Muscular System
Joann Colville
OUTLINE
INTRODUCTION, 211
SKELETAL MUSCLE, 212
Gross Anatomy of Skeletal Muscle, 212
Microscopic Anatomy of Skeletal Muscle, 217
Physiology of Skeletal Muscle, 220
CARDIAC MUSCLE, 223
Gross Anatomy of Cardiac Muscle, 223
Microscopic Anatomy of Cardiac Muscle, 223
Physiology of Cardiac Muscle, 223
SMOOTH MUSCLE, 224
Gross Anatomy of Smooth Muscle, 224
Microscopic Anatomy of Smooth Muscle, 224
Physiology of Smooth Muscle, 224
LEARNING OBJECTIVES
When you have completed this chapter you will be able to:
1.
List the three types of muscle and describe the general characteristics of each type.2. Describe the structure and function of tendons, aponeuroses, and ligaments.
3. Differentiate between prime mover, antagonist, synergist, and fixator muscles.
4. List the locations and actions of muscles of the head and neck.
5. List the locations and actions of muscles of the abdomen, thoracic limb, and pelvic limb.
6. List the locations and actions of muscles of respiration.
7. Describe the microscopic anatomy of skeletal muscle, smooth muscle, and cardiac muscle cells.
8. List the components of a neuromuscular junction and describe the function of each component.
9. List and describe the roles of the connective tissues in skeletal muscles.
10. Describe the events that occur in skeletal muscle cells during muscle contraction and relaxation.
11. Differentiate between visceral smooth muscle and multi-unit smooth muscle.
VOCABULARY FUNDAMENTALS
A band A bahnd
Abductor muscle ahb-duhck-tohr muhs-uhl
Actin filament ahck-tihn fihl-ah-mehnt
Adductor muscle ahd-duhck-tohr muhs-uhl
Adenosine diphosphate (ADP) ah-dehn-o-sen dι-fohs-fat
Adenosine triphosphate (ATP) ah-dehn-o-sen trι-fohs-fat
Aerobic metabolism or-ro-bihck meh-tahb-uh-lihz-ehm
Agonist ahg-uh-nihst
Anaerobic metabolism ahn-or-ro-bihck meh-tahb-uh-lihz-ehm
Antagonist ahn-tahg-uh-nihst
Aponeuroses ahp-uh-noo-ro-sez
Brachium bra-ke-uhm
Cardiac muscle kahr-de-ahck muhs-uhl
Creatine phosphate (CP) kree-ah-ten fohs-fat Cross-bridge krohs brihj
Cutaneous muscle kyoo-ta-ne-uhs muhs-uhl
Dense body dehnz boh-de
Diaphragm dι-ah-frahm
Endomysium ehn-do-mι-se-uhm
Epimysium ehp-ih-mι-se-uhm
Expiratory muscle ehcks-spιr-uh-tohr-e muhs-uhl
Fascicle fahs-ih-kuhl
Fixator fihck-sa-tar
H band H bahnd
I band I bahnd
Inspiratory muscle ihn-spιr-uh-tohr-e muhs-uhl
Intercalated disc ihn-tar-kuhl-la-tehd dihsk
Intramuscular ihn-trah-muhs-kyoo-lar
Involuntary muscle ihn-vohl-uhn-tear-e muhs-uhl
Involuntary striated muscle ihn-vohl-uhn-tear-e strι-a-tehd muhs-uhl
Lactic acid lahck-tihck ah-sihd
Linea alba lihn-e-ah ahl-bah
Motor unit mδ-tar u-niht
Multi-unit smooth muscle muhl-tι-u-niht smooth muhs-uhl
Muscle muhs-uhl
Myofibril mι-δ-fi-brihl
Myoglobin mι-δ-glδ-bihn
Myosin filament mι-δ-sihn fihl-ah-mehnt
Neuromuscular junction nar-δ-muhsk-u-lahr juhngk-shuhn
Nonstriated involuntary muscle nohn-strι-a-tehd ihn-vohl-uhn-tear-e muhs-uhl
Perimysium pear-ih-mihz-e-uhm
Sarcolemma sahr-kδ-lehm-ah
Sarcomere sahr-kδ-mer
Sarcoplasm sahr-kδ-plahz-ehm
Sarcoplasmic reticulum sahr-kδ-plahz-mihck reh-tihck-u-luhm
Sinoatrial node sι-nδ-a-tre-ahl nδd
Skeletal muscle skehl-ih-tahl muhs-uhl
Skeletal muscle fiber skehl-ih-tahl muhs-uhl fι-bar
Smooth muscle smooth muhs-uhl
Synergist sihn-ar-jihst
Tendon tehn-dohn
Transverse tubule (T tubule) trahnz-vars too-byool Twitch contraction twihtch kohn-trahck-shuhn Viscera vih-sar-ah
Visceral smooth muscle vih-sar-ahl smooth muhs-uhl
Voluntary striated muscle vohl-uhn-tear-e strι-a-tehd muhs-uhl
Z line Z lιn
INTRODUCTION
When we think about an animal's body moving, it all seems so simple and automatic.
The animal wants to move forward, so it moves its legs appropriately to walk, trot, or run in that direction. At the same time, things are moving inside its body, too. Blood is being pumped through the blood vessels, food is being moved along the digestive tract, and little adjustments are being made all over to help keep the body operating smoothly. All this just seems to happen, but all these activities and many more are produced by the work of the muscular system.Muscle is one of the four basic tissues of the body (epithelial tissue, connective tissue, and nervous tissue are the other three). It is made up of muscle cells with four common characteristics (1) excitability: they can respond to a stimulus such as a nerve impulse, (2) contractibility: they shorten in length when stimulated, (3) extensibility: they will stretch when pulled, and (4) elasticity: they will return to their original shape and length after contraction or extension. In the body, muscles have three primary functions: to provide motion, maintain posture, and generate heat.
When we hear the word muscle we usually think of large muscles, like the biceps or gluteal muscles. Actually, three different types of muscle make up the muscular system: skeletal muscle (the most familiar kind), cardiac muscle, and smooth muscle (Table 8-1). Skeletal muscle is controlled by the conscious mind and moves the bones of the skeleton so that the animal can move around. This type is what we usually think of as muscle. The other two types are a little less obvious.
Cardiac muscle is found only one place in the body— the heart. It makes up most of the structure of the heart. It starts the entire heart beating long before an animal is born and keeps it going until the animal dies. It has some interesting features, which we will discuss later in this chapter.
Smooth muscle is found all over the body in places such as the eyes, the air passageways in the lungs, the stomach and intestines, the urinary bladder, the blood vessels, and the reproductive tract.
It carries out most of the unconscious, internal movements that the body needs to maintain itself in good working order.In general the nervous system gives the orders, and the muscular system carries them out. This is certainly true of the skeletal muscles, although things are different for cardiac muscle and smooth muscle. They do not require stimulation from nerves to carry out their basic functions; instead, this kind of activity is “built in” to the cardiac and visceral smooth muscle cells. The nervous system can influence cardiac and visceral smooth muscle cells, but only to adjust and modify their basic activities, not to start them.
Like most parts of the body, the muscular system is associated with some strange and unique terminology. The prefix myorefers to muscle generally, and sarco- more specifically refers to muscle cells. For example, myo logy is the study of muscles, and myo sitis is inflammation of muscle tissue. Down at the cellular level, the cytoplasm of a muscle cell is called the sarco plasm. We will use these terms fairly often in this chapter.
TEST YOURSELF 8-1
1. What is muscle?
2. What are the three types of muscle and what are some of the general characteristics of each type?
SKELETAL MUSCLE
Skeletal muscle is the type that usually comes to mind when we hear the word muscle. It is called skeletal muscle because it moves the bones of the skeleton, which in turn move the animal around. You might also hear it referred to by an old name—voluntary striated muscle (called voluntary because it is under the control of the conscious mind). However, not every movement an animal makes is a conscious one. That would be very cumbersome because so many movements are going on all the time. Actually, many skeletal muscle movements, such as maintaining balance and an upright posture, are governed by built-in “cruise control” settings that involve sensory structures, the central nervous system, nerve fibers, and muscle fibers. This kind of system allows animals to breathe, swallow, and stand upright without having to think consciously about each part of the process.
And yet animals can voluntarily change their breathing, swallowing, or standing position. To illustrate this yourself, try consciously to control your breathing—how often you breathe, how much air you take in with each breath, how long you hold it in, and how much air you exhale. Pretty soon the process of breathing becomes a chore. The antidote is to think about other things and let the cruise control kick back in. By the time you read the next few paragraphs, your breathing cruise control will probably have taken over again.The striated part of skeletal muscle's alias (voluntary striated muscle) comes from its microscopic appearance. Even under low-power magnification, skeletal muscle cells are obviously striped (striated) (see Figure 5-29, A). Alternating, crosswise, dark and light bands run the length of each cell. Under higher magnification, the pattern of bands appears more complex than just dark and light bands. We will look into what gives skeletal muscle that appearance when we discuss its microscopic anatomy.
GROSS ANATOMY OF SKELETAL MUSCLE
By gross anatomy we mean those features that can be seen with the unaided eye; that is, without microscopes or magnifying glasses. (Some people think all anatomy is gross, but that's another story.)
A skeletal muscle is a well-defined group of muscle cells surrounded by a fibrous connective sheath called the epimysium. Skeletal muscles come in a variety of shapes and sizes, but they usually have a thick central portion called the belly and two or more attachment sites that join them to whatever structures they move when they contract.
SKELETAL MUSCLE ATTACHMENTS
Most skeletal muscles are attached to bones at both ends by tough, fibrous connective tissue bands called tendons that are a continuation of the epimysium. However, as usual, a few oddballs exist. Instead of connecting to bandlike tendons, some muscles are attached to bones or to other muscles by broad sheets of fibrous connective tissue called aponeuroses.
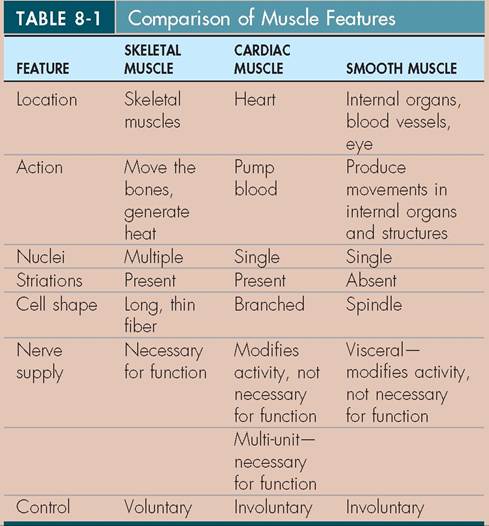
The most prominent aponeurosis is the linea alba (white line) (Figure 8-1) that runs lengthwise between the muscles on an animal's ventral midline. It connects the abdominal muscles from each side together and is a common site for surgical entry into the abdomen (see the Clinical Application on abdominal incisions).One of a skeletal muscle's attachment sites is generally more stable (moves less) than the other. This more stable site is called the origin of the muscle. It does not move much when the muscle contracts. The insertion of the muscle is the site that undergoes most of the movement when a muscle contracts. For example, the origin of the gastrocnemius muscle (the muscle on the back of your calf) is on the femur; its insertion is on the calcaneal tuberosity of the fibular tarsal bone (the point of the hock). When the gastrocnemius muscle contracts, it pulls on the calcaneal tuberosity. In a standing animal contraction of the gastrocnemius muscle flexes the knee and extends the tarsus, which helps propel the animal forward (Figure 8-2).
TEST YOURSELF 8-2
1. What is the difference between a tendon and an aponeurosis?
2. What is the origin of a muscle? The insertion?
3. Why might it be of clinical importance to know the origin and insertion of a muscle?
SKELETAL MUSCLE ACTIONS
A skeletal muscle does only one thing, but it does it really well. When stimulated by a nerve impulse, a muscle contracts (shortens). By pulling on its attachment sites (its origin and insertion), the contraction of the muscle produces movement of bones and other structures. Skeletal muscles rarely act alone. They usually work in groups, with certain muscles producing most of the desired movement
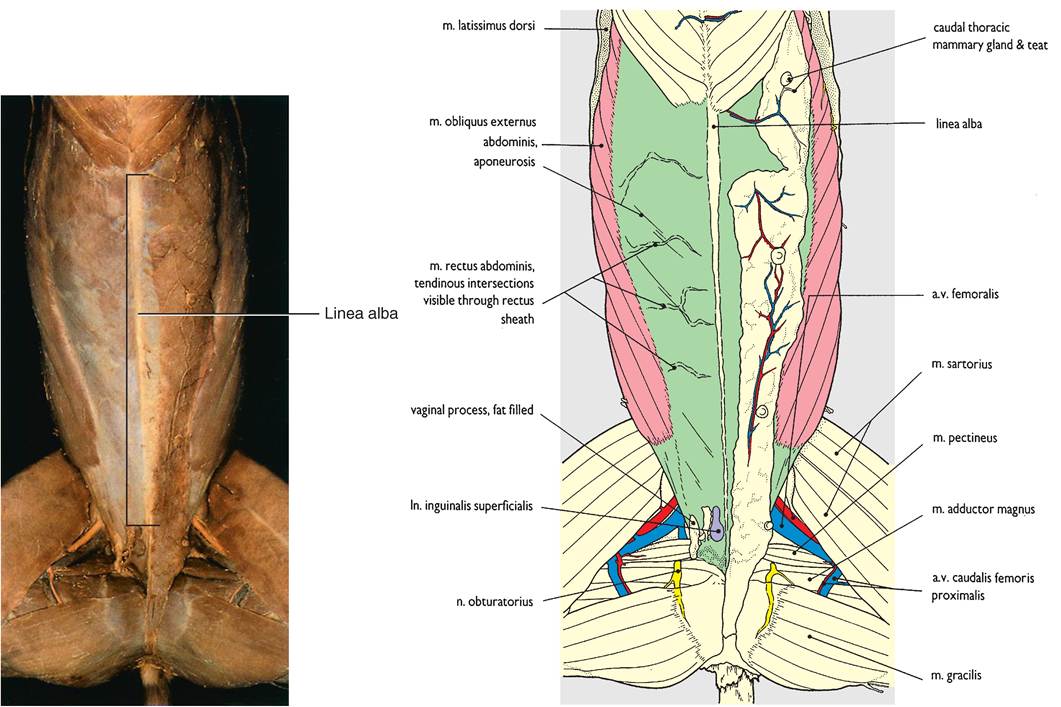
FIGURE 8-1 Linea alba.Theaponeurosisof severa I abdomina I muscles. (From Done S et al: Color atlasofveterinary anatomy, sal 3, St Lauio, 2009, Eloasiar Ltd.)
and others stabilizing nearby joints and providing smooth control over body movements.
The term agonist (or prime mover) is used to describe a muscle or muscle group that directly produces a desired movement.An antagonist is a musclc or muscle group that directly opposes the action of an agonist. Through partial contraction^ antagonists can help smooth out the movements of agonists, or they can contract forcefully at the same time as the agonist, resulting in rigidity and lack of motion. For example, the biceps brachii muscle that flexes (bends) the elbow and the triceps brachii mn^le that extends (straightens) the elbow can each act as an agonist or antagonist, depending on the movement desired.
A synergist is a skeletal muscle that contracts at the same tsgniomneisat a a and assists it in carrying out its action. For example, the deep digital flexor nπιole flexes the digits of the front limb and at the same time the superficial digital flexor muscle acts as a synergist to aid the motion. Fixator UiuscIcs stabilize joints to allow other movements to take place. For eoxmame ple, s of the muscles that flex the digits also can flex
the carpus, or wrist. If a muscle that extends the carpus contracts at Ae same time as a digital flexor muscle, it fixes the carpus in ^ce (prevents it from moving) while the digits are pulled into a flexed position. You can demonstrate this by istharting w the fingers of one of your hands extended and then tightly squeezing them into a fist while feeling your fuosrceleasrm m with your other hand. You will be able to feel tuhseclmes on the underside of your forearm contracting to onfluegxerrys, fi but you will also feel muscles on the top of yooreuarrmf contracting to stabilize your wrist (carpus).
Movements of the body are complex, so each skeletal ymuscle ma fulfill all four of these roles at one time or another. For one type of movement a muscle may act as the agonist, but for others it may act as an antagonist, a synergist, or a fixator.
MUSCLE-NAMING CONVENTIONS
Among the biggest causes of misery for students of anatomy adrde the o and seemingly random names given to muscles. ourely anatomists must be sadistic ogres who delight in thinking up the most obscure and complicated names oossible for body structures, especially muscles? Actually, tghicere is lo behind the names given to most muscles. feTtehney ar o named for physical characteristics, such os the following:
• Action: A portion of a muscle's name is often related to its function. Muscles that flex a joint are often called flexor muscles. For example, the action of the superficial digital
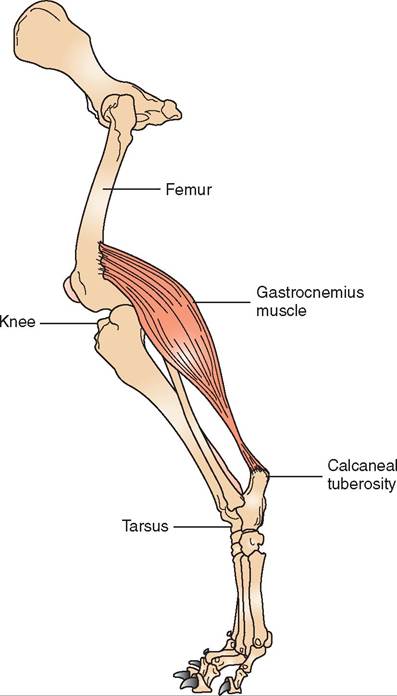
FIGURE 8-2 The gastrocnemius muscle. When the muscle contracts it pulls on the calcaneal tuberosity, causing the knee joint to flex and the tarsal joint to extend. (From Cunningham JG: Texbook of veterinary physiology, ed 4, St Louis, 2007, Elsevier.)
flexor muscle is fairly apparent from its name. It flexes the digits when it contracts. Extensor muscles do the opposite in that they extend joints.
• Shape: A muscle's name can reflect its distinctive shape, such as with the deltoid muscle. “Deltoid” means triangular. The deltoid muscle is a triangular muscle that is located in the shoulder region.
• Location: A muscle's name can indicate its physical location in the body. For example, the biceps brachii muscle is located in the brachial (upper “arm”) region.
• Direction of fibers: The term “rectus” means straight. The rectus abdominis muscles are two straplike muscles on either side of the linea alba on the ventral abdomen. (When someone who lifts weights is referred to as having “six-pack abs,” the rectus abdominis muscles are being noticed.) The fibers of the rectus muscles run straight lengthwise with the long axis of the body and parallel to each other.
• Number of heads or divisions: The number of heads refers to the number of attachment sites that a muscle has to its origin. From the term “cephal,” meaning head, comes the combining form -cep. We can then deduce that the biceps brachii muscle has two heads, the triceps brachii muscle has three heads, and the quadriceps femoris muscle has four heads.
• Attachment sites: Origin and insertion sites are used to name some muscles. For example, the origin of the ster- nocephalicus muscle is the sternum, and its insertion is (the back of) the head.
SELECTED SKELETAL MUSCLES
Animals have several hundred muscles in their bodies. Complete descriptions of each of them can be found in larger anatomy textbooks. Rather than attempting to catalog all the muscles in the common domestic animal species, we will discuss only muscles that are of clinical importance or those that can be used as reference points or landmarks on an animal's body. Figure 8-3 shows the locations of many of the superficial muscles in the horse. The general arrangement of muscles in other species is similar.
CUTANEOUS MUSCLES. Have you ever watched an animal twitch its skin to get rid of an annoying insect? If so, you have seen that animal contracting one of its cutaneous (skin) muscles. Actually, the muscles are not in the skin itself but are in the connective tissue (fascia) just beneath it. Unlike most skeletal muscles, the cutaneous muscles have little or no attachment to bones. They are thin, broad, and superficial and serve only to twitch the skin. (It's a pity we humans lack cutaneous muscles. They could save us a lot of swatting during mosquito season.)
HEAD AND NECK SKELETAL MUSCLES. The muscles of the head have many roles. They control facial expression, enable chewing (mastication), and move sensory structures such as the eyes and ears. The muscles of the neck help support the head and allow the neck to flex, extend, and move the head laterally. The large masseter muscle in the cheek area of the skull is the most powerful of the chewing muscles. Its main action is to close the jaw. Two of the main muscles that extend (raise) the head and neck are the splenius and trapezius muscles, which are located on the dorsal (upper) part of the neck. Another muscle that extends the head and neck and also pulls the front leg forward is the brachiocephalicus muscle. It is a fairly large, straplike muscle that runs from the proximal area of the humerus (brachio-) up to the base of the skull (cephalicus). Neck flexor muscles are located on the ventral (lower) portion of the neck. The sternocephalicus muscle is a smaller, straplike muscle that extends from the sternum to the base of the skull and acts to flex (lower) the head and neck. Flexors of the head and neck do not have to be particularly large or strong because gravity helps them lower the head and neck.
ABDOMINAL SKELETAL MUSCLES. The most obvious function of the abdominal muscles is to support the abdominal organs. However, that's not all they do. They also help flex (arch) the back and participate in various functions that involve straining, such as expulsion of feces from the
FIGURE 8-3 Major superficial muscles of a horse.
rectum (defecation), urine from the bladder (urination), a fetus from the uterus (parturition), and the processes of vomiting and regurgitation. Abdominal muscles also play a role in respiration. We discuss their respiratory role more in tchtieonse on respiratory muscles.
The abdominal muscles are arranged in layers. From outside in, they are the external abdominal oblique muscle, the internal abdominal oblique niueHe, the rectus abdominis muscle, and the transversus abdominis muecle. ^e left and trsight par of each muscle come together on the ventral midline at the linea alba (the aponeurosis that extends from the xiphoid process [caudal end] of the sternum to the cranial ⅛n of the pubis).
The oblique muscles are given that name because their fibers ran in an oblique (slanting) direction to the long axis of the body and opposite to each other. The fibers of the external abdominal oblique muscle ran in a caudoventral (backward and downward) direction. The internal abdominal oblique mn^e fibers run in the opposite direction, which iasnicorventral, or forward and downward. The rectus abdominis Imiscle forms the floor (ventral portion) of the muscular abdominal wall. It consists of two straplike muscles iotnheer side of the linea alba that run from the ribs and sctkernum ba to the brim of the pubis. The transversus abdominisumscle is the deepest of the abdominal muscles. Ieutrnss fib r directly downward in a ventral direction to innsert o the linea alba.
THORACIC LIMB SKELETAL MUSCLES. The muscles of the thoracic (front) limb function mainly in locomotion, ltohwerienbgy al the animal to walk and run and generally omuonvde ar its environment. The primary muscles we will ediscuss ar the large muscles of the shoulder and brachial (upper “arm”) regions, although we do touch on the smaller, ebruyt v important, muscles of the lower leg.
The superficial muscles of the shoulder region are the latissimus dorsiumscle, the pectoralumscles, and the deltoid hmeuscle. T latissimus dorsiumscle is a broad, triangular muscle that extends from the spinal column down to its innsertion o the humerus. It flexes the shoulder, which helps propel the body forward. Usually two pectoral muecles, one superficial and one deep, are located on each side of the shteyrnum. T both extend from the sternum to the humerus and act as adductor muscles (inward movers) oS the front leg. uAsdcdleusctor m help keep the front legs under the animal raenvde npt them from splaying out to the sides. The deltoid muscle is also triangular and extends from the lateral portion of the scapula down to the humerus. It is an abductor muscle (outward mover) and fl∞s the shoulder joint.
As mentioned earlier, the names of the biceps brachii muscle and triceps brachii musdes reveal their general location and basic physical appearance. Both are muscles of the brachium, om upper ⅛n,1 region, and they have opposite actions on the elbow joint. The biceps brachii mnde has two proximal head attachments and extends from the distal end of the scapula to the proximal end of the radius. When it contracts, it flexes the elbow joint. The triceps brachii mnde rehoeaxsimtharl p head attachments and extends from the distal scapula and proximal humerus down to the olecranon process of the ulna (the point of the elbow). When it contracts, it e^⅛nds the elbow joint.
The muscles distal to the elbow joint are an important collection of carpal and digital flexors and extensors that play important roles in locomotion. Their names often reveal their actions and something about their location. They have names life extensor carpi radialis (∣extemls the carpus and is located over the radius), and deep digital flexor (fl^s the digit and is located beneath some of the other digital flexor muscles). Despite the general similarities in these muscles eacmieosn, g sp their precise locations, names, and actions
vary greatly. Consult more in-depth anatomic references if eyodu ne more information about these muscles.
PELVIC LIMB SKELETAL MUSCLES. Like the thoracic luismclbesm, the pelvic limb muscles are involved mainly in locomotion. The large gluteal muscles and the hamstring muscle group are extensor muscles of the hip joint. These powerful m nscles help propel the body forward by extending the hip joint (pulling the leg backward). The gluteal muscles eoxmtend fr the bones of the pelvis down to the trochanters eomf tuhre. f The hamstring muscle group is three muscles located on the caudal part of the thigh region: the biceps femoris muscle, the semimembranosus muscle, and the semi- tendinosushumsecyle. T not only help extend the hip joint ebut also ar the main flexors of the stifle joint.
The quadriceps femoris muscle is the main extensor muscle of the stifle joint. It is located in the cranial part of tehhgeiotnh.ig r When an animal has taken a stride with its hind leg, the quadriceps femoris muscle helps bring the leg forward to prepare for the next stride. As its name implies, iotmispcosed of four heads.
xTohres fle and extensors of the tarsus and digits are soimilar t the flexors and extensors of the carpus and digits ofthe front legs. One important landmark muscle in some species is the gastrocnemius ncu^ede, which is the equivalent of our main calf muscle. It extends from the caudal portion of the distal end of the femur and inserts on the calcaneal tuberosity of the fibular tarsal bone (the point of the hock). aTshtreodcinsetaml igus tendon in humans attaches to our
heel and is called the Achilles tendon. The gastrocnemius muscle is a powerful extensor muscle of the hock. It also rhoeplpesl p the body forward as an animal takes a stride.
SKELETAL MUSCLES OF RESPIRATION. The muscles of renpiration increase and decrease the size of the tho- rvaitcyic ca to draw air into, and push air out of, the lungs. Because drawing air into the lungs is called inspiration, the
CLINICAL APPLICATION
Abdominal Incisions
Abdominal surgery is commonly performed on veterinary patients. From rumenotomies in cattle to ovariohysterectomies (spays) in dogs, abdominal surgical procedures have one thing in common—the surgeon must make an incision somewhere in the abdominal muscles to expose the contents of the abdomen. The location of the incision is usually carefully selected to offer maximum exposure of the required organ or structure. It will also allow a secure closure when the srourcgedicuarlep is over and the incision is sutured shut.
The pnsitions and arrangements of the abdominal muscles and the direction in which their fibers run are important considerations when choosing the site for an abdominal incision. De most common abdominal incision site is the ventral midline, whue the linea alba is located. It offers several advantages ∞^ other sites, such as excellent exposure of abdominal organs, easy closure, and few sensory nerves. Nearly all abdominal organs and tissues can be reached through a ventral midline incision. Also, because all of the abdominal muscles eoma together at the linea alba, an incision through it opens the abdomen in one cut. When it is time to close the abdomen, one layer of sutures (stitches) in the linea alba can effectively aud securely close the abdominal cavity. The linea alba contains fewer sensory nerves than the adjacent muscles, therefore less perioperative pain is involved with a ventral midline inci- istihon than w other abdominal incision sites. The only real disadvantage of a ventral midline incision is that the weight lof al the abdominal contents presses on it during the healing process, so it must be closed with very secure sutures.
At tι'mes, however, a ventral midline incision is not practical, sueh as when a cesarean section must be performed on o cow. De complicated digestive system of a ruminant eanimal lik a cow can make it dangerous to position the annimal o its back for a surgical procedure. Therefore abdominal surgery in cattle is often done with the animal standing and wide awake. Local anesthetic blocks are used to numb the flank (side) area. The incision is usually made in an up-and-down (dorsal-ventral) direction in the flank area. This means that three layers of muscle—the external abdominal oblique, internal abdominal oblique, and transversus abdominis muscles—must be cut to gain access to the abdominal cavity. To minimize trauma and allow normal feurnction aft surgery, many surgeons suture each muscle lvaiydeuralilnydi according to the direction that its fibers
run. Optimal closure of this type of incision requires a sepa- fryaetre la o sutures for each muscle layer that was incised, which entaiIs a lot more work than suturing closed a ventral midline incision.
Several other common abdominal incisions can be used, such as the paramedian incision (parallel to, but beside, the ventral midline), the paracostal incision (parallel to, and just behind, the last rib), and the transverse incision (crosswise, perpendicular to the linea alba). The abdominal muscles present at each incision site determine how the abdominal cavity should be entered and how it can be sutured most securely. (fee, this anatomy stuff is important even after you’re dune with your anatomy class!) muscles that increase the size of the thoracic cavity when they contract are called inspiratory muscles. Pnsliing air out uf the l nngs is called expiration, so the muscles that decrease the thoracic cavity size are called expiratory muscles.
The main inspiratory muscles are the diaphragm and the external intercostal muscles, ⅞e diaphragm is a thin, domeshaped sheet of skeletal muscle that separates the thoracic cavity from the abdominal cavity. The convex surface of its edome shap protrudes into the thoracic cavity. The caudal- bmeosst lo of the lunis are in contact with the diaphraim, and the liver is just behind (caudal to) it. When the diaphragm contracts, it flattens out somewhat. This pushes the abdominal organs caudally. It also increases the size of the tvhitoyr,acic ca causini air to be drawn into the lunis.
The external intercostal muscles h? the same general inspiratory effect, but they accomplish it by a different mech- hnism. The word intercostal means between ribs. Tmimals tohsave tw se of intercostal muscles, located between each dair ot adjacent ribs. The external intercostal muscles are inspiratory muscles, and the deeper, internal intercostal emuscles ar expiratory muscles. The difference is the orientation of their fibers. The fibers of the external intercostal muscles are directed in an oblique direction so that when tohnetyracct they rotate the ribs upward and forward. This increases the size of the thoracic cavity and causes air to be dorawn int the lungs.
Expiration (pushing air out of the lungs) does not require faufscohrmt e as inspiration, because mechanical forces, sruavcihtya, s g and the elastic nature of the lungs help col- iapse the rib cage and push air out. Nonetheless, two sets of expiratory muscles that aid the process are the internal intercostal vhis⅛s and the abdominal muscles. ⅞e fibers of the ^cernal intercostal muscles run at right angles to those of the external intercostal muscles. When the internal intercos- tuaslclmes contract, they rotate the ribs backward, which decreases the size of the thorax and pushes air out of the lungs. When abdominal muscles contract, they push the abdominal organs against the caudal side of the diaphragm. This pushes the diaphragm back into its full dome shape and decreases the size of the thorax. The contributions of the abdominal muscles to breathing become important mainly when animals are breathing hard and fast, such as when they axreerteing themselves physically.
MICROSCOPIC ANATOMY OF SKELETAL MUSCLE
SKELETAL MUSCLE CELLS
Slceletal muscle cells are large cells. They are not very wide, but they are quite long. Most body cells are a few micrometers (μm) in length or diameter (1 μm = O.OOS millimeter [mm]). Skeletal muscle cells can be several inches long. Ti inch is equal to about 25 mm, or 25,000 μm, which is
∕j CLINICAL APPLICATION
Intramuscular Injection Sites
Because skeletal muscles have large blood supplies, drugs injected into them are absorbed into the bloodstream and quickly carried off to the rest of the body. This method of drug administration is called an intramuscular injection (IM injection), and it is commonly used, particularly when a rapid drug effect is desired. Ao intravenous injection (IV injection), into a Wtι, provides the fastest method of drug distribution. An IM injection is the next fastest.
In theory we should be able to use any skeletal muscle for an intramuscular injection. In practice, however, only a few muscles are suitable m each species. Many muscles are either too small or ton thin to alow such an injection; others have prominent structures nearby, such as nerves that could be damaged by the injec- sion. T be useful for an intramuscular injection, a muscle must be fairly large, must be easily accessible, and must have a sufficiently Wck “belly” into which we can deposit the drug.
The following are some common intramuscular injection sdites use in domestic animals.
Cats and Dogs
Pelvic Limb
Gluteal muscles
Quadriceps femoris muscle
Gastrocnemius muscle
Hamstring group (biceps femoris, semimembranosus, and semitendinosus muscles)
Thoracic Limb
Triceps brachii muscle
Cattle and Goats
Pelvic Limb
uGslculteesal m
Hamstring group
Thoracic Limb
Triceps brachii muscle
Neck
Trapezius muscle
Horses
Pelvic Limb
uGslculteesal m
Hamstring group
Thoracic Limb
Triceps brachii muscle
Neck
Tsapezius muscle
Chest
Pectoral muscles
Swine
Pelvic Limb
Semitendinosus muscle
Neck
Brachiocephalicus muscle
Tsapezius muscle
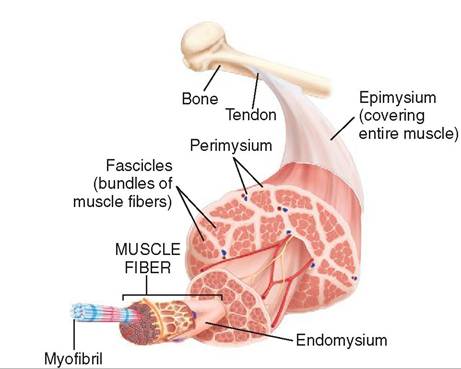
FIGURE 8-4 Structure of skeletal muscle. Skeletal muscle is composed of bundles of muscle fibers called fascicles. Each bundle is surrounded by the connective tissue perimysium. A single muscle fiber is surrounded by the connective tissue endomysium and is made up of smaller myofibrils. (From Thibodeau G, Patton K: Structure and function of the body, ed 14, St Louis, 2012, Elsevier.)
really large on a cellular scale. Despite being so long, skeletal muscle cells are very thin (up to 80 μm in diameter). This gives them an overall threadlike or fiberlike shape. In fact, skeletal muscle cells are usually called fibers rather than cells (Figure 8-4).
Aside from their large size, skeletal muscle fibers have some other unique characteristics. Instead of having just one nucleus, like most cells, skeletal muscle fibers have many. Large fibers can have 100 or more nuclei per cell, all located out at the edge of the cell just beneath the sarcolemma (muscle cell membrane). This reflects the fiber's development from numerous primitive muscle cells that fused. The interior of a muscle fiber is even more interesting. Most of the volume of one skeletal muscle fiber is made up of hundreds or thousands of smaller myofibrils packed together lengthwise, which are themselves composed of thousands of even tinier protein filaments. Prominent organelles between the myofibrils in a muscle fiber include many energyproducing mitochondria, an extensive network of sarcoplasmic reticulum (a storage organelle for calcium ions), and a system of tubules called transverse tubules, or T-tubules, that extend in from the sarcolemma (cell membrane) (Figure 8-5). We will look at the important roles that these organelles play in muscle cells when we discuss how they contract.
As mentioned above, one myofibril is made up of a series of protein filaments. These filaments form the contractile units of a myofibril. Each one of these contractile units is called a sarcomere and is the basic contracting unit of skeletal muscle. There are many sarcomeres laid end to end in one myofibril. Each sarcomere has a disc on each end called the Z line or Z disc. Sarcomeres share discs, so there is one common disc between adjacent sarcomeres. Within a sarcomere there are two primary protein filaments that are responsible for contraction. There are thin protein filaments called actin that attach to the Z lines and extend toward the
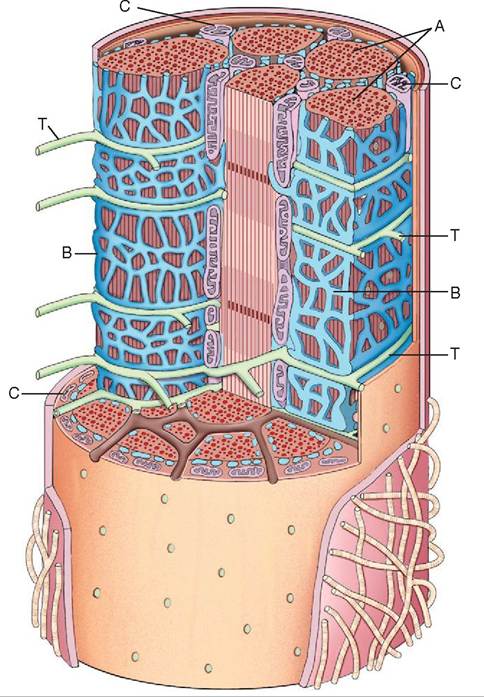
FIGURE 8-5 A three-dimensional view of the internal structure of a muscle fiber. A, Myofibril. B, Sarcoplasmic reticulum. C, Mitochondria. T, Transverse tubules (T-tubules).
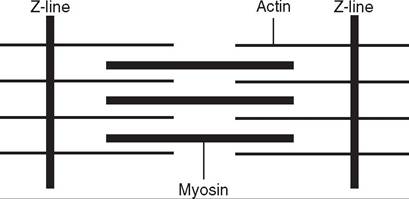
FIGURE 8-6 A sarcomere. (From Cunningham JG: Textbook of veterinary physiology, ed 4, St Louis, 2007, Elsevier.)
center of the sarcomere, but don't meet. There are also thick protein filaments called myosin that appear to float in the middle of the sarcomere between parallel actin fibers. They don't connect to the Z lines (Figure 8-6).
Looking at a myofibril at a higher magnification we can see large light-colored bands (Figure 8-7). These are called I bands and are made up of the thin actin filaments. Each I band extends from one end of the thick myosin filaments in one sarcomere across the Z line to the beginning of the myosin fibers in the next sarcomere. In the center of the I band is the dark Z disc or line that is the attachment site for
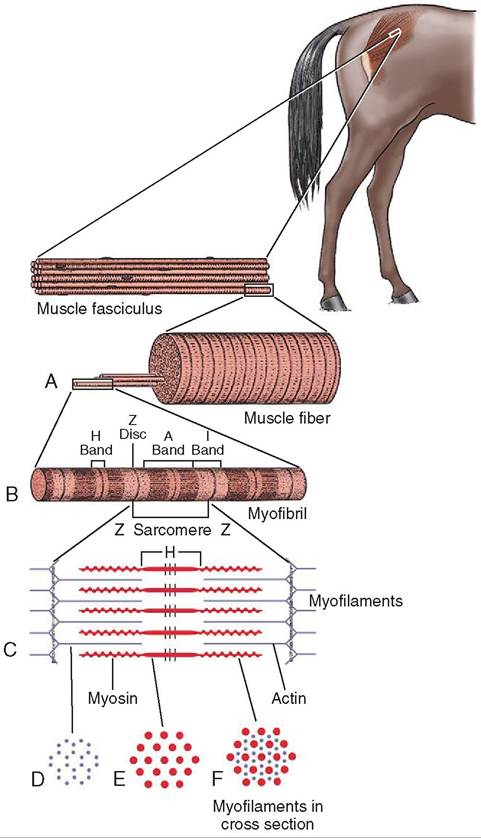
FIGURE 8-7 Ultrastructure of a sarcomere. A, One muscle fiber with an extended myofibril. B, One myofibril composed of four sarcomeres (each one runs from Z line to Z line). C, One sarcomere composed of actin (thin) filaments and myosin (thick) filaments. D, A cross section of the I band (actin filaments only). E, A cross section of the H band (myosin filaments only). F, A cross section of the A band (actin and myosin filaments). (From Hodgson D, McGowan C, McKeever K: The athletic horse:
the actin filaments. From one Z line to the next Z line is one sarcomere. Between the light I bands are darker bands called A bands. They are areas where the thick myosin filaments and thin actin filaments overlap. The H band is the lightcolored area located in the middle of the A band. It is made up of myosin filaments only, with no overlapping actin filaments, so the H band doesn't cover the entire width of the myosin filament.
Looking at one sarcomere you can see that the myosin filaments don't extend all the way from one Z line to the next. There is part of an I band between the end of the A band and the Z line. The I band crosses the Z line and continues until it comes to the beginning of the next myosin filament, where the A band begins again.
The actin fibers are actually two strands of protein twisted together to form a helical structure similar in appearance to a DNA molecule. The myosin molecule has a twisted tail attached to two globular heads that form cross-bridges to actin and interact with the actin to shorten the sarcomere during muscle contraction (Figure 8-8).
TEST YOURSELF 8-3
1. Describe a skeletal muscle cell in terms of cell size, shape, number of nuclei, and appearance under the microscope.
2. What are the differences among a skeletal muscle fiber, a skeletal muscle myofibril, and a skeletal muscle protein filament?
3. Which contractile protein filaments make up the dark bands of skeletal muscle cells? Which make up the light bands?
4. What is a sarcomere and what are its components?
NEUROMUSCULAR JUNCTION
Skeletal muscle is under conscious, voluntary control. Unless it receives nerve impulses, it does not do anything. If a skeletal muscle's nerve supply is interrupted for a lengthy period as a result of injury, the muscle will not only lack the ability to function, it will shrink down through a process called atrophy.
principles and practice of equine sports medicine, ed 2, St Louis, 2014, Saunders. Adapted with permission from Gloom W, Fawcett DW: A textbook of histology, Philadelphia, 1986, Saunders.)
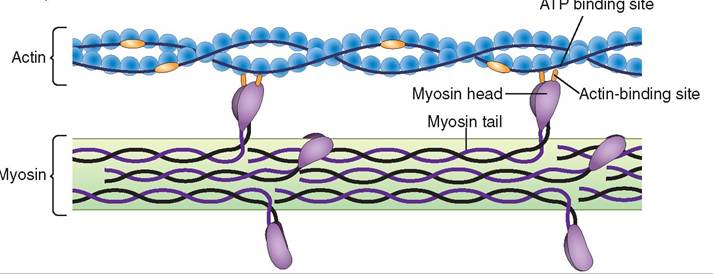
FIGURE 8-8 Actin and myosin. Note the cross-bridges formed between the heads on the myosin tails and the actin filament.
Sites where the ends of motor nerve fibers connect to muscle fibers are called neuromuscular junctions. However, the word connect is not accurate, because a very small space—called the synaptic space—exists between the end of the nerve fiber and the sarcolemma (cell membrane) of the muscle fiber (Figure 8-9 A). Within the end of a nerve fiber in a neuromuscular junction are tiny sacs called synaptic vesicles that contain the chemical neurotransmitter acetylcholine. When a nerve impulse comes down the motor nerve fiber, it causes the release of acetylcholine, which quickly diffuses across the synaptic space and binds (attaches) to receptors on the sarcolemma. This starts the process that leads to the contraction of the muscle fiber. (We explore this process more fully in the section on skeletal muscle physiology.) The effect of acetylcholine on its receptor is very short. The enzyme acetylcholinesterase found in the synaptic space quickly removes the acetylcholine molecule from its sarcolemma receptor and splits it apart. This ends the effect of that nerve impulse. If the body needs to contract the muscle fiber again, it must send down another nerve impulse.
Each nerve fiber innervates (sends impulses to) more than one muscle fiber. The number of muscle fibers per nerve fiber determines how small a movement will result from a nerve stimulus. The term motor unit is used to describe one nerve fiber and all the muscle fibers it innervates (Figure 8-9 B). Muscles that must make very small, delicate movements, such as the muscles that position the eyes, have only a few muscle fibers per nerve fiber in each motor unit. On the other hand, large, powerful muscles, such as leg muscles, may have a hundred or more muscle fibers per motor unit. This allows the nervous system to control the activities of the skeletal muscles in an economical manner. If each nerve fiber attached to only one skeletal muscle fiber, immense numbers of nerve impulses would be constantly necessary to control the muscles' activities. That would require so much work that the nervous system would not be able to do anything else.
CONNECTIVE TISSUE LAYERS
Because they exert a lot of force when they contract, skeletal muscle fibers must be securely fastened together and securely fastened to the structures (usually bones) they move. A delicate connective tissue layer called the endomysium surrounds each individual skeletal muscle fiber. It is composed of fine, reticular fibers. Groups of skeletal muscle fibers, called fascicles, are bound together by a tougher connective tissue layer, called the perimysium, which is composed of reticular fibers and thick collagen fibers. Groups of muscle fascicles are surrounded by epimysium, a fibrous connective tissue layer composed largely of tough collagen fibers. The epimysium is the outer covering of the entire muscle (see Figure 8-4). These three connective tissue layers are continuous with the tendons or aponeuroses that connect the muscle to bones or other muscles. So they not only hold the components of the muscle together but also help fasten the muscle firmly to its attachment mechanisms.
Aside from holding the muscle firmly together and attaching it to the appropriate structures, the connective tissue layers of a muscle also contain the blood vessels and nerve fibers that supply the muscle fibers. They commonly contain varying amounts of adipose tissue, or fat. The fat deposits are often grossly visible in meat and are called marbling.
PHYSIOLOGY OF SKELETAL MUSCLE INITIATION OF MUSCLE CONTRACTION AND RELAXATION
When a nerve impulse travels down a motor nerve fiber and reaches the end bulb at the neuromuscular junction, acetylcholine is released into the synaptic space. The acetylcholine molecules bind to receptors on the surface of the sarco- lemma (cell membrane) of the muscle fiber, which starts an impulse that travels along the sarcolemma and through the T-tubules to the interior of the cell. When the impulse reaches the sarcoplasmic reticulum, it causes the release of stored calcium ions (Ca2+) into the sarcoplasm (cytoplasm). As the Ca2+ diffuses into the myofibrils, it turns on the contraction process, which is powered by high-energy molecules of adenosine triphosphate (ATP). ATP's function of providing cells with energy was discussed with cell metabolism in Chapter 2. We will discuss its role in muscle contraction later, in the section on the chemistry of muscle contraction.
Almost as soon as the sarcoplasmic reticulum releases its Ca2+ into the sarcoplasm, it begins pumping it back in again. This pulls the Ca2+ out of the myofibrils, and the contraction process shuts down. The elasticity of the muscle fiber then restores it to its original length, relaxing the fiber. Pumping the Ca2+ back into the sarcoplasmic reticulum requires energy, which is also supplied by ATP molecules. So not only does muscle contraction require energy, but muscle relaxation does too.
The amount of calcium in the muscle fiber is determined largely by the level of calcium in the bloodstream. If the blood calcium level is too high or too low, abnormalities in skeletal muscle function can result. (See Chapter 11 for a discussion of the hormones calcitonin and parathyroid hormone, which control the blood calcium level, as well as the Clinical Application on hypocalcemia.)
MECHANICS OF MUSCLE CONTRACTION
When a muscle fiber is in a relaxed state, the actin and myosin filaments overlap only a little. When the fiber is stimulated to contract, the globular heads attached to the tails of the myosin filaments, called cross-bridges, which are in contact with the actin filaments, ratchet back and forth and pull the actin filaments on both sides toward the center of the myosin filaments. This sliding of the filaments over each other shortens the sarcomere. The combined shortening of all the end-to-end sarcomeres in a muscle fiber results in what we call a muscle contraction. Microscopically the H band becomes narrower as the actin filaments are pulled toward the center of the sarcomere and the band of overlapping of actin and myosin filaments (the A band) becomes wider.
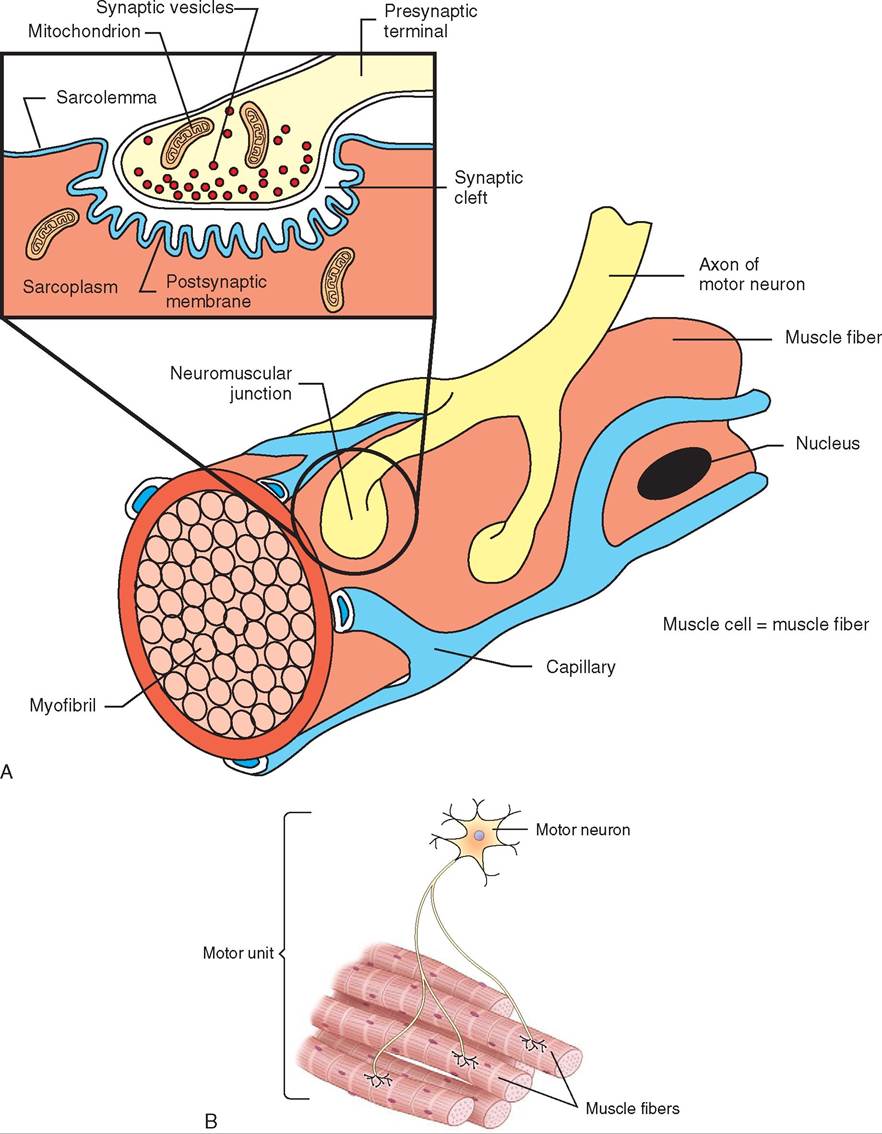
FIGURE 8-9 A, A neuromuscular junction. B, A motor unit showing one motor neuron innervating several muscle fibers.
CHARACTERISTICS OF MUSCLE CONTRACTION
An individ ual muscle fiber either contracts completely when it receives a nerve impulse, or it does not contract at all. This is known as the all-or-nothing principle. We know this is not true of whole muscles, so how does the body produce movements that vary in range and strength when individual muscle fibers are doing all or nothing? It does so by carefully controlling the number oc muscle fibers it stimulates for a particular movement. Small, fine movements require only a few muscle fibers to contract. Larger, more powerful move- emqeunirtse r the contraction of many muscle fibers. The nervous system is calling the shots; therefore, it must predict how large and powerful a movement needs to be, and then it must send the appropriate nerve impulses down to the appropriate muscle fibers in the appropriate muscle(s). This results in what we refer to as the “muscle memory” necessary to perform learned repetitive activities skillfully, such as knitting, shooting a basketball, or walking.
Getting down to the basics of muscle contraction, a single muscle fiber contraction (called a twitch contraction) can ve divided into three phases: (1) the latent phase, (2) the contracting phase, and (3) the relaxation phase. The latent phase is the brief hesitation between the nerve stimulus and the beginning of the actual contraction. It lasts about 0.01 second (10 milliseconds [ms]). The contracting phase lasts about 0.04 second (40 ms), and the relaxation phase lasts about 0.05 second (50 ms). The entire contraction cycle takes about 0.1 second (100 ms). So maximum contraction cefcfiucrisency o if nerve impulses arrive about 0.1 second eahspiusalrtts. T r in a series of complete muscle fiber twitches.
Whole muscles rarely contract by twitching, so how do they contract smoothly? They do so mainly by careful timing of the nerve impulses to the various motor units of the wmiutcshclees. T of individual muscle fibers are stimulated out of phase with each other; that is, they each occur at selriegnhttly diff times. Some muscle fibers are contracting rwshile othe are relaxing. When all the muscle fiber activity is averaged out, smooth, sustained muscle contractions result. Pretty tricky, eh?
CHEMISTRY OF MUSCLE CONTRACTION oTnhseidcerable mechanical work of muscle contraction must be powered by a plentiful supply of energy. The imme- dnieartgey e source that powers the sliding of the actin and myosin filaments is adenosine triphosphate (ATP), rwohdiuchcedis p by the many mitochondria in muscle
fiTbPers. A molecules are like tiny batteries that can release energy and then be recharged so that they can do it again. iArs the name implies, ATP molecules have three phosphate cghroeudps atta to a central adenosine core. When one of the phosphate groups is split off (forming adenosine diphosphate [ADP]), a considerable amount of energy is rheilcehased, w powers the sliding of the actin and myosin hfiilsaments. T also “discharges” the ATP molecule. Another energy source has to reattach the phosphate group to “recharge” the ATP to get it ready to supply energy again.
The “battery charger” that converts ADP back to ATP is another compound in the muscle fiber called creatine phosphate (CP). e^en the CP molecule splits, the energy that is rdedlesased a a phosphate group to the ADP, converting it boTaPc. k t A The newly recharged ATP molecule is ready to pnreorgvyide e for further muscle contraction or relaxation (by helping to pump Ca2+ back into the sarcoplasmic reticulum).
iTmhaeteult source of energy used to produce ATP and eCePp and k the whole system operating comes from the catabolism (breokdown) of nutrient molecules. The two main compounds involved are glucose and oxygen. Glneose iusgars molecule that is the primary energy source for eomldlosy,st b c including the muscle cells. The muscles have aervy larg blood supply that constantly brings new supplies loufcgose and oxygen to the muscle fibers.
uWphpelinesthe s are plentiful and the cells are fairly iunsaccletive, m fibers can also store glucose and oxygen for feudtsu. re ne Glucose is stored in the fibers in the form of glycogen, and oxygen is stored attached to large protein molecules called myoglobin. Lilo So, after a strenuous burst of exercise, an aynimal ma continue to breathe heavily for a while as its body repays its so-called “oxygen debt.”
TEST YOURSELF 8-4
1. What ion, released fromthe sarcoplasmic reticulum by a nerve iepslse, starts the aontraation proaess in a essale fiber?
2. What mo iecsleh in museIesa ct as the " bo tteries" to power the sliding of the aatin and eyosin filaeents? What eoleasles fsnation as the "battery chargers"?
3. If indiviCuol m usrle riOoreontractions obeytho all-or- nothing principle, how does an anieal control the size and strength of its essaslar eoveeents?
4. What ir nt and l obin and wtyisit important?
5. Why anssanonimal brevttn heanilyfen a while after heavy exercise?
HEAT PRODUCTION
Likiie all machines, muscles are less than 100% efficient at cnoenrgvyerting e to useful work (or in this case, motion). oAncsiderable amount of the energy produced in muscles iosrmin the f of heat. In fact, muscular activity is one of the major heat-generating mechanisms that the body uses to maintain a constant internal temperature. If heat produc- txicoenedes body needs, the excess must be eliminated by
CLINICAL APPLICATION
Rigor Mortis
The term used to describe the stiffness of skeletal muscles that occurs shortly after an animal dies is rigor mortis, which is Latin for “stiffness of death.” It would seem more sensible for the muscles to go limp after death, because all nerve stimulation ceases, but chemical reactions at the cellular level send things in another direction.
When the animal dies, lack of oxygen to the cells causes normal activities and barriers within the cells to break down. One of the things that happens in skeletal muscle cells is that most of the Ca2+ spills out of the sarcoplasmic reticulum. This causes contraction of many of the muscle fibers, fueled by the last of the ATP molecules in the sarcoplasm. However, all the ATP is used up in the contraction, and no more is being made; therefore no energy source is available to relax the muscles. The result is that the muscles get stuck in the contracted position. Rigor mortis is not a permanent condition. As soon as the muscle fibers begin to decompose, the cross-bridges between the myosin and actin filaments break down and the muscles go into a relaxed state. When this happens rigor mortis has passed and the body becomes limp.
Forensic experts can use the onset of rigor mortis and the subsequent muscle relaxation to help establish the time of death.
mechanisms such as panting or sweating. Under cold conditions, the body may need to increase the production of heat to avoid hypothermia (too low a body temperature). It often does this by producing the small, spasmodic muscle contractions we know as shivering.
CARDIAC MUSCLE
Cardiac muscle is also known as involuntary striated muscle. It is called involuntary because its contractions are not under conscious control. The striated part of the name is given because under the microscope its cells have the same kind of striped appearance as skeletal muscle cells.
GROSS ANATOMY OF CARDIAC MUSCLE
Cardiac muscle is found in only one place in the body—the heart. It forms most of the volume of the heart and makes up the majority of the walls of the cardiac chambers (the atria and the ventricles). Instead of being organized into distinct muscular structures, like skeletal muscle, cardiac muscle cells form elaborate networks around the cardiac chambers. The arrangement and physical characteristics of cardiac muscle allow it to start contracting early in the embryonic period before birth and to continue contracting without a rest until the animal dies. To get a feel for how amazing that is, make a fist and, for the next minute or two, clench it tightly and then relax it about once per second. Before too long, the forearm muscles that tighten your fist will become fatigued. The heart does this same kind of work, but it gets no rest periods.
MICROSCOPIC ANATOMY OF CARDIAC MUSCLE
Cardiac muscle cells are striated (striped) like skeletal muscle cells, and they contain many of the same organelles and intracellular structures, such as myofibrils. However, cardiac muscle cells and skeletal muscle cells are otherwise very different. Cardiac muscle cells are much smaller than skeletal muscle cells and have only one nucleus per cell. They are not shaped like the long, thin fibers of skeletal muscle. They are longer than they are wide and often have multiple branches. They are securely attached to each other end to end to form intricate, branching networks of cells. The firm, end-to-end attachments between cardiac muscle cells are visible under the microscope as dark, transverse lines between the cells (see Figure 5-29, B). These attachment sites are called intercalated discs. The intercalated discs securely fasten the cells together and also transmit impulses from cell to cell to allow large groups of cardiac muscle cells to contract in a coordinated manner. In fact, the networks of cardiac muscle cells around the cardiac chambers function as if they were each a large, single unit instead of a whole bunch of individual cells.
PHYSIOLOGY OF CARDIAC MUSCLE
MUSCLE CONTRACTIONS
If we looked through a microscope at individual cardiac muscle cells growing in a tissue culture flask, we would see something amazing. Each cell would be contracting rhythmically with no external stimulation at all. Furthermore, each cell would be contracting at a constant rate set by its own internal metronome—some rapidly and others more slowly. However, if two cells touch, the slower contracting cell adopts the faster cell's contraction rate. This demonstrates two unique and important things about cardiac muscle: (1) it contracts without any external stimulation, and (2) groups of cardiac muscle cells adopt the contraction rate of the most rapid cell in the group.
These self-starting and self-controlling aspects of cardiac muscle enable the heart to function as a very efficient pump. Rather than large numbers of muscle cells contracting at the same time, as in skeletal muscle, cardiac muscle cells contract in a rapid, wavelike fashion. The impulse that coordinates the contractions spreads from cell to cell across the intercalated discs like a wave. These rapid, wavelike contractions effectively squeeze blood out of the cardiac chambers, much like milk being squeezed out of a dairy cow's teat at milking time.
For these wavelike contractions of cardiac muscle to move blood effectively through the chambers and valves of the heart and out into the rest of the body, they must be carefully initiated and controlled. This is the role of the heart's internal impulse conduction system, which functions like a “mini nervous system.” This impulse conduction system consists entirely of cardiac muscle cells. The impulse that starts each heartbeat begins in the heart's “pacemaker,” the sinoatrial (SA) node, located in the wall of the right atrium (see Figure 14-17).
Why does the SA node have so much control over things? The reason goes back to that business about cardiac muscle cells adopting the contraction rate of the most rapidly contracting cells in the group. The contraction rate of the cardiac muscle cells in the SA node is faster than those in the walls of the atria or ventricles, therefore its rate takes precedence. The impulse that starts in the SA node follows a carefully controlled path through the conduction system of the heart. Structures in the system transmit, delay, and redirect each impulse so that the cardiac muscle cells in the walls of the heart chambers contract in the coordinated, effective manner necessary to pump blood around the body. Details of the cardiac impulse conduction system can be found in Chapter 14.
NERVE SUPPLY
Although it is not needed to initiate the contractions of cardiac muscle, the heart does have a nerve supply that can modify its activity. We know from successful heart transplants that the heart's nerve supply is not essential to its function. (The nerves to the heart are severed when it is removed from the donor.) So what role does the heart's nerve supply play in controlling the heartbeat?
The nerves to the heart are from both divisions of the autonomic portion of the nervous system; that is, the sympathetic and parasympathetic systems. Sympathetic fibers stimulate the heart to beat harder and faster as part of the fight-or-flight response that kicks in when an animal feels threatened. Parasympathetic fibers do the opposite in that they inhibit cardiac function, thereby causing the heart to beat more slowly and with less force when the body is relaxed and resting. The two opposing systems strike a balance that keeps the heart's activity appropriate for what is going on inside and outside the animal at any particular time. More information about the autonomic portion of the nervous system and its effect on cardiac function can be found in Chapters 9 and 14.
TEST YOURSELF 8-5 the microscope that skeletal muscle and cardiac muscle cells have (see see Figure 5-29 C). Nonstriated involuntary muscle is really very different from the other two types of muscle.
GROSS ANATOMY OF SMOOTH MUSCLE
Smooth muscle is found all over the body but not in distinct structures like skeletal muscles and the heart. Rather, it is found in two main forms: (1) as large sheets of cells in the walls of some hollow organs (visceral smooth muscle) and (2) as small, discrete groups of cells (multi-unit smooth muscle) (Figure 8-10).
MICROSCOPIC ANATOMY OF SMOOTH MUSCLE
Smooth muscle cells are small and spindle shaped (tapered at the ends) with a single nucleus in the center. They have a smooth, homogeneous appearance under the microscope because their filaments of actin and myosin are not arranged in parallel myofibrils as in skeletal and cardiac muscle. Rather, small, contractile units of actin and myosin filaments crisscross the cell at various angles and are attached at both ends to dense bodies that correspond to the Z lines of skeletal muscle (Figure 8-11). When these contractile units shorten, they cause the cell to ball up as it contracts. Because their contractile units are not organized into regular, parallel sarcomeres, individual smooth muscle cells can shorten to a greater extent than skeletal or cardiac muscle cells.
PHYSIOLOGY OF SMOOTH MUSCLE
VISCERAL SMOOTH MUSCLE
Visceral smooth muscle is found in the walls of many internal soft organs, which are also known by the general name viscera. The muscle cells are linked to form large sheets in the walls of organs such as the stomach, intestine, uterus, and urinary bladder. Fine movements are not possible with visceral smooth muscle; rather, it experiences large, rhythmic waves of contraction. These contractions can be quite strong, as in the peristaltic contractions that move food along the
1. Describe a cardiac muscle cell in terms of size, shape, number of nuclei, and appearance under the microscope.
2. What are intercalated discs and why are they important to the functioning of cardiac muscle?
3. Describe the effect of a cardiac muscle's nerve supply on its functioning.
4. What is the general effect of sympathetic nervous system stimulation on cardiac muscle? What is the effect of parasympathetic nervous system stimulation?
SMOOTH MUSCLE
Smooth muscle is also called nonstriated involuntary muscle, or sometimes just involuntary muscle. Like cardiac muscle, it is called involuntary because its contractions are not under conscious control. The smooth part of the name is because its cells do not have the striped appearance under
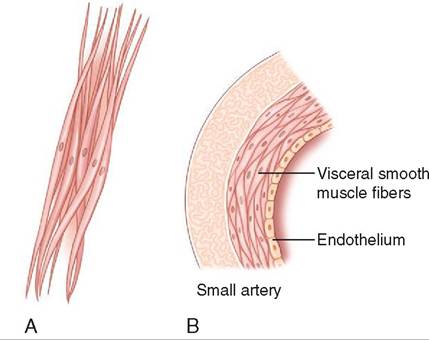
FIGURE 8-10 Multi -unit smooth muscle cells (A) and visceral smooth muscle cells (B).
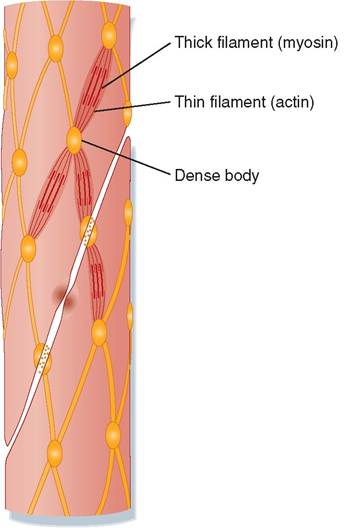
FIGURE 8-11 Smooth muscle cells.(From Koeppen B, Stanton B:
Berne & Levy physiology [updated], ed 6, St Louis, 2010, Mosby.)
gastrointestinal tract and the uterine contractions that push the newborn animal out into the world at parturition (birth).
dike cardiac muscle, visceral smooth muscle contracts without the need for external stimulation. It does react to stretching, however, by contracting more strongly. This is useful in the gastrointestinal tract, where the presence of fuomodenin the l stretches the organ and the smooth muscle ilenspiotsndwsal r with increased contractions that help
move the food along. Something similar happens in the urinary bladder; however, the slow, gradual stretching of the lbladder wal caused by urine accumulation does not trigger contraction of the smooth muscle in its wall until the bladder is nearly full.
In the pregnant uterus it is very important that the smooth muscle in its wall does not contract as the fetus grows and stretches the uterine wall. The would result in a prema- fture loss o the fetus and termination of the pregnancy. The uterus must be kept quiet as the fetus enlarges and develops. cTchoims pislisahed through the activity of hormones,
srougchestaesropne, that inhibit the smooth muscle in
the Iherine wall from contracting during pregnancy. When the time comes to give birth, the level of progesterone in the bloattstream drops dramatically. This removes the inhi- fbition o the uterine smooth muscle, and a combination of factors, including rising levels of other hormones (such as estrogens and oxytocin), stimulates the smooth muscle to choisntract. T starts the process of labor.
dike cardiac muscle, visceral smooth muscle has a nerve supply that is not necessary to initiate contractions but serves to modify them. Also like cardiac muscle, the nerve soupply t smooth muscle consists of the sympathetic and parasympathetic divisions of the autonomic nervous system. However, the effects of the two are the reverse of what is seen in cardiac muscle. Sympathetic stimulation decreases ViTceraI sumscoleoth m activity, and parasympathetic stimulation increases it. This makes sense if we think about sympathetic and paraaympathetic functions. Sympathetic stimulation prepares an animal for intense physical activity. Blood is dwiavyerted a from the viscera and redirected to the heart, skeletal muscles, and brain to help deal with whatever threat initiated the fight-or-flight response. Decreasing gastrointestinal motility as a part of this response makes sense. Digestion is not a priority at this point. On the other hand, when the animal is relaxed and resting, the parasympathetic system predominates and enhances functions such as gastrointesti- contailviaty t help supply nutrients to the body cells during this “down time.” As with the heart, the two opposing autonomic divisions strike a balance to keep smooth muscle activity at an appropriate level for the body's ever- echdas.nging ne
MULTI-UNIT SMOOTH MUSCLE
iWscherearel as v smooth muscle is large and relatively pow-
eurlftui-lu, mnit smooth muscle is small and delicate. Instead of being formed into large sheets that function as a single, large unit, multi-unit smooth muscle is made up of indi- voitdhual smo muscle cells or small groups of cells. It is fhoeurend w small, delicate contractions are needed, such as itshe ir and ciliary body of the eye, the walls of small blood vessels, and around small air passageways in the lungs. Also iusncelirkael v smooth muscle, contractions of multi-unit sumscoleoth m are not automatic. They require specific impulses from autonomic nerves to contract.
The actions of multi-unit smooth muscle are specific and coanrterfoullleydc. This allows fine control of actions, such
as adj^ting the size of the pupil of the eye or the accommodation (focusing) of the lens. It also allows delicate fcontrol o blood flow throughout the body and airflow through the lungs by adjusting the size of blood vessels and air passageways according to the body's needs.
TEST YOURSELF 8-6
1. Describe a smooth muscle cell in terms of its size, shape, number of nuclei, and appearance under the microscope.
2. What are themaindifferencen between visceral smooth muscle and multi-unit smooth muscle?
3. Describe the effect of nerve stimulation on the functioning of visceral smooth muscle vs. multi-unit smooth muscle.
4. What Is the fecerol effect of Symeathetic nervous system stimulation on visceral smooth muscle? What is the effect of parasfmpathetic nervous sfstem stimulation?
5. What are ttn mam s iffe rencet ic the saructures and functions of skeletal muscle, cardiac muscle, and smooth muscle?