The Krebs cycle
Let us now trace the fate of pyruvate that is produced at the end of glycolysis under aerobic conditions. Remember our goal here is to understand how the catabolism of glucose and other carbohydrates is used to generate the ATP that is essential to cells and tissues.
Table 3.3. Summary of ATP production in glycolysis.
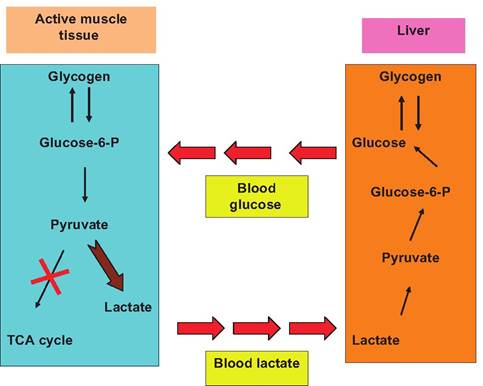
Fig. 3.6. The Cori cycle. In active muscle (e.g., with anaerobic conditions) the conversion of pyruvate into acetyl-CoA and processing via the TCA cycle is blocked (red X). This leads to lactate or lactic acid production. The lactate diffuses out of the muscle tissue into blood where the liver and kidney can convert lactate into glucose-6-phosphate for storage as glycogen or conversion to glucose-1-phosphate then glucose. Muscle or other tissues in the body can then use this regenerated glucose.
Once this foundation is established, we will then be able to understand how other nutrients can also be used for energy production. The next major biochemical pathway for this processing is called the citric acid or Krebs cycle. These reactions occur inside the mitochondria, so before pyruvate can be modified, it has to pass across the mitochondrial membrane.
This occurs via the action of a specific membrane transporter. The pyruvate is then quickly oxidatively decarboxylated (removal of CO2) to produce acetylcoenzyme A (acetyl-CoA). This reaction is catalyzed by the action of the multienzyme complex pyruvate dehydrogenase. The product, acetyl-CoA plays an especially central role in energy metabolism. This overall reaction involves two coenzymes:
Pyruvate + NAD+ + CoA
→ Acetyl-CoA + NADH + CO2.
The oxidized-reduced NAD+ and NADH are familiar from the action of glyceraldehyde 3-phosphate dehydrogenase or lactate dehydrogenase and reactions of glycolysis. Now a description of CoA is in order. CoA is a complex molecule derived from pantothenic acid (common in meats and grains), thioethanolamine, and ATP. The essential feature is that CoA acts as a carrier of acyl groups. Specifically, the thiol group of the thioethanolamine residue of the molecule functions in this manner in a variety of reactions involved in fatty acid oxidation and fatty acid synthesis and acetylation reactions. The molecule is also important in oxidative decarboxylation reactions as with pyruvate. Acommon convention is to abbreviate the structure of the reduced form of the molecule as CoA∙SH, which designates the reactive SH group of the molecule. The acetyl group that is now part of acetyl-CoA is derived from the catabolism of pyruvate (two carbons remaining after decarboxylation of pyruvate). As we will see, acetyl- CoA is at the confluence of a variety of major metabolic pathways. Almost all of the many specific carbohydrates and fats that are Catabolized for energy production are utilized to generate acetyl-CoA. In addition, a number of the nonessential amino acids from degraded proteins also are cannibalized to generate acetyl-CoA. As a special case in ruminants, one of the major products from fermentation of dietary carbohydrates is acetate, which is readily converted to acetyl-CoA for subsequent processing through the mitochondria.
Back to our story: at this point, the two carbons of the acetyl group of acetyl-CoA and the 4-carbon molecule Oxaloacetate condense to create the 6-carbon compound citrate. This is the first step of the Krebs cycle as outlined in Figure 3.7. There are two critical physiological points to the Krebs cycle reactions. The first is that some ATP is produced directly via substrate-level phosphorylation of ADP similar to that which occurs in glycolysis.
The second and most important is that with each turn of the cycle, reduced forms of the coenzymes NAD and flavin adenine dinucleotide (FAD) are produced. When enzymes of the electron transport chain, also located in the mitochondria, subsequently oxidize these molecules, this yields the energy for synthesis of the vast majority of ATP that can be created from overall catabolism of glucose. We have provided only a skeleton outline showing the names of the intermediates of the Krebs cycle reactions and locations of specific events. Remember that each molecule of glucose generates two molecules of pyruvate so there are two turns of the cycle for each glucose molecule.As was the case with glycolysis, the coupled oxidation-reduction reactions are a critical part of the processing. The combination NAD+ and NADH appears again along with FAD and FADH. The enzymes that catalyze these oxidation reactions by removal of hydrogen atoms are dehydrogenases, for example, lactate dehydrogenase whose action is illustrated in Figure 3.9. For these reactions to take place, the enzymes require the assistance of coenzymes that act to hold or carry these hydrogen atoms. It can be a source of confusion, but transfer of the hydrogen atom, with its lack of a neutron but paired electron and proton, is effectively viewed as an electron transfer. Thus, oxidation-reduction reactions defined by either electron acceptance or electron donation are often linked with movement of the hydrogen atom. This explains the abbreviations related to FAD versus FADH2 or
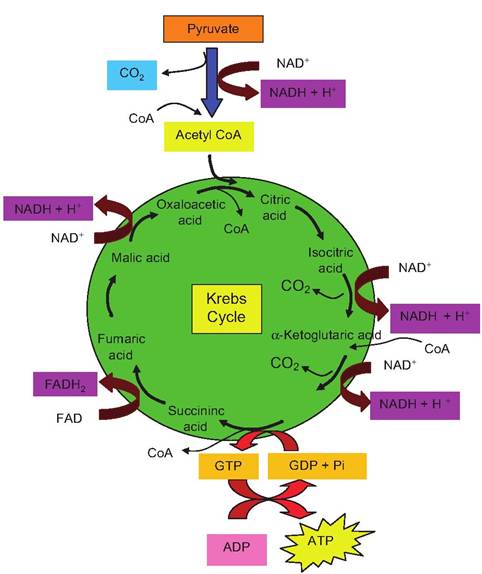
Fig. 3.7. Outline of the Krebs cycle reactions. Since each glucose molecule yields two pyruvate molecules (in the presence of oxygen), this allows two turns of the cycle. With each turn, two carbons are removed from the citric acid (six carbons) by decarboxylation reactions; this leads to the production of the 4-carbon intermediate oxaloacetic acid.
Oxaloacetic acid initiates the cycle as it condenses with acetyl-CoA (two carbons) to produce citric acid. Although the pyruvate that entered the mitochondria had three carbons, remember that two carbons were lost as CO2 to generate acetyl-CoA. Although not "officially" part of the Krebs cycle, at the time of decarboxylation of the pyruvate, NAD+ is simultaneously reduced. Four additional oxidations by the removal of hydrogen atoms occur during the cycle. This yields four molecules of reduced coenzymes (three NADH + H+ and one FADH2). One ATP molecule is made with each turn of the cycle due to the initial creation of GTP, which then provides the phosphate group to make ATP from ADP. As in glycolysis, this is another example of substrate-level phosphorylation.NAD+ versus NADH + H+ in the Krebs cycle reactions outlined in Figure 3.7. Figure 3.8 gives an example of this type of reaction.
At this point you are probably wondering just how much ATP is generated from glucose catabolism and when this actually occurs. As we have seen, each turn of the Krebs cycle only generates two molecules of ATP via substrate-level phosphorylation. The key depends on a cluster of interrelated membrane-bound enzymes that make up the electron transport chain. The activity of these enzymes also accounts for essentially all of our need for oxygen. As the electron chain enzymes function, the hydrogen atoms (electrons) that are removed as various intermediates of glycolysis and Krebs cycle are oxidized and are progressively
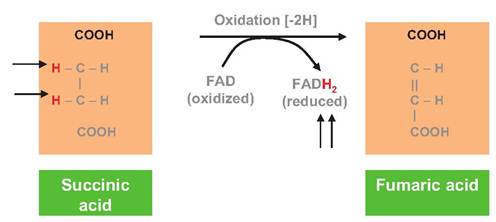
Fig. 3.8. Reduction of FAD. Coupled reduction of FAD in conjunction with the conversion of succinic acid to fumaric acid as occurs in the Krebs cycle is shown. Arrows indicate the fate of hydrogen atoms (electrons).
passed along until they are combined with oxygen. Oxygen is the final electron acceptor in the chain so that water is formed.
The reduced forms of both NADH and FADH2 that were generated in the Krebs cycle become oxidized again as their hydrogen is donated to the electron chain enzymes. Energy that is produced as electrons passes ultimately to oxygen and is indirectly used to power the attachment of inorganic phosphate groups to ADP to create ATP The enzyme responsible for this final step is ATP synthase, whose activity is linked to the movement of hydrogen atoms down a concentration gradient across the membrane of the mitochondria. Some of the energy from the action of the electron transport chain acts to transport hydrogen ions out of the mitochondrial matrix space. The resulting electrochemical gradient drives hydrogen ions back across the membrane in conjunction with ATP synthase leading to ATP generation. Because of the need for oxygen as the final electron acceptor in the electron transport chain, the production of ATP in this manner is called oxidative phosphorylation.Interestingly, the position along the electron transport chain at which FADH2 or NADH + H+ donate their electrons differs. Because of this, the amount of energy that is produced is greater for NADH compared with FADH2. Specifically, each pair of hydrogen atoms from NADH + H+ supplies energy for the creation of three ATP, but the two hydrogen atoms from FADH2 yield only two ATP Most of the proteins of the electron transport chain are closely linked clusters within the inner mitochondrial membrane along with more mobile proteins (coenzyme Q and cytochrome C) that act as carriers between complexes. Figure 3.9 illustrates the release of energy associated with oxidation of NADH or FADH2 in the electron transport chain, and Table 3.4 summarizes ATP production from completed catabolism of glucose.
Under ideal conditions, the complete oxidation of one glucose molecule to CO2 and water yields 36-38 ATP Alternative figures come from uncertainty about the energy yield of reduced NAD+ that is produced
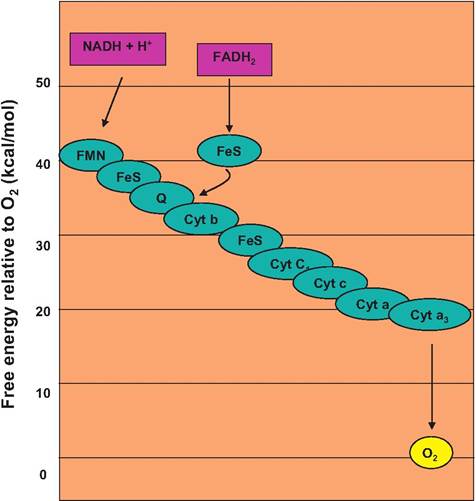
Fig.
3.9. Electron transport chain and reduced coenzymes. Each linked protein oscillates between reduced and oxidized states. As an upper protein is reduced, its capacity to hold the electron is lowered, and the next protein in the cascade captures the electron. This continues until the oxygen atom at the end of the cascade captures the electron. The overall reduction in energy for electrons passed from NADH to oxygen is 53kcal∕mol, but the energy is captured stepwise. Since electrons from FADH2 enter the chain further down, less energy is available so fewer ATP molecules are created.Table 3.4. Summary of ATP production from the Krebs cycle and electron transport chain reactions.
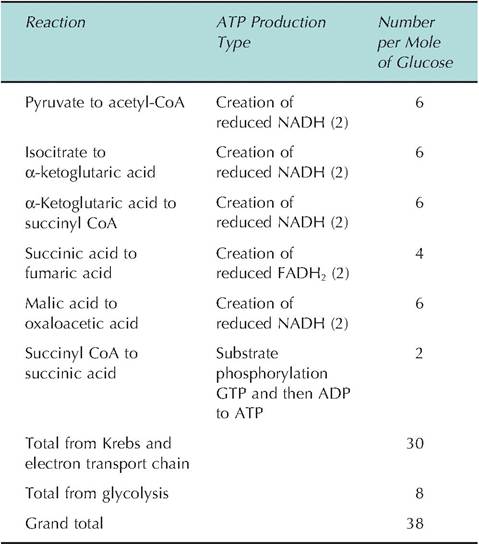
from glycolysis. For these molecules to be utilized, they must be passed across the mitochondrial membrane by active transport. An estimate of this "expense" is that the net ATP gain from reduced NAD+ derived from the cytoplasm is only two ATP per molecule instead of the usual three ATP for those created inside the organelle. Since two of these molecules are produced in the cytoplasm during glycolysis, the total yield is reduced to 36 ATP per molecule of glucose. Regardless, when oxygen is available, energy capture from the biological oxidation of glucose is highly efficient. If a mole of glucose is completely combusted, as in a calorimeter, it yields 686 kcal. Energy obtained in the creation of high-energy ATP bonds equals 262 kcal for an efficiency of 38% [262/686 ? 100]. This is markedly more efficient than most machines. Energy not captured in the formation of ATP is liberated as heat.