysis so that products made from this point are doubled. Step 6 depends on the enzyme glyceraldehyde 3-phosphate dehydrogenase. As we will discuss in more detail relative to mitochondrial activity, this enzyme depends on the oxidized form of the coenzyme nicotinamide adenine dinucleotide or NAD+. During this reaction the NAD+ becomes reduced (NADH), and inorganic phosphate is added to the substrate to produce 1,3-bisphosphate glycerate. The first direct production of ATP occurs in step 7 as a phosphate group is cleaved, and the energy is utilized to simultaneously add a phosphate group to ADP This is an example of substrate phosphorylation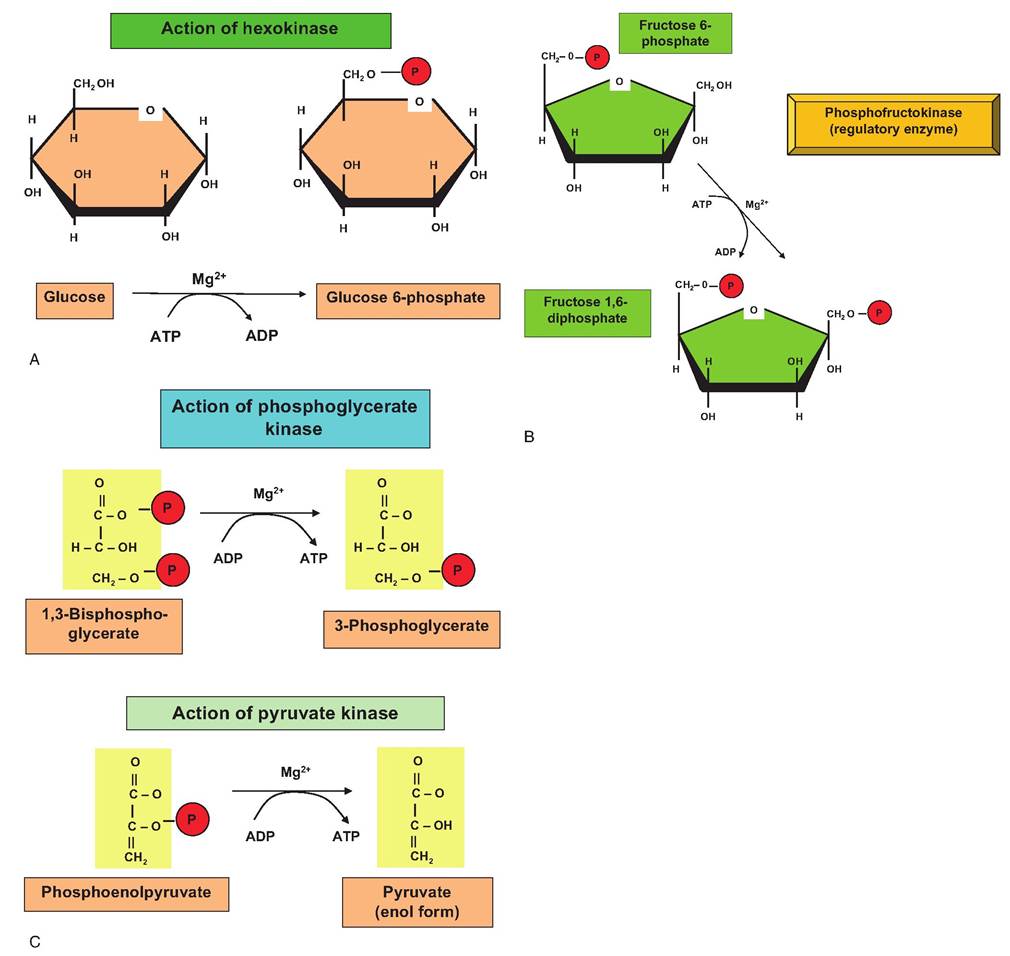
Fig. 3.4. Selected reactions of glycolysis. Panel A shows the action of the hexokinase. This enzyme adds a phosphate group (red circle) to carbon 6 of glucose. This effectively traps the glucose inside the cell since it can no longer bind to membrane transporters. Panel B shows phosphorylation of fructose 6-phosphate by the enzyme phosphofructokinase (PFK). PFK is a critical regulatory enzyme of glycolysis because its activity is modulated by the ATP/ADP ratio in the cytoplasm. A high ratio inhibits and a low ratio stimulates the enzyme. These first two reactions can be thought of as energy requiring activation phases of glycolysis. Panels C and D illustrate two reactions that produce ATP by substrate phosphorylation. Remember, two molecules of each substrate are produced when fructose 1,6-diphosphate is cleaved (steps 4 and 5 of glycolysis) so a total of four molecules of ATP are produced.
When the ATP debt is paid for use of ATP in the first two reactions, the net gain is +2 ATP under anaerobic conditions. If the reduced NADH (two molecules) produced in step 6 of glycolysis can be oxidized in the mitochondria (aerobic conditions), an additional six ATP are generated as a consequence of glycolysis.to produce a molecule of ATP, and the reaction is catalyzed by phosphoglycerate kinase. Step 8 depends on phosphoglycerate mutase to induce a rearrangement of the 3-phosphoglycerate made previously to yield 2-phosphoglycerate. Step 9 is the conversion of the 2-phosphoglycerate into phosphoenolpyruvate catalyzed by enolase. Step 10 is another ATP-making event as the enzyme pyruvate kinase acts to transfer a phosphate from phosphoenolpyruvate to ADP as pyruvate is also created. At this point, the pyruvate is at a crossroads. The redox state of the tissue determines which of two alternative paths will be followed. If oxygen is available, the pyruvate is shuttled into the mitochondria as subsequently described. If, however, oxygen is
limited, that is, there are anaerobic conditions, pyruvate is reduced by the action of lactate dehydrogenase, and the coenzyme NADH becomes oxidized again. This is a critical process under these conditions. You may recall that step 6 of glycolysis requires the oxidized form of this coenzyme (NAD+). Although only a small amount of ATP is derived directly from glycolysis, even this would be lost were it not for the action of lactate dehydrogenase. As an aside, there are also several isozymes of lactate dehydrogenase that are important clinically. For example, the unique structure of the LDH from heart muscle can be detected in blood serum in animals (or people) that have suffered cardiac injury (Fig. 3.5). Table 3.3 summarizes ATP production that is associated with glycolysis.
The Cori cycle
Most carbohydrates in the diet can be readily converted into glucose, galactose, or fructose when digested.
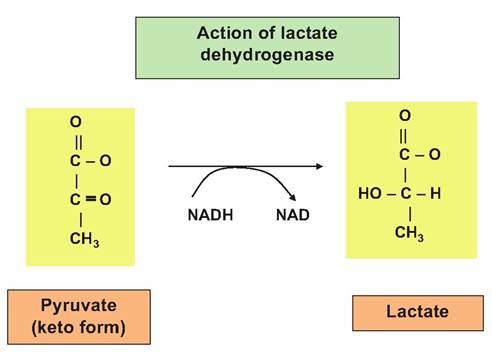
Fig.
3.5. The action of lactate dehydrogenase (LDH) is illustrated. After an initial spontaneous rearrangement of pyruvate from the enol form to the keto isomer, LDH catalyzes the conversion of pyruvate to lactate. Most important in anaerobic conditions, this is linked with the simultaneous oxidation of reduced NADH. This newly produced NAD is essential for glycolysis to continue under these conditions. This is because NAD+ is required for step 6 of glycolysis.These molecules are absorbed into the portal vein that drains the intestinal tract for use by the liver and other tissues. There are various other compounds that are considered glucogenic. These are molecules that can readily be converted into glucose to be processed via glycolysis for subsequent ATP production or for use in other biochemical pathways. For example, propionate that is derived from fermentation of dietary carbohydrates in ruminant animals is an essential substrate to allow ruminants to synthesize the glucose they need. These glucogenic compounds can be divided into two groups: (1) those that are essentially direct conversions into glucose without a significant amount of recycling, for example, propionate and certain amino acids, and (2) products of partial metabolism of glucose in particular tissues that are then transported to the liver or kidney for generation of glucose, for example, lactate. In all animals, there are times when oxygen is locally limited so that anaerobic respiration is favored and lactic acid accumulates. For example, the accumulation of lactic acid occurs in active muscle tissue and produces the sensation of muscle fatigue. In addition, since erythrocytes lack mitochondria, they rely on glycolysis for all of their ATP needs and consequently continually produce lactic acid regardless of oxygen availability. Most of the lactic acid from muscle or erythrocytes diffuses into the bloodstream. Fortunately, it is transported to the liver and, to a lesser extent, the kidney where it can be converted into glucose. At this point, it can be stored as hepatic or renal cell glycogen or released back into the bloodstream for use by muscle or other tissues (see Fig. 3.10). Cycling of lactic acid from muscle (or other tissues) to liver and the return of glucose is called the Cori cycle and is outlined in Figure 3.6.