Nervous System
Thomas Colville
OUTLINE
INTRODUCTION, 228
NEURONS AND SUPPORTING CELLS, 229 ORGANIZATION OF THE NERVOUS SYSTEM, 230 Anatomic Location: CNS Versus PNS, 230
Direction of Impulses: Afferent Versus Efferent, 231
Function: Autonomic Versus Somatic, 231
NEURON FUNCTION: DEPOLARIZATION AND REPOLARIZATION, 231
Resting State, Polarization, and Resting Membrane Potential, 231
Depolarization, 232
Repolarization, 232
Depolarization Threshold, Nerve Impulse Conduction, and All-or-Nothing Principle, 233
Refractory Period, 234
How Myelinated Axons Conduct Action Potentials Quicker: Saltatory Conduction, 234
HOW NEURONS COMMUNICATE:
THE SYNAPSE, 235
Types of Neurotransmitter and Their Effect on
Postsynaptic Membranes, 236
Stopping and Recycling the Neurotransmitter, 238
THE CENTRAL NERVOUS SYSTEM-BRAIN AND
SPINAL CORD, 238
Cerebrum, 238
Cerebellum, 239
Diencephalon, 240
Brainstem, 240
Other Clinically Important Structures of the Brain, 240 Spinal Cord, 243
THE AUTONOMIC NERVOUS SYSTEM, 244 Structure, 244
General Functions, 246
Neurotransmitters and Receptors, 247
REFLEXES AND THE REFLEX ARC, 247
Stretch Reflex, 248
Withdrawal Reflex, 249
Crossed Extensor Reflex, 249
The Role of the Upper CNS in Moderating Reflexes, 249 Other Clinically Significant Reflexes, 250
LEARNING OBJECTIVES
When you have completed this chapter you will be able to:
1.
Describe the structures and functions of neurons and neuroglia.2. Differentiate between white matter and gray matter.
3. Describe the functions of afferent and efferent nerves.
4. List the components of the central nervous system and the peripheral nervous system.
5. Differentiate between the autonomic and somatic nervous systems.
6. Describe the process of depolarization and repolarization of neurons.
7. List excitatory and inhibitory neurotransmitters and describe their role in the conduction of nerve impulses.
8. Describe the structures and functions of the cerebrum, cerebellum, diencephalon, and brainstem.
9. Describe the connective tissue layers that surround the brain and spinal cord.
10. Explain the functions of the cerebrospinal fluid.
11. List the cranial nerves and describe their functions.
12. Differentiate between the sympathetic and parasympathetic divisions of the autonomic nervous system.
13. Differentiate between autonomic and somatic reflexes.
14. Describe the components of a reflex arc.
15. Describe the stretch reflex, withdrawal reflex, crossed extensor reflex, palpebral reflex, and pupillary light reflex.
VOCABULARY FUNDAMENTALS
Acetylcholine ah-set-ehl-ko-len
Acetylcholinesterase ah-set-ehl-ko-luh-nehs-tuh-ras
Adrenergic neuron ahd-reh-nar-jihk nar-ohn
Afferent a-for-ehnt
Afferent nerve a-for-ehnt norv
All-or-nothing principle awl or nuhth-ihng prihn-suh-puhl
Alpha1-adrenergic receptor ahl-fuh wuhn ahd-reh-nar- jihk reh-sehpt-or
Anesthesia ahn-uhs-the-zhuh
Antiparasitic drug ahn-te-peor-uh-siht-ihck druhg
Arachnoid ah-rahck-noyd
Autonomic nervous system aw-to-noh-mihck narv-uhs sihs-tehm
Autonomic reflex aw-to-noh-mihck re-flehcks
Axon ahck-sohn
Beta1-adrenergic receptor bat-ah wuhn ahd-reh-nar-jihk reh-sehpt-or
Beta2-adrenergic receptor bat-ah too ahd-reh-nar-jihk reh-sehpt-or
Blood-brain barrier bluhd ban bear-e-or
Brainstem bran stehm
Catecholamine kaht-ih-kol-ih-men
Central canal sehn-trahl kuh-nahl
Central nervous system sehn-trahl nar-vuhs sihs-tehm
Cerebellum sehr-eh-behl-luhm
Cerebral cortex seh-re-brahl kohr-tehx
Cerebral hemisphere seh-re-brahl hehm-ih-sfeer
Cerebrospinal fluid seh-re-bro-spι-nahl floo-ihd Cerebrum seh-re-bruhm
Cholinergic neuron ko-luh-nar-jihk nar-ohn
Cholinergic receptor ko-luh-nar-jihk reh-sehpt-or
Conduction of the action potential kuhn-duhck-shuhn of the ahk-shuhn puh-tehn-shuhl
Contralateral reflex kohn-trah-laht-or-ahl re-flehcks
Contrasp radiography kohn-trahst ra-de-ohg-rah-fe Corpus ca^mι kohr-pahs kal-lo-suhm rCvreanial ne kra-ne-ahl norv
Cranial-sacral system kra-ne-ahl sa-krahl sihs-tehm Crossed extensor reflex krohst ehck-stehn-sohr re-flehcks Dendrite dehn-drit
Depolarization de-po-lor-uh-za-shuhn
Diencephalon di-ehn-sehf-uh-lohn
Dopamine do-puh-men
nDorsal hor dohr-nsahl hohr
rDvoersal ne root dohr-sahl norovotr
eDrura mat duhr-ah mah-tor
Effector cell e-fehck-tor sehl
Efferent e-far-ehnt
Efferent nerve e-far-ehnt norv
Endocrine system ehn-do-krihn sihs-tehm
Enzyme ehn-zim
Epidural anesthesia ehp-ih-duhr-ahl ahn-uhs-the-zhuh Epinephrine ehp-ih-nehf-rihn
Excitatory neurotransmitter ehcks-si-tuh-tor-e nor-o-trahnz-miht-or
Fenestration fehn-ih-stra-shuhn
Fight-or-flight response fit or flit reh-spohns
Fissure fihsh-or
GABA gah-buh
Gamma-aminobutyric acid gahm-uh ah-me-no-byoo-
tihr-ihck ah-sihd
Ganglion gahng-gle-uhn
General anesthesia jehn-or-ahl ahn-uhs-the-zhuh
Glial cell gle-ahl sehl
Glycine gli-sen
Gray matter gra maht-or
Gyrus (plural gyri) ji-ruhs (plural ji-ri)
Hormone hohr-mon
Hypermetria hi-por-me-tre-uh
Hyperreflexive hi-por-re-flehcks-ihv
Hyporeflexive hi-po-re-flehcks-ihv
Hypothalamus hi-po-thahl-uh-muhs
Inhibitory neurotransmitter ihn-hihb-ih-tohr-e
nor-o-trahnz-miht-or
Interneuron ihn-tor-nar-ohn
Ipsilateral rφex ihp-sih-lah-tor-ahl re-flehcks
Ivermectin i-vor-mehck-tihn
Lobe lob
Local anesthesia lo-kuhl ahn-uhs-the-zhuh
Longitudinal fissure lohn-jih-tud-ihn-ahl fihsh-or Medulla oblongata meh-duhl-uh ohb-lohng-gah-tah Meninges meh-nihn-jez
Midbrain mihd-bran
rMviexed ne mihckst norv
rMveotor ne mo-tor norv
Motor nrnon mo-tor nar-ohn
Muscarinic receptor muhs-kuh-rihn-ihck reh-sehpt-or
Muscle spindle muhs-uhl spihn-duhl
Myelin mi-eh-lihn
aMtyhelin she mi-eh-lihn sheth
Myelography mi-ehl-ohg-rahf-e
NIerve impulse norv ihm-puhls
Nerve norv
Nerve fiber norv fi-bor
Neuroglia nor-og-le-ah
Neuron nar-ohn
Neurotransmitter nor-o-trahnz-miht-or
Nicotinic receptor nihck-uh-tihn-ihck reh-sehpt-or Node of Ranvier nod of ronn-ve-a
Norepinephrine nohr-ehp-ih-nehf-rihn
Nuclei noo-kle-i
Oligodendrocyte ohl-ih-go-dehn-dro-sit
Palpebral reflex pahl-pe-brahl re-flehcks
Parasympathetic nervous system peor-uh-sihm-puh-
theht-ihck nar-vuhs sihs-tehm
Perikaryon peor-ih-kear-e-ohn
Peripheral nervous system puh-rihf-or-uhl nar-vuhs
sihs-tehm
ePria mat pe-ah mah-tor
Pituitary gland pih-too-ih-teor-e glahnd
Pons pohnz
Postganglionic neuron post-gahng-gle-ohn-ihck nar-ohn Postsynaptic neuron post-sih-nahp-tihck nar-ohn Preganglionic neuron pre-gahng-gle-ohn-ihck nar-ohn Presynaptic neuron pre-sih-nahp-tihck nar-ohn Pupillary light reflex (PLR) pyoo-peh-lear-e lit re-flehcks Receptor reh-sehpt-ar
Reflex re-flehcks
Reflex arc re-flehcks ahrk
Refractory period re-frahck-tar-e peer-e-uhd
Repolarization re-po-lar-uh-za-shuhn
Resting membrane potential rehs-tihng mehm-bran puh-tehn-shuhl
Resting state rehs-tihng stat
Saltatory conduction sahl-tuh-tohr-e kuhn-duhck-shuhn Schwann cell shwahn sehl
Sensory nerve sehn-sar-e narv
Sensory neuron sehn-sar-e nar-ohn
Sensory receptor sehn-sar-e reh-sehpt-ar Sodium-potassium pump so-de-uhm puh-tahs-e-uhm puhmp
Soma som-uh
Somatic nervous system so-maht-ihck nar-vuhs sihs-tehm
Somatic reflex so-maht-ihck re-flehcks
Spinal nerve spi-nahl narv
Stretch reflex strehch re-flehcks
Sulcus (plural sulci) suhlck-uhs (plural suhlck-i)
Sympathetic ganglion chain sihm-pah-theht-ihck gahng-gle-uhn chan
Sympathetic nervous system sihm-pah-theht-ihck nar-vuhs sihs-tehm
Synapse sih-nahps
Synaptic cleft sih-nahp-tihck klehft
Synaptic end bulb sih-nahp-tihck ehnd buhlb
Synaptic knob sih-nahp-tihck nohb
Synaptic transmission sih-nahp-tihck trahnz- mihsh-uhn
Target tahr-giht
Telodendron tel-uh-dehn-drohn
Terminal bouton tar-muh-nuhl boo-tawn
Thalamus thahl-uh-muhs
Thoracolumbar system thohr-ah-ko-luhm-bahr sihs-tehm
Threshold threhsh-old
Threshold stimulus threhsh-old stihm-u-luhs
Ventral horn vehn-trahl hohrn
Ventral nerve root vehn-trahl narv root
Wave of depolarization wav of de-po-lar-uh-za-shuhn
White matter whit maht-ar
Withdrawal reflex wihth-draw-uhl re-flehcks
INTRODUCTION
An animal's body is enormously complex, whether we are talking about something small, like a 1-pound chinchilla, or something large, like a 2000-pound camel.
In order to maintain homeostasis, and therefore health, all those cells, tissues, organs, and systems have to be able to communicate with each other, and their functions have to be coordinated and controlled. Fortunately the body has two communication and control systems that help keep things working properly: the nervous system and the endocrine system. Both use chemicals to carry their messages, but they do it by different means, and on different timescales. The nervous system's chemical messengers are called neurotransmitters, and they are produced only by neurons (nerve cells). The neurotransmitters travel only very short distances, across spaces between nerve cells called synapses. This allows the system to react quickly, but the limited supplies of neurotransmitters in the cells do not allow it to sustain individual activities for long periods of time. The chemical messengers of the endocrine system, on the other hand, called hormones, are secreted directly into the bloodstream, where they travel comparatively long distances to reach their targets. The hormone targets, therefore, react more slowly to changes, but hormones can be secreted for long periods of time, so they can sustain individual activities for long periods of time. We discuss the endocrine system in Chapter 1 1. This chapter is about the nervous system.The nervous system is the rapid response, boss of bosses, communication and control system in the animal body. It monitors what's going on inside and outside the animal, and directs activities to maintain well-being. Understanding how the nervous system is organized and how it functions can help us appreciate what is going on in an animal that is anesthetized, intoxicated with a neurotoxin (poison affecting the nervous system), or unable to move properly because of trauma (hit by a car, intervertebral disc rupture, and so on).
Structurally the nervous system has two main divisions: the central nervous system (CNS) and the peripheral nervous system (PNS).
The central nervous system is composed of the brain and spinal cord, and the peripheral nervous system consists of cordlike nerves that link the central nervous system with the rest of the body.Functionally, the nervous system's activities fall into three main categories: (1) sensory functions, (2) integrating functions, and (3) motor functions. The nervous system senses changes from within the body or from outside the body and conveys this information to the spinal cord and brain. In the brain and spinal cord, the sensory information is received, analyzed, stored, and integrated to produce a response. A motor response instructs the body to do something, such as contract a muscle or cause a gland to secrete its product(s).
The branch of science that studies the nervous system is called neurology; neuro- refers to the nervous system, and logos means study of.
NEURONS AND SUPPORTING CELLS
Neurons (nerve cells) are the stars of the nervous system show. They are the basic, functional units of the system. That means they are the smallest pieces of the nervous system that show basic nervous system functions, such as responding to stimuli and conducting impulses from one part of the cell to another.
Like many Hollywood stars, neurons are high maintenance. They have a very high requirement for oxygen; they can't live without it for more than a few minutes. That is why cardiopulmonary resuscitation must be started within a few minutes of cardiac arrest. The heart may start beating again after that, but there could be brain damage if the neurons have been without oxygen for too long.
Shortly after an animal is born, its neurons lose their ability to reproduce, but they can regenerate cell processes if the cell body remains intact. The lack of reproductive ability is why serious nervous system injuries, such as strokes and spinal cord damage, are often so debilitating and have such long-lasting effects. Emerging research suggests that it may be possible to turn neurons' reproductive ability back on.
This holds great promise for the future of patients with neurologic deficits.Also, like Hollywood stars, neurons need a great supporting cast and crew to be successful. The neuroglia, or glial cells (from the Greek glia, meaning glue), structurally and functionally support and protect the neurons. They outnumber neurons about 10 to 1, but they are not directly involved in the transmission of information or impulses through the nervous system. Rather they are important parts of the infrastructure necessary for the neurons to do their jobs. The neurons are the stars of the nervous system show, and the glial cells are the supporting actors, tech crew, and minions that surround and support them.
Although neurons in different parts of the nervous system vary somewhat in appearance, their basic structure is the same (Figure 9-1). Structurally a neuron can be divided roughly into the central cell body, also called the soma or perikaryon, and the two different types of processes (extensions) from the cell body, called dendrites and axons.
Dendrites receive stimuli, or impulses, from other neurons and conduct this stimulation to the cell body. They can be referred to as afferent processes, because they conduct impulses toward the cell body (ad means “toward,” and ferre means “to carry”). Dendrites also may be modified into sensory receptors that receive, or sense, stimuli such as heat, cold, touch, pressure, stretch, or other physical changes from inside or outside the body. Dendrites tend to be short, numerous, and have many branches. (The word dendro is derived from the Greek word for branch because, when examined under a microscope, dendrites resemble the branches of a tree.)
The axon is the other type of process from the neuron cell body. Axons conduct nerve impulses away from the cell body
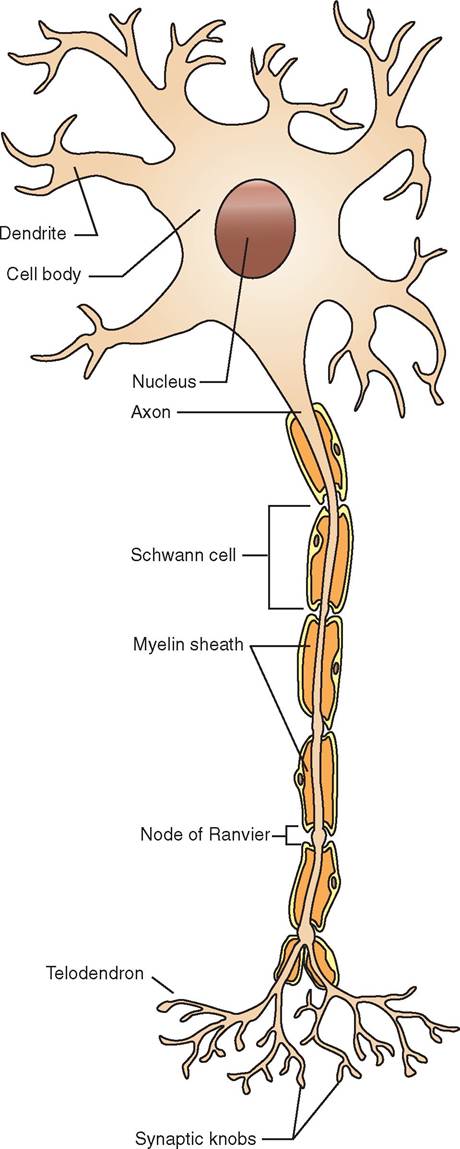
FIGURE 9-1 Structure of neuron.
toward another neuron or an effector cell (a cell that does something when stimulated, such as a muscle or gland cell).
They can be called efferent processes, because they conduct impulses away from the cell body (ex, “away”; ferre, “to carry”). In contrast to the short, numerous, branched dendrites, the axon is a single process that can be very long. Forexample, a single axon in the horse may extend for several feet—from the spinal cord all the way to the lower leg. Note: axons are sometimes referred to by another name, nerve fibers. When we're talking about the components of nerve cells, the term axon is usually used. When we're talking about the bundles of axons that make up cordlike nerves in the body, they are usually called nerve fibers. “Axons” and “nerve fibers” are two different names for the same thing. Think of them as aliases for each other.
Axons are often covered by a sheath of a fatty substance called myelin. Myelin grossly (without magnification) appears white. For that reason, nervous tissue containing many myelinated axons is often referred to as white matter. (Conversely, nervous tissue that is made up largely of neuron cell bodies appears darker and is called gray matter.) The myelin sheath is actually made of the cell membranes of specialized glial cells called oligodendrocytes in the brain and spinal cord, and Schwann cells in the nerves outside of the brain and spinal cord. These special glial cells wrap themselves around the axon like a thin pancake tightly wrapped around a hot dog. Because the axon of most neurons is fairly long, it takes multiple Schwann cells or oligodendrocytes lined up end to end to cover the entire length of the axon. Between adjacent glial cells are small gaps in the myelin sheath called nodes of Ranvier. The myelin sheath and nodes of Ranvier work together to enhance the speed of conduction of nerve impulses along the axon. Myelinated axons conduct nerve impulses faster than unmyelinated axons (Figures 9-2 and 9-3).
TEST YOURSELF 9-1
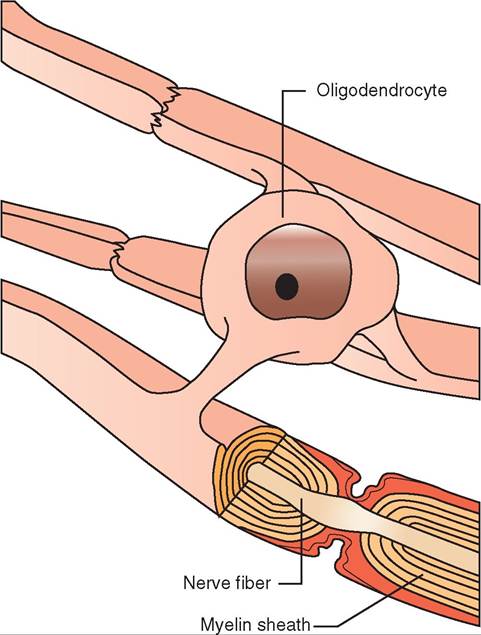
FIGURE 9-2 Oligodendrocyte, nerve fiber (axon), and myelin sheath. Oligodendrocytes wrap around nerve fibers in the central nervous system to form myelin sheaths.
1. How do basic communication and control functions differ between the nervous system and the endocrine system?
2. How are the functions of neurons and neuroglia different from each other?
3. Name the parts of a typical neuron.
4. How are the dendrites and axons different in structure and function?
5. What is the difference between gray matter and white matter?
6. What is the relationship between the myelin sheath and the nodes of Ranvier?
ORGANIZATION OF THE
NERVOUS SYSTEM
Many schemes are used to describe the anatomic or functional organization of the nervous system. Because the organizational terminology sets the foundation for our discussion about the nervous system, let's first look at how this terminology is used.
ANATOMIC LOCATION: CNS VERSUS PNS
A simple way to organize the nervous system anatomically is to think of it as being divided into two components: the
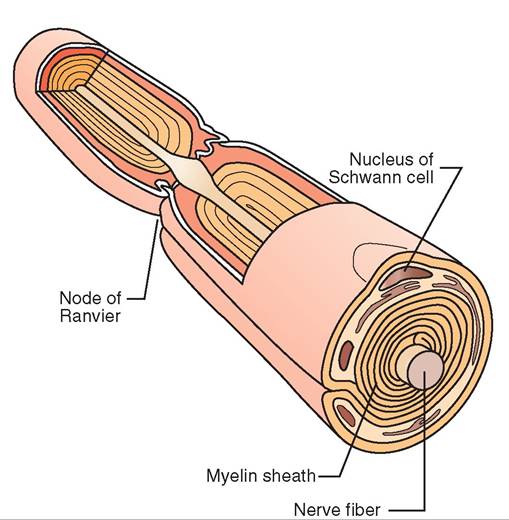
FIGURE 9-3 Myelin sheath. Schwann cells wrap around peripheral nerve fibers (axons) to form thick myelin sheaths.
central nervous system (CNS) and the peripheral nervous system (PNS). As the name implies, the CNS is anatomically composed of the brain and the spinal cord, which are found along the central axis of the body. Peripheral means “to the side” or “away from the center.” Therefore the peripheral nervous system (PNS) is made up of those components of the nervous system that extend away from the central axis outward, toward the periphery of the body. Cranial nerves are those few nerves of the PNS that originate directly from the brain. Most PNS nerves are spinal nerves that emerge from the spinal cord.
DIRECTION OF IMPULSES: AFFERENT VERSUS EFFERENT
Some nerve fibers conduct electrical impulses from the periphery toward the CNS, and other nerve fibers conduct impulses in the opposite direction, from the CNS toward the periphery. These two functional types of nerve fiber are called afferent nerve fibers and efferent nerve fibers. Afferent nerve fibers conduct nerve impulses toward the CNS, whereas efferent nerve fibers conduct nerve impulses away from the CNS.
Because afferent nerve fibers conduct sensations from the sensory receptors in the skin and other locations in the body to the CNS, afferent nerve fibers are usually called sensory nerve fibers. In contrast, efferent nerve fibers conduct impulses from the CNS out toward muscles and other organs. Because the efferent impulses are the ones that, among other things, cause skeletal muscle contraction and movement, efferent nerve fibers are usually called motor nerve fibers. The cranial and spinal nerves in the PNS and nerve tracts (bundles of axons) in the CNS may be made up of nerve fibers that are sensory or motor, or a combination of both. A nerve that contains only sensory nerve fibers is called a sensory nerve. A nerve that contains only motor nerve fibers is called a motor nerve. Nerves that contain both kinds of nerve fibers are called mixed nerves. Most nerves in the PNS are mixed nerves.
FUNCTION: AUTONOMIC VERSUS SOMATIC
When an animal turns its head in response to its name being called by its owner, efferent (outgoing) motor impulses from the brain are consciously sent to the muscles in the neck to turn the head toward the sound. This conscious, or voluntary, control of skeletal muscles is referred to as a somatic nervous system function. Because the action of the animal turning its head was caused by voluntary initiation of efferent impulses, this function would be classified as a somatic motor function. Impulses being sent to the CNS from receptors in the muscles, skin, eyes, or ears would be classified as somatic sensory functions, because they are consciously perceived by the brain.
In contrast to the voluntary movement of the somatic nervous system function, animals do not consciously have to think to contract their intestines, increase their heart rate in response to a threat, or stimulate release of digestive juices in response to ingestion of a meal. The animal also does not have to be consciously aware of blood pressure receptors informing the body that the blood pressure is too low or of stretch receptors indicating that the lungs have inflated. The part of the nervous system that controls and coordinates these automatic functions is called the autonomic nervous system (auto means “self,” and nomos means “law,” so the autonomic nervous system is the self-regulating system).
Like the somatic (voluntary) system, the autonomic system also has motor nerves and sensory nerves. However, instead of these motor nerves going to skeletal muscle to cause voluntary limb or body movement, the autonomic motor nerves send impulses to smooth muscle, cardiac muscle, and glands to regulate a wide variety of automatic body functions. Autonomic sensory nerves receive the afferent sensory impulses from sensory receptors that are used automatically to regulate these body functions.
TEST YOURSELF 9-2
1. What are the anatomic differences between the CNS and the PNS?
2. Which are afferent nerve fibers: motor nerve fibers or sensory nerve fibers? Which are efferent?
3. Identify each of the following as being controlled by the autonomic or the somatic nervous system and as being either sensory or motor:
Conscious movement of the forelimb
Slowing of the heart rate in response to an increased blood pressure
Constriction of blood vessels in the skin in response to cold temperatures
Perception of pain from an injection of antibiotics Perception of the amount of acidity present in the duodenum.
NEURON FUNCTION: DEPOLARIZATION AND REPOLARIZATION
It is often said that a nerve “fires,” or that an impulse is conducted from one end of a neuron to the other. What actually is occurring in the neuron when this happens? If we understand the concepts of depolarization and repolarization it is easier to understand how drugs such as local anesthetics can prevent nerves from firing or how imbalances of sodium or potassium in the body can adversely affect nerve function.
RESTING STATE, POLARIZATION, AND RESTING MEMBRANE POTENTIAL
When a neuron is not being stimulated, it is said to be in a resting state. However, even when the neuron is resting, it is still working to maintain its resting state. The cell membranes of neurons are electrically polarized at rest, like tiny charged batteries. Specialized molecules located in the neuron's cell membrane pump sodium ions (Na+) from the inside of the neuron to the outside and pump potassium ions (K+) from the outside to the inside. This specialized molecule is called the sodium-potassium pump (Figure 9-4).
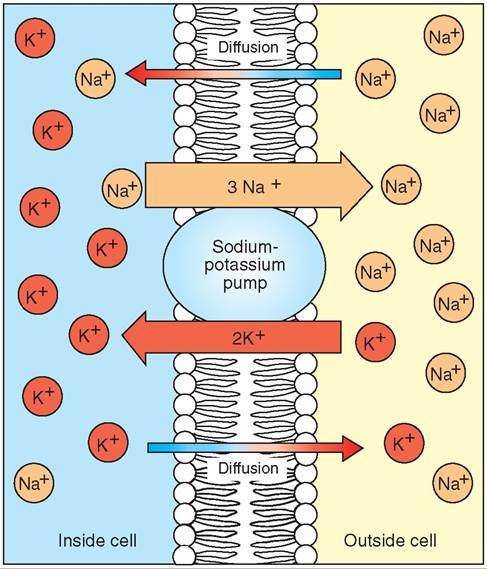
FIGURE 9-4 Sodium-potassium pump. This cellular mechanism is located in the cell membrane and actively pumps sodium ions (Na+) out of the neuron and potassium ions (K+) into the neuron.
Sodium (Na+) cannot readily diffuse or leak through the cell membrane on its own. Because sodium cannot diffuse across the membrane, the action of the sodium-potassium pump causes a higher concentration of sodium to accumulate outside the cell. The action of the sodium-potassium pump and the negative charges inside the cell cause a higher concentration of potassium to accumulate inside the cell. By keeping the sodium on one side of the membrane (outside) and the potassium on the other (inside), the cellular membrane separating the two is said to be polarized (because it has two distinct poles of ions on either side of the membrane).
The distribution of positive and negative charges from sodium, potassium, proteins, and other charged ions on either side of the neuronal membrane creates a difference in electrical charge across the membrane, with the inside of the neuron being more negatively charged than the outside. This electrical difference in charges across the membrane is called the resting membrane potential. The net negative resting membrane potential usually is stated as a certain negative number of millivolts (for example, -70 mV), indicating the net negative charge within the cell. By selectively pumping sodium out and potassium in, the sodium-potassium pump maintains this negatively charged, resting membrane potential—the cell membrane “battery” is charged (Figure 9-5, A).
DEPOLARIZATION
When an impulse from an adjoining neuron or from a specific type of external stimulus (such as heat, touch, or taste) stimulates a neuron, a set of specific steps occurs, resulting in the nerve “firing” or depolarizing. At the point where the stimulus occurs on the neuron, a specialized molecular structure on the neuron cell membrane called a sodium channel opens (Figure 9-5, B). This sodium channel allows only sodium ions (Na+) to pass through it. Because a higher concentration of sodium ions exists outside the cell than inside the cell, the sodium ions readily flow through the open sodium channels from the outside to the inside by passive diffusion. Not only is the sodium driven into the cell by the concentration gradient (the differences in concentration between the outside and inside), but the positive sodium (Na+) ions are attracted into the cell by the net negative charge inside the cell. Remember, opposite charges attract each other, and therefore the positive Na+ ions are attracted to the relative negative charge within the cell.
Depolarization refers to this opening of the sodium channels and the sudden influx of many sodium ions into the cell. It is called depolarization, because the sodium influx results in the loss of the two distinct poles of sodium and potassium on either side of the membrane. If we hooked an electric meter to the neuron, we would see the inside of the neuron go from a negatively charged resting membrane potential to a net positive charge during depolarization. This shift inside the cell from negative to positive makes sense when we consider the positive sodium ions flooding into the neuron. This significant change in electric charge from negative to positive is also referred to as an action potential.
REPOLARIZATION
Within a fraction of a second after sodium begins to flood into the cell during depolarization, the sodium channels snap shut, halting the influx. Almost simultaneously, specialized potassium channels open up in the cellular membrane (Figure 9-5, C). Analogous to the sodium channels, the potassium channels only allow potassium ions to pass through them.
With the potassium channels open, the potassium ions (K+) passively diffuse out of the cell, propelled by both the potassium concentration gradient (a high concentration inside and a lower concentration outside) and the strong positive charge brought into the cell by the influx of sodium ions. Remember that like charges repel, therefore the positive potassium ions are repelled by the relatively positive charge inside the neuron caused by the sodium influx. This outflow of potassium ions continues until these specialized potassium channels snap shut a split second after they have opened (Figure 9-5, D). Because the potassium ions (K+) are positive, the exodus of potassium ions from the neuron causes the charge inside the cell to swing back in the negative direction.
This change of the cell's charge back toward the net negative resting membrane potential is called repolarization. The cell is said to be repolarized, because the sodium and potassium ions are once again on opposite sides (opposite poles) of the cell membrane. The only difference between the end of the repolarization phase and the resting state is that
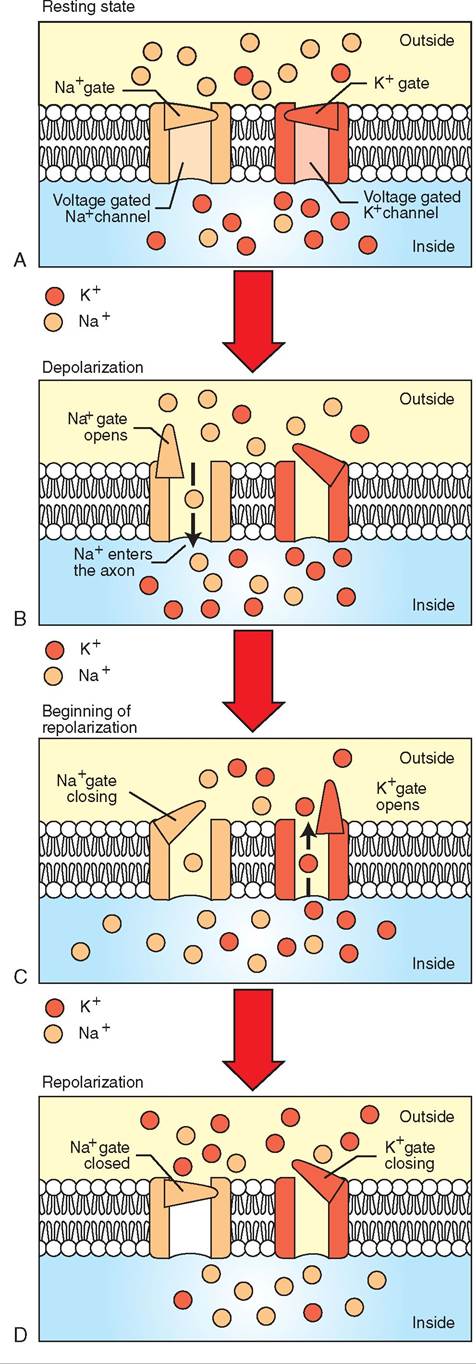
FIGURE 9-5 Depolarization and repolarization. A, Resting state. Sodium has been pumped out of the cell and potassium has been pumped in, producing a net negative electrical charge inside the cell membrane compared to the outside. B, Depolarization. A stimulus has caused the gate on the sodium channel to open, allowing sodium ions to flow into the cell. This produces a net negative charge on the outside of the cell membrane—the opposite of the resting state. C, Beginning of repolarization. The gate on the sodium channel is closing and the gate on the potassium channel is opening to allow potassium ions to flow out of the cell. D, Repolarization. A sufficient outflow of potassium ions has restored the net negative charge to the inside of the cell, but the sodium and potassium ions are on opposite sides of the cell membrane from where they started. Therefore, sodium ions are pumped out of the cell and potassium ions are pumped into the cell. The regain of the resting state is shown in Figure 9-4.
the sodium and potassium ions are on the opposite sides from where they began. To restore the sodium and potassium to their original locations on either side of the membrane, the sodium-potassium pump quickly moves the misplaced sodium and potassium ions back to their original sides (see Figure 9-4).
Note: It may seem as though the acts of depolarization and repolarization must involve a molecular tidal wave of ions moving back and forth. The truth is that relatively few sodium and potassium ions move at each point of the depolarization-repolarization cycle, which explains why the cycle is completed rapidly and why even minor imbalances in sodium or potassium in the body can greatly affect normal neuron function.
DEPOLARIZATION THRESHOLD, NERVE IMPULSE CONDUCTION, AND ALL-OR-NOTHING PRINCIPLE
Not every depolarization stimulus results in the complete depolarization-repolarization cycle. The initial stimulus must be sufficient to make the neuron respond. When the stimulus is strong enough to cause complete depolarization, it is said to have reached the threshold, and this causes the cell to depolarize or “fire.” A stimulus of sufficient intensity to generate a nerve impulse is called a threshold stimulus.
To understand how this works, let's use an example in which a neuron with sensory receptors on its dendrites receives a very weak stimulus. The weak stimulus results in only a few sodium channels opening and therefore only a small influx of sodium ions into the neuronal cell. Because of this, we would only see a slight positive change in neuron charge from the resting membrane potential. Because the charge was not very significant, the cell did not reach the threshold, and the few sodium channels that opened would close without causing further effect on other sodium channels. The sodium-potassium pump would quickly move the few displaced sodium ions from inside back to the outside, and the neuron would go back to its resting state. In this case, the stimulus failed to depolarize the neuron, and the information from the sensory receptors in the dendrites was not transmitted to the brain.
If, however, the stimulus on these sensory receptors had been larger, more sodium channels would have opened, and a larger number of sodium ions would have entered the neuronal cell. This would have produced a significant positive change in the membrane potential in the immediate area of the cellular membrane. If the change had been sufficient to reach the threshold, sodium channels adjacent to this area would also open. This would allow sufficient sodium influx into these adjacent areas to reach the threshold, causing further adjacent sodium channels to open. In other words, the initial stimulus would cause a spreading wave of opening sodium channels to travel along the cell membrane of the entire neuron. This wave of sodium channels opening to allow sodium influx is called the wave of depolarization. As you recall, the strong influx of sodium ions during depolarization was called the action potential; therefore this wave of depolarization can also be called conduction of the action potential. In clinical terms, however, this wave of depolarization or conduction of the action potential along the cell membrane is most commonly called a nerve impulse.
In the simplest terms, a nerve impulse is conducted along a nerve fiber by the “flipping” of the electric charges across the cell membrane (depolarization), followed quickly by the “unflipping” of the electric charges (repolarization). That process stimulates the adjacent area of the cell membrane in the direction of impulse conduction (remember that dendrites conduct impulses toward the cell body, and axons conduct impulses away from the cell body) to flip and unflip, which stimulates the adjacent area, and so on. If we were very tiny and could watch the cell membrane in one area of the neuron while a nerve impulse was being conducted, we could watch the flipping and unflipping of charges happen as the impulse passed by us.
Regardless of how strong the initial stimulus was, if it were sufficient to achieve the threshold for a neuron to fire (depolarize), the nerve impulse (action potential) would be generated and conducted along the entire neuron with uniform strength. This phenomenon is called the all-or- nothing principle, because either the complete neuron depolarizes to its maximum strength, or it does not depolarize at all. This is an important concept! A nerve impulse is a nerve impulse is a nerve impulse. They are all basically the same. What makes one nerve impulse signify the color red, another represent a particular odor, and a third cause a muscle fiber to contract? It depends on where the impulse is going. Sensory (afferent) nerve impulses go to particular areas of the brain, where they are interpreted as the appropriate sensation. Motor (efferent) nerve impulses go to effector organs, which are stimulated to perform particular actions.
REFRACTORY PERIOD
For a very brief period during and after a neuron has generated a nerve impulse, it cannot generate another impulse. This is called the neuron's refractory period. If a second threshold stimulus arrives at the dendrites or on the neuron cell body while the sodium channels are open or while the potassium molecules are moving rapidly through their open channels, the stimulus will not cause a second depolarization. Because cells in the depolarization and early repolarization phases are already in the process of executing the depolarization-repolarization cycle (firing), they can't depolarize (fire) again until the cycle is completed. Thus, any stimulus arriving at that point in the depolarization- repolarization cycle would die out. The neuron is said to be in a refractory period because it is refractory or “insensitive” to new stimuli until it recovers from the previous nerve impulse.
The period of sodium influx and early potassium outflow is a part of the refractory period during which no stimulus, no matter how strong, can cause the cell to depolarize again. This period is called the absolute refractory period because the cell absolutely cannot respond. However, if a very strong stimulus comes during the tail end of the time the membrane is repolarizing and restoring the resting membrane potential, it may be possible to stimulate another depolarization. Therefore, during this part of the refractory period, the cell may depolarize again if the stimulus is much stronger than normal. This period is called the relative refractory period, because the cell is still refractory to stimuli of normal intensity but may respond to very strong stimuli.
HOW MYELINATED AXONS CONDUCT ACTION POTENTIALS QUICKER: SALTATORY CONDUCTION
If all neurons sent their wave of depolarization or their conduction of action potentials from one sodium channel to the next in a series of tiny steps, the transmission of the nerve impulse from one end of a neuron to the other would be relatively slow. Think of the sodium channels as a set of tiny dominos set up in a line several feet long. When we tip over the first domino, we know that it is going to take some time for the last domino to fall. The same thing happens if each sodium channel opening stimulates the opening of the adjacent channel.
If, however, our domino line tipped over 10 dominoes at a time, the speed at which we would reach the end of the domino line would be greatly accelerated. In neurons with axons wrapped in a myelin sheath, a similar effect happens with the depolarization wave. Like a rubber coating on an electrical wire that prevents electrical shorts, the myelin sheath prevents sodium ions from flowing across the neuronal cell membrane. Therefore depolarization in myelinated axons can only take place at the gaps in the myelin sheath that occur at the nodes of Ranvier. Thus, when the sodium influx at one node is sufficient to open adjacent sodium channels, the next available sodium channel is at the next node of Ranvier. The depolarization wave in the myelinated axon skips from one node of Ranvier to the next, greatly accelerating the rate at which the depolarization wave moves from the neuron cell body to the other end of the axon. This rapid means of conducting an action potential is called saltatory conduction (the word saltatory is derived from the Latin saltare, which means “to leap”) (Figure 9-6).
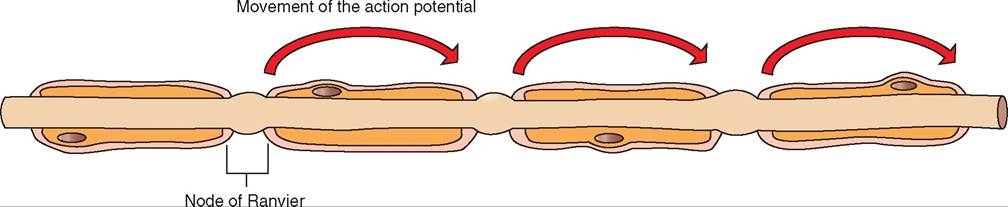
FIGURE 9-6 Saltatory conduction. A nerve impulse jumps from one node of Ranvier to the next, producing rapid conduction of the nerve impulse.
∕j CLINICAL APPLICATION
Local Anesthetics
Local anesthetics are drugs that are injected into superficial areas of the body to block the conduction of sensations from that area. You may have experienced this form of anesthesia if your dentist administered a local anesthetic to numb an area of your mouth. Local anesthetic drugs such as lidocaine prevent sensory nerves from depolarizing despite stimulation from the dental or surgical procedure. If these sensory nerves do not depolarize, the brain is unaware of any sensation from that area of the body, therefore you do not feel pain. Lidocaine prevents the sensory neuron from depolarizing by blocking the sodium channels through which sodium ions usually flood into the neuronal cell. If the sodium channels are plugged by the local anesthetic molecule, no sodium can flood into the cell despite the channels being stimulated to open. No sodium influx means that no positive charge occurs in the neuron, threshold is not attained, and the stimulus is not turned into a depolarization wave. Any nerve impulse that has been generated stops at that point.
Anesthesia means without sensitivity. If a sensory nerve does not depolarize, the animal’s brain does not perceive sensations from that area of the body. Local anesthetics are used not only to anesthetize areas of the body for minor surgical procedures, but also to aid in identifying sources of pain that cause lameness in horses. In a lame horse, a local anesthetic may be injected around selected sensory nerves to prevent them from transmitting impulses. If injection around a particular nerve improves the horse’s movement or reduces the lameness, the veterinarian knows that the source of the problem is in the area of the leg or hoof whose sensations are supplied by the “blocked” nerve. If the “nerve block” does not improve the lameness, the veterinarian injects another specific sensory nerve and repeats the process until the horse appears to have less pain.
The rapid conduction of impulses along myelinated neurons by saltatory conduction makes processes such as vision and fine motor control possible in larger animals such as humans and many domestic animal species. The importance of the myelin sheath and saltatory conduction to normal functioning can be illustrated by the symptoms of demyelinating diseases (diseases that damage or destroy myelin) such as multiple sclerosis
HOW NEURONS COMMUNICATE:
THE SYNAPSE
Once the nerve impulse, or action potential, has been successfully conducted to the end of the axon, it must be transmitted to the next neuron or to the cells of the target organ or tissue. Because two adjacent neurons do not physically touch each other, this process cannot be accomplished by directly continuing the depolarization wave. Instead, the neuron must release a chemical that stimulates the next neuron or cell. This perpetuation of the nerve impulse from one neuron to the next cell is called synaptic transmission.
The synapse is the junction between two neurons or a neuron and a target cell. The synapse consists of a physical gap between the two cells called the synaptic cleft. The neuron bringing the nerve impulse to the synapse and releasing the chemical to stimulate the next cell is called the pre- synaptic neuron. The chemical released by the presynaptic neuron is called the neurotransmitter, and the neuron that contains the receptors that receive the neurotransmitter is the postsynaptic neuron (Figure 9-7).
If we look closely at the end of the axon on the presyn- aptic neuron, we see a branched structure called the telodendron. Each branch of the telodendron ends in a slightly enlarged bulb called the terminal bouton (bouton meaning “button”), synaptic end bulb, or synaptic knob. The synaptic knobs contain many mitochondria that provide energy for the processes that occur there, and also many vesicles (small sacs) that contain the neurotransmitter. When the axon’s wave of depolarization reaches the synaptic knob, calcium channels open in the knob’s cellular membrane, resulting in an influx of calcium into the synaptic knob. This influx of calcium causes the vesicles containing neurotransmitters to fuse with the knob’s cellular membrane and dump their contents into the synaptic cleft. These neurotransmitters diffuse rapidly across the tiny synaptic cleft toward the postsynaptic membrane.
∕ j CLINICAL APPLICATION
Multiple Sclerosis
Multiple sclerosis, also known as MS, is a disease of humans that results in damage to the myelin sheaths of nerve fibers in the brain and spinal cord. Nerve fibers whose myelin sheaths eheanve b damaged conduct impulses abnormally, or not at all. Because both sensory and motor nerve fibers can be affected, the clinical signs of MS can be sensory and/or motor. Sensory effects include tingling, numbness, visual problems, iaensd difficult with coordination and balance. Motor effects include muscle weakness, muscle spasms, difficulty moving, and problems with speech and swallowing. The exact cause of MS is not known, but it is believed to be caused, at least in part, by the person’s own immune system attacking the sntermvo. us sy
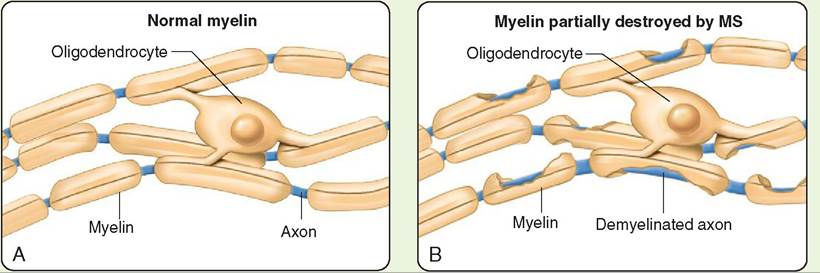
Effects of multiple sclerosis (MS). A, A normal myelin sheath allows rapid conduction. B, In MS, the myelin sheath is damaged, disrupting nerve conduction. (From Thibodeau G, Patton K: Structure and function of the body, ed 14, St Louis, 2012, Mosby.)
/ TEST YOURSELF 9-3
1. During depolarize tion, whatio n channels open and what ion moves? Where Sees it move?
2. During repolahzation, whaleon channels open and whrt ion moves? Where 1oes it move?
3. Whzt normally rr^r^inteinntner^^fr^gmcoe^^^^>oten- tizl of a neuron Suring the resting state?
4. What isthrehhold? What roledoeS tCrrjsh^ld play in the all-or-none principle?
5. What estoe differanen benween the absolute anS the relative refractory cerioSs?
6. Explain why waves of Secolarization are conSucteS faster in myelinateS axons than in une∣selinateS ones.
On the postsynaptic membrane are specialized proteins called receptors. Tlir neurotransmitter molecules released by the synaptic knob bind to these receptors and trigger a change in the postsynaptic cell. However, the postsynaptic membrane receptors are very specific about which neurotransmitters they will bind. If the neurotransmitter and reeceptor ar not matched, they will not bind to each other, and no change will be triggered in the postsynaptic cell. An analogy that illustrates this concept is that of a lock and key. Only c^Maiιι keys (types of neurotransmitter) will fit in a lock (the receptor) and cause the lock to open (the receptor to tπ'rger cellular changes). Thus, synaptic transmission is only effective if receptors to the neurotransmitter exist on the yostsycaptic cell’s membrane.
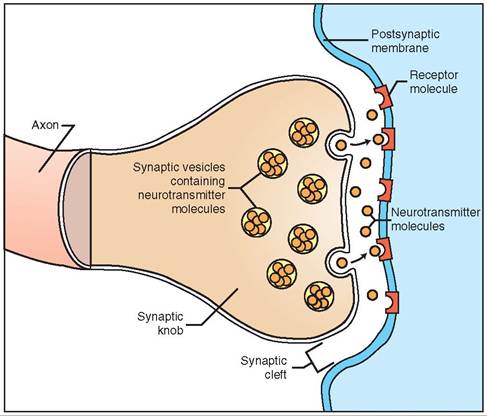
FIGURE 9-7 Chemical synapse. Neurotransmitters are released into the synaptic space, where they combine with receptors on the postsynaptic membrane (another neuron or an effector cell).
TYPES OF NEUROTRANSMITTER AND THEIR EFFECT ON POSTSYNAPTIC MEMBRANES
Many different types of neurotransmitter are associated with synapses in both the CNS and PNS. Generally, we can classify these neurotransmitters into two categories: excitatory neurotransmitters and inhibitory neurotransmitters. A their name implies, excitatory neurotransmitters Iiwe an excitatory effect on the postsynaptic membrane when they combine with their specific receptors. Specifically, excitatory neurotransmitters usually cause an influx of sodium so that the postsynaptic membrane moves toward threshold. If the postsynaptic membrane is stimulated sufficiently by enough excitatory neurotransmitter, then threshold will be reached and depolarination of the postsynaptic membrane will occur, beginning a new nerve impulse.
on car^ist to excitatory neurotransmitters, inhibitory neurotransmitters tend to hyperpolarize the postsynaptic membrane, making the inside of the cell more negative ifnstead o positive and moving the charge within the postsynaptic cell farther away from threshold. When inhibitory neurotransmitters combine with their specific recep- tnors o the postsynaptic side, they may cause chloride rchannels o potassium channels to open up on the post- synaptic membrane. This allows the negatively charged chloride ions (Cl-) tr enter the postsynaptic cell and allows potassium (K+) ions to leave the cell, making tehe insid of the cell more negatively charged (a change in charge that is opposite from that needed to reach threshold).
Neurotransmitters usually can be classified as excitatory iotro riynhib based on the effect they have on the postsyn-
aeolpml.tiec c S neurotransmitters, however, can have an excitatory effect on some cells and an inhibitory effect on others, so it is difficult in most cases to make sweeping statements about whether a given neurotransmitter is one ro.r the othe
Acetylcholine is one of the most commonly studied neurotransmitters in the body. It can be either an excitatory or inhibitory neurotransmitter depending on its location in the body. A the junction between somatic motor neurons aunscdletshe m they supply, acetylcholine is an excitatory neurotransmitter that stimulates muscle fibers to contract. However, at the site where parasympathetic nerns synapse with the heart, acetylcholine has an inhibitory effect that slows the heart rate.
Norepinephrine, dopamine, and epinephrine are all neurotransmitters that belong to a group called catecholamines. Norepinephrine is associated with arousal and fight-or- flight reactions of the sympathetic nervous system. Epinephrine is rel^^ed primarily from the adrenal medulla (center of the adrenal gland) and therefore plays more of a role as a hormone in the fight-or-flight reactions of the sympathetic nervous system. Dopamine is found in the brain, where it is itnhvolved w autonomic functions and muscle control. iHthumans w a decreased number of functioning dopamine nweurons sho the muscle tremors and shaky gait associated with !-'onk^^^^irfs disease.
Gamma-aminobutyric acid (GABA) and glycine are two neurotransmitters that are inhibitory. GABA is found in the brain, and glycine is found in the spinal cord. Some groups of tranquilizers, such as diazepam (Valium), work by increas- iAng the GAB effect on the brain, thus inhibiting activity in the brain and producing tranquiliuation (reduced anxiety) with sedation (drowsiness).
One oottsgnaptic membrane may have multiple types of presynaptic neurons across the synaptic cleft. For example, a postsynaptic motor neuron in the brain may have some pre- suyrnoanpstic ne that release the excitatory neurotransmit- tcertyalcholine into the synaptic cleft and other presynaptic
neurons that release the inhibitory neurotransmitter GABA into that same synaptic cleft. Depending on which set of neurons is more active—the excitatory, acetylcholine- relensing neurons or the neurons releasing the GABA inhibi- tuorroytrnaensmitter—the postsynaptic motor neuron may iemithuelartbed st or inhibited.
By having both inhibitory and excitatory neurotransmit- trevrosu, sthe ne system can selectively increase or decrease
Clinical application
Poisons That Affect the Nervous System
Every rar, animals are injured or killed by nervous system poisons in the form of insecticides (flea products, bug sprays, agricultural chemicals), rodenticides (mice and rat killers), poisonous plants, or other chemical poisons that disrupt the function of the nerve synapse. Many of these poisons act by combining with or blocking the neurotransmitter receptors on the postsynaptic membranes. To ictohmbine w the receptors, these poisons must have a similar molecular structure to the natural neurotransmit- toedrys.in the b
fMany o these poisons bind to the receptors just like the natural neunofransmitters do, thereby stimulating the post- synaptic cell or neuron. In these cases of poisoning, we see avenrsotimulation of some aspect of the nervous system or itshseuets innervated by that part of the nervous system. Animals my∙ show signs of seizures or muscle tremors, indi- icmatuinlagtisotn of the somatic motor system or over
stimulation of the autonomic nervous system, resulting in rvomiting o changes in respiration, heart rate, or other autonomic functions.
Imneso cases, the poison can combine with the receptor, but it does not pdocture an effect. Io this case, the poison wou 1d prevent the natural oensolsaoemittes from combining with the receptor to produce its normal effect. Because the poison acts as a blocker of that receptor, we ewould se a suppression of that part of the nervous system. A Jcssi'c example of this effect is curare, the nerve poison fnound o the skin of the brightly colored poison dart frogs in South America. Curare combines with the recep- tenolertsalo sk muscles and prevents the presynaptic neuront nevroternsmitter from stimulating the muscle to contract; thus, curare paralyzes the animal's muscles. South American natives use this toxin to coat the tips of their hunting darts, arrows or spears to paralyze their quarry. In tyh, is wa the animals are easily captured, even if the wound itself is not fatal.
We use a similar effect medically to paralyze the normal respiratory movements of animals during open chest surgery. By paralyzing the respiratory muscles, we can more easily mechanically ventilate, or breathe for, an animal without Hdhting its body’s own contraction of the diaphragm and rib cuasgcleesm.
the activity of specific parts of the brain or spinal cord. Drugs or poisons that imitate inhibitory or excitatory neurotransmitters will cause CNS depression or increased CNS activity, respectively. For example, ivermectin, a commonly used antiparasitic drug (it kills parasites), causes an increased inhibitory neurotransmitter effect. In animals receiving an overdose of ivermectin, the main clinical signs are severe depression, loss of normal control of voluntary movements, and coma, all of which reflect inhibition of the neuronal activity in the brain.
STOPPING AND RECYCLING THE NEUROTRANSMITTER
If the neurotransmitter were released, combined with its corresponding receptor, and allowed to remain in the synapse or on the postsynaptic receptor, the postsynaptic cell would either continue to be excited or continue to be inhibited depending on the type of receptor being stimulated. Therefore the body must have a way of stopping the effect of the neurotransmitter quickly so that this does not occur.
In the case of acetylcholine, the neurotransmitter is broken down quickly by an enzyme found on the postsyn- aptic membrane called acetylcholinesterase. The -ase suffix tells us that this compound is an enzyme that acts on acetylcholine. The broken-down components of acetylcholine are reabsorbed by the synaptic knob, reassembled into new acetylcholine molecules, and repackaged into vesicles for release with the next wave of depolarization. If acetylcholinesterase is prevented from working, acetylcholine will not be broken down and acetylcholine receptors will continue to be stimulated. This is what happens when animals are exposed to poisonous levels of flea products containing organophosphate insecticides. In organophosphate poisoning, the insecticide combines with the acetylcholinesterase and inactivates it. The small amount of acetylcholine normally released by presynaptic neurons causes overstimulation of acetylcholine receptors, resulting in diarrhea, vomiting, difficulty in breathing, and constricted pupils.
After the release of norepinephrine from the presynap- tic neuron, the norepinephrine is rapidly taken back into the synaptic knob, where it is broken down into its components by the enzyme monoamine oxidase (MAO). Any norepinephrine not reabsorbed by the synaptic knob is degraded by another enzyme called catechol-O-methyl transferase (COMT). Compared with acetylcholinesterase, the activity of MAO and COMT is relatively slow, which helps to explain why effects of these excitatory neurotransmitters can linger for a while after their release. One of the mechanisms by which some human antidepression medications work is by blocking MAO or COMT, which allows norepinephrine to prolong its excitatory effect on the brain.
Because drugs and poisons encountered in veterinary medicine often produce their effects by increasing or decreasing excitatory or inhibitory neurotransmitter effects or by affecting the enzymes that terminate these effects, it’s important to understand the concepts of synaptic function and neurotransmitter release and termination.
TEST YOURSELF 9-4
1. What role do the synaptic cleft, presynaptic neuron, neurotransmitter, and postsynaptic neuron play in the continuation of a depolarization wave from one nerve to another?
2. What is the functional relationship between a neurotransmitter and a receptor? Will any neurotransmitter stimulate any receptor?
3. What is the difference between an excitatory and an inhibitory neurotransmitter?
4. How is acetylcholine different from acetylcholinesterase?
5. What are catecholamines?
6. What are GABA and glycine?
THE CENTRAL NERVOUS SYSTEM-BRAIN AND SPINAL CORD
To the unaided eye, the central nervous system (CNS) consists of the brain and the spinal cord. At the microscopic level, however, the main components of the CNS are neuron cell bodies, myelinated and uynmyelinated nerve fibers, and glial cells. The gray matter of the CNS contains most of the neuron cell bodies, and appears a dark brownish-gray color grossly. It is usually thought of as the “thinking” part of the CNS. The white matter contains most of the myelinated nerve fibers and appears white because of all the myelin. It is the “wiring” that connects the various components of the brain. It has been estimated that the white matter of the human brain makes up a network of some 100,000 miles of nerve fibers. (See “Secrets of the Brain” in the February 2014 issue of National Geographic.)
Neurologic disease or disorders affecting the brain produce clinical signs that sometimes can be identified as belonging to specific areas of the brain. Although knowing all the centers or nuclei (clusters of neurons within the CNS) of the brain is not essential, the veterinary technician should be familiar with what the main parts of the brain do, to better understand the effect of neurologic disease and medications that affect the CNS (Figure 9-8).
In general, we can think of the brain as being divided into four different sections: the cerebrum, cerebellum, diencephalon, and brainstem. Each section of the brain has its own particular functions. The brainstem and diencephalon are the more primitive parts of the brain; the cerebellum coordinates motor control; and the centers of higher learning and intelligence are found in the cerebrum. Therefore disease in each part of the brain produces different clinical signs.
CEREBRUM
The cerebrum is made up of gray matter in the cerebral cortex (the outer-most superficial layer of the brain) and white matter beneath the cortex, including the corpus
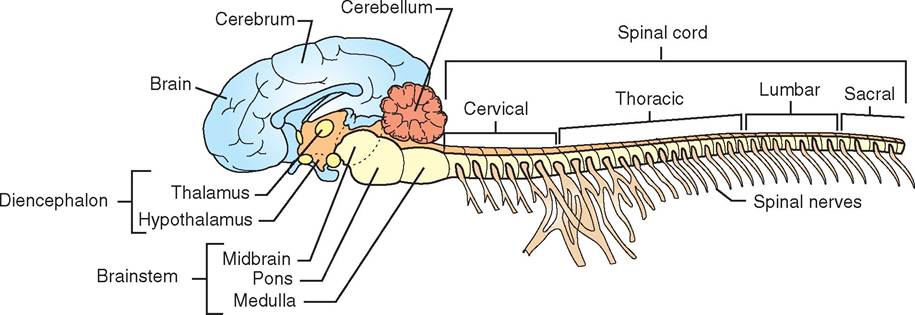
FIGURE 9-8 Anatomy of the central nervous system.
callosum (a set of fibers that connects the two halves of the cerebral cortex). The cerebrum is the largest part of the brain in domestic animals and constitutes the area of the brain responsible for those functions most commonly associated with higher-order behaviors, such as learning, reasoning, and intelligence. The cerebrum receives and interprets sensory information, initiates conscious (voluntary) nerve impulses to skeletal muscles, and integrates neuron activity that is normally associated with communication, expression of emotional responses, learning, memory and recall, and other behaviors associated with conscious activity.
The surface of the cerebrum is covered by “wrinkles” which serve to increase the area of the cerebral cortex, therefore making more room for gray matter. The wrinkled appearance is made up of folds called gyri (plural of gyrus) separated by deep grooves called fissures and more shallow grooves called sulci (plural of sulcus). The most prominent groove is the longitudinal fissure, which divides the cerebrum into right and left cerebral hemispheres (Figure 9-9). Each hemisphere is divided by sulci into lobes. Different lobes of the cerebral hemispheres specialize in certain functions. For example, a section of lobes in the front half of the brain contains the organized areas that initiate voluntary motor functions, whereas the lobe immediately posterior to this section contains the area that identifies the locations of sensations in or on the body.
If neurons of certain lobes of the cerebrum begin to fire spontaneously as a result of drugs, cellular damage, or neurotransmitter imbalance, the animal can exhibit spontaneous movements, seizure activity, abnormal behaviors, or hallucinations, depending on which lobes are affected. If parts of the cerebrum become damaged and nonfunctional from lack of oxygen, poisonous substances, or blood clots (strokes), the animal may lose the perception of specific sensations, may experience loss of voluntary movement, or may be unable to retain or recall information (unable to learn).
CEREBELLUM
The cerebellum, located just caudal to the cerebrum, is the second largest component of the brain. It also has a
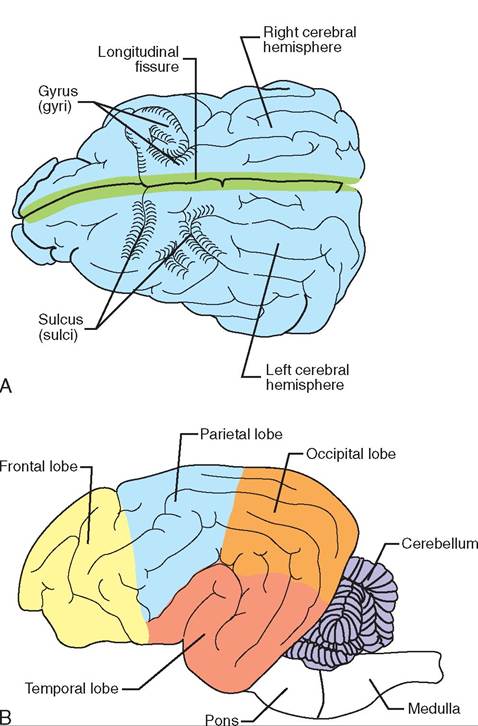
FIGURE 9-9 Sulci and gyri of the cerebrum. A, Top view. B, Side view.
wrinkled-appearing surface, and a gray matter cortex with white matter beneath it. The cerebellum allows the body to have coordinated movement, balance, posture, and complex reflexes. Essentially, the cerebellum compares the movement the body intends to make with the actual position of muscles and joints to determine whether the intentions of the cerebral cortex are actually being carried out. If the movements are not being carried out accurately, the cerebellum will stimulate or inhibit muscles to fine-tune the movements.
For example, if you flex your arm, you can feel your biceps muscle contract while the opposing triceps muscle relaxes. When your arm begins to flex, stretch receptors associated with the muscles send feedback to the cerebellum to keep it informed of the position of the arm. The cerebellum then sends impulses to both the cerebral cortex and the muscles involved in the arm movement so that adjustments in the contraction can be made. In addition to making the voluntary body movements smooth and accurate, the cerebellum also uses this same sensory feedback from the muscles to maintain posture and balance.
Damage or disease involving the cerebellum results in hypermetria, a condition in which voluntary movements become jerky and exaggerated. A condition like this occurs in pigs with cerebellar disease and causes the affected animals to exhibit a goose-step gait in which the lifting and placing of the foot become exaggerated. Similar abnormal gaits can be seen in the young of other species born with an incompletely developed cerebellum or in animals with viral or bacterial disease that affects the cerebellum.
DIENCEPHALON
The diencephalon is not as physically defined as the cerebrum and cerebellum. It does not have clearly visible layers of gray matter and white matter. It serves as a nervous system passageway between the primitive brainstem and the cerebrum. That gives rise to its common name: the between brain. Although many structures are associated with the diencephalon, veterinary technicians need to be familiar with three major ones:
1. The thalamus acts as a relay station for regulating sensory inputs to the cerebrum.
2. The hypothalamus is an interface between the nervous system and the endocrine system.
3. The pituitary is the endocrine “master gland” that regulates production and release of hormones throughout the body.
The hypothalamus also plays major roles in temperature regulation, hunger, thirst, and components of rage and anger responses. Disease or drugs that result in fever (hyperthermia) or compulsive eating or drinking often involve centers within the hypothalamus.
BRAINSTEM
The brainstem is the connection between the rest of the brain and the spinal cord. Its name comes from its appearance as a stem on which the other parts of the brain (the cerebrum, cerebellum, and diencephalon) sit. It is the most primitive part of the brain and is composed of the medulla oblongata, the pons, and the midbrain. Like the diencephalon, the brainstem does not have clearly visible layers of gray matter and white matter.
The brainstem's role is to maintain basic support functions of the body, so it operates at the subconscious level. It is heavily involved in autonomic control functions related to the heart, respiration (including coughing, sneezing, and hiccupping), blood vessel diameter (vasomotor control), swallowing, and vomiting. Many of the cranial nerves (see later text) originate from this area of the brain. Because functions related to the heart, blood vessel diameter, and respiration are critical to life, damage to the brainstem can result in the animal dying rapidly from respiratory failure or cardiovascular collapse. Fortunately, the brainstem is well protected by the skull, so brainstem injury rarely occurs unless there is major damage to the skull.
TEST YOURSELF 9-5
1. What part of the brain is responsible for conscious thought and perception of sensations?
2. What are the correct names for the bumps and fissures that make the cerebral cortex appear wrinkled?
3. What part of the brain is critical for coordination, posture, and fine motor control? How does this part of the brain accomplish these responsibilities?
4. What part of the brain serves as a relay station for impulses going to and from the cerebrum?
5. Which part of the brain controls many autonomic functions related to cardiovascular, respiratory, and gastrointestinal functions?
OTHER CLINICALLY IMPORTANT STRUCTURES OF THE BRAIN
Other structures in the brain are important for the veterinary technician to be aware of because of their role in how drugs or disease affect the brain or their role in diagnostic procedures used in veterinary medicine.
Meninges
The meninges are a set of connective tissue layers that surround the brain and spinal cord (Figure 9-10). The three
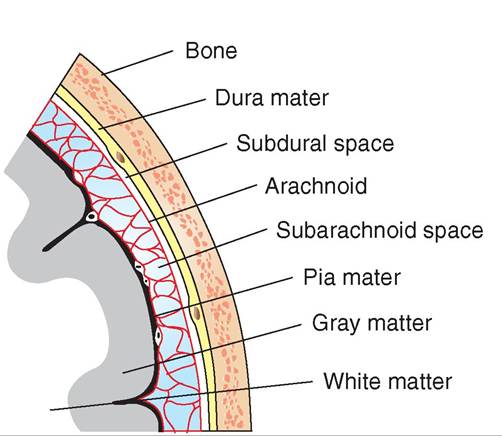
FIGURE 9-10 The meninges. (Adpated from Gilbert S: Pictorial anatomy of the cat, Seattle, 1975, University of Washington Press.)
layers of the meninges—from outside to the innermost layer—are the tough, fibrous dura mater, the delicate, spiderweb-like arachnoid, and the very thin pia mater, which lies directly on the surface of the brain and spinal cord. These connective tissue layers contain a rich network of blood vessels that supply nutrients and oxygen to the superficial tissues of the brain and spinal cord. The fluid, fat, and connective tissue found between the layers of the meninges also provide some cushioning and distribution of nutrients for the CNS. Inflammation of these meningeal membranes resulting from viral or bacterial infections is called meningitis.
CEREBROSPINAL FLUID
rTahine b and spinal cord are bathed and protected from the uard inner surfaces of the skull and spinal column by a fluid called the cerebrospinal fluid (CSF). The clear, slippery CSF ceitrwceuelnates b layers of the meninges and through cavities (canals and ventricles) inside the brain (ventricles) and the spinal cord (central canal). In addition to its cushioning function, the CSF's chemical composition may be involved iengutlhaetiorn of certain autonomic functions, such as
respiration and vomiting. For example, if the pH of the CSF becomes more acidic, the respiratory center in the brainstem ewaislel incr the respiratory rate.
Because the CSF circulates throughout the CNS, infection, inflammation, or cancer in the brain or spinal cord can cause changes in the amount of protein contained in the CSF; they can also cause changes in the composition of the CSF cel 1s, including white blood cells or cancer cells. Veterinarians can diagnose certain nervous system diseases or cancers by taking a sample of the CSF, called a CSF tap, and examining it for particular types of cells or for specific cohmapnogseistiionnc.
BLOOD-BRAIN BARRIER
The blood-brain barrier is a functional barrier separating ltahreiecsapil in the brain from the nervous tissue itself. Unlike other capillaries in the body that have small openings between the cells of the capillary walls, the cells that make luaprythe capil walls in the brain are aligned tightly together without these openings, or fenestrations. In addition, the capillaries in the brain are covered by the cell membranes of glial celIs. Thus, the tightly constructed capillary wall and the laidalditional g cell membranes result in a cellular barrier trhevaetnpts many drugs, proteins, ions, and other mole- ceouamdleislyfr r passing from the blood into the brain. In this way, the blood-brain barrier protects the brain from omisaonnysp circulating in the bloodstream. For example, the heartworm preventive drug ivermectin is poisonous to
CLINICAL APPLICATION
Epidural Anesthesia
We sometimes inject anesthetic agents into the space outside the spinal cord dura mater—the outermost layer of the meninges—to prod uce large areas of local anesthesia. Anesthetic drugs introduced in this way block depolarization waves through spinal nerves as they emerge from the dura mater and tehmuosvre the perception of pain from the part of the body they supply. This is called epidural anesthesia, because the anesthetic is injected into the space between the dura mater and uhe surrounding bone. Epidurals have the advantage of decreasing the perception of pain without having to anesthetize the brain. By not anesthetizing the brainstem and diencephalon, the body can more readily maintain its normal autonomic function during this type of anesthesia.
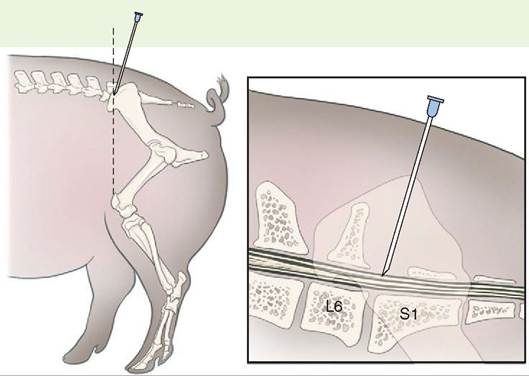
Needle placement for epidural anesthesia in the pig. L6 is the sixth lumbar vertebra, and S1 is the first sacral vertebra. (Redrawn from William WM, III, Hubbell J: Handbook of veterinary anesthesia, ed 5, St Louis, 2013, Mosby.)
∕ j CLINICAL APPLICATION
Myelogram
We can use the spaces between the meninges in veterinary medicine ⅛n an animal is suspected of having spinal trauma ⅛e to intervertebral disc disease. For example, Dachshunds often suffer from rupture of intervertebral discs, so-called slipped discs, between the cranial lumbar vertebrae and/or caudal thoracic vertebrae. The rupture of an intervertebral disc forces the gelatinous material inside the disc through the fibrous ring of the disc dorsally, where it pnresses o the spinal cord. The pressure exerted by the gelat- ienrioaul s mat compresses, or closes off, the space between the meninges on the ventral side (the underside) of the soprdinal c at the point of disc rupture. To identify the exis- tfence o this material pushing up against the spinal cord, we can inject a radiopaque dye—a dye that shows up white on x-rays—into the subarachnoid space in the spinal cord, which is the space just beneath the arachnoid membrane. uTbhseesquent radiograph will show places along the spinal
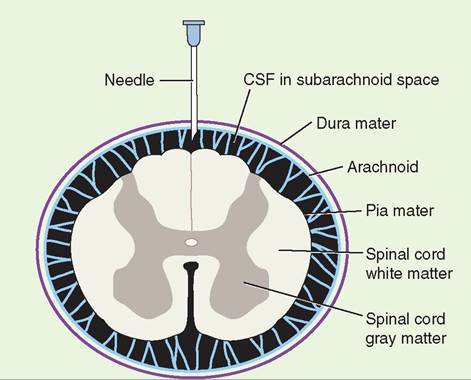
Transverse section showing the relationship among the meninges, the cerebrospinal fluid (CSF), and the spinal cord. The tip of the needle is in the subarachnoid space, as it would be for myelography. (Redrawn from Nelson R, Couto C: Small animal internal medicine, ed 4, St Louis, 2009, Mosby.)
cord where the dye did not flow. These areas are where the gelatinous disc material is pressing against the spinal cord and causing damage. This procedure is a form of contrast radiography called myelography.
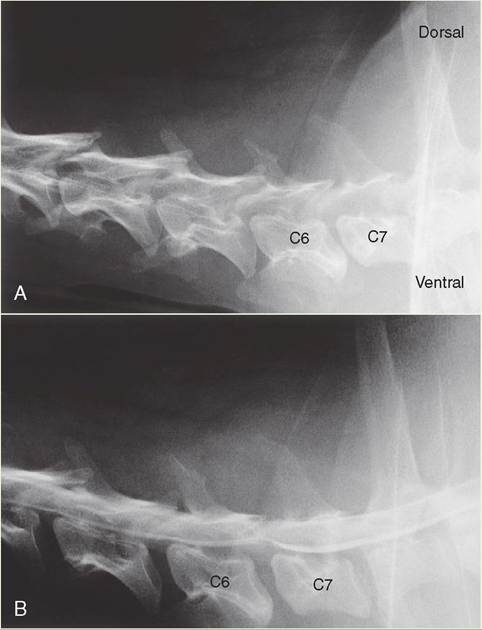
A, Plain radiograph. Cervical region of a 6-year-old Doberman Pinscher with a sudden onset of stumbling and weakness in the rear limbs, and mild cervical pain. B, Myelogram. The addition of the white-appearing myelography contrast media shows the normal smooth line of the spinal cord being compressed from the ventral side at the junction of vertebrae C6 and C7. Surgery revealed a large amount of ruptured disc material ventral to the spinal cord at this site. (From Nelson R, Couto C: Small animal internal medicine, ed 4, St Louis, 2009, Mosby.)
insects and parasites but, at the proper dose, does not adversely affect the dogs and cats that receive it. The reason for this selective toxicity of iverπmctin is that mammals have a blood-bnain barrier that prevents ivermectin from reaching tar get receptors on cells within the brain; however, insects and parasites do not have such a barrier, so the ivermectin can re adi Iy reach target receptors throughout the nervous system.
The blood-brain barrier can also prevent drugs that we administer from penetrating into the brain. If we want to treat an infection in the brain, for example, we must choose an antibiotic that is capable of crossing the blood-brain barrier and reaching the site of the infection.
CRANIAL NERVES
Cranial nerves are a special set of 12 nerve pairs in the peripheral nervous system that originate directly from the brain. Each pair of cranial nerves is conventionally numbered in Roman numerals from I through XII (1 through 12). The eranial nerve itself may contain axons of motor neurons, axons of sensory neurons, or combinations of both. Cranial nerve I (CN I, olfactory nerve) and cranial nerve II (CN II, optic nerve),ire both examples of pure sensory cvaaial nerves. The eorlfvaectory n is responsible for conveying sensory impulses
fercoempt orrs in the nose to the brain for the sense of smell,
ahpnteidc t o nerve is responsible for perception of light and vision. Unlike CN I and CN II, CN III is the oculomotor nerve and, as the name implies, it is a motor cranial nerve that controls eye movement. Other cranial nerves include CN V (trigeminal nerve), which controls muscles of the jaw for chewing and also conveys sensations from the nose, mouth, and part of the throat. Thus, CN V is an example of a cranial nerve that serves both sensory and motor functions. See Table 9-1 for a list of the different functions of the 12 cranial nerves.
Table 9-2 shows two mnemonic devices (memory aids) that can be used for remembering the names of the cranial
TEST YOURSELF 9-6
1. What are the protective membranes that surround, support, and protect the CNS?
2. What is the fluid called that bathes, cushions, and aids in transport of materials to and from the CNS?
3. What helps keep dangerous poisons and certain drugs from leaving the blood and entering the brain? Describe this structure.
4. What are the 12 cranial nerves? Which nerves are motor, which are sensory, and which are both? nerves and their functions (sensory, motor, or both). In the first mnemonic, each word of the saying begins with the same letter as the corresponding cranial nerve. Of course, you have to remember which O represents which cranial nerve in the first three cranial nerves, but the saying still helps students remember the names of the cranial nerves more readily.
The second mnemonic tells whether the cranial nerve in question is a sensory nerve, a motor nerve, or both sensory and motor. In the saying, the words beginning with S indicate that the corresponding nerve is primarily sensory. If the word begins with M, the corresponding nerve is primarily a motor nerve; and if the word begins with B, the nerve is both sensory and motor.
SPINAL CORD
The spinal cord is the caudal continuation of the brainstem outside the skull that continues down the bony spinal canal formed by the vertebrae. It conducts sensory information and motor instructions between the brain and the periphery of the body, but it is not simply a “cable” of nerve fibers. It
| TABLE 9-1 | Functions of the 12 Cranial Nerves | |||||
| NUMBER | NAME | TYPE | KEY FUNCTIONS | |||
| I | Olfactory | Sensory | Smell | |||
| II | Optic | Sensory | Vision | |||
| III | Oculomotor | Motor | Eye movement, pupil size, focusing lens | |||
| IV | Trochlear | Motor | Eye movement | |||
| V | Trigeminal | Both sensory and motor | Sensations from the head and teeth, chewing | |||
| VI | Abducent | Motor | Eye movement | |||
| VII | Facial | Both sensory and motor | Face and scalp movement, salivation, tears, taste | |||
| VIII | Vestibulocochlear | Sensory | Balance, hearing | |||
| IX | Glossopharyngeal | Both sensory and motor | Tongue movement, swallowing, salivation, taste | |||
| X | Vagus (wanderer) | Both sensory and motor | Sensory from gastrointestinal tract and respiratory tree; motor to the larynx, pharynx, parasympathetic; motor to the abdominal and thoracic organs | |||
| XI | Accessory | Motor | Head movement, accessory motor with vagus | |||
| XII | Hypoglossal | Motor | Tongue movement | |||
| TABLE 9-2 | Mnemonic Devices for Cranial Nerve Names and Functions | |||||
| CRANIAL NERV | 'E NERVE NAME | WORD OF THE SAYING | TYPE OF NERVE | WORD OF THE SAYING | ||
| I | Olfactory | On | Sensory | Six | ||
| II | Optic | old | Sensory | sailors | ||
| III | Oculomotor | Olympus' | Motor | made | ||
| IV | Trochlear | towering | Motor | merry, | ||
| V | Trigeminal | top, | Both sensory and motor | but | ||
| VI | Abducent | a | Motor | my | ||
| VII | Facial | fine, | Both sensory and motor | brother | ||
| VIII | Vestibulocochlear | vocal | Sensory | said, | ||
| IX | Glossopharyngeal | German | Both sensory and motor | "Bad | ||
| X | Vagus | viewed | Both sensory and motor | business, | ||
| XI | Spinal accessory | some | Motor | my | ||
| XII | Hypoglossal | hops | Motor | man” | ||
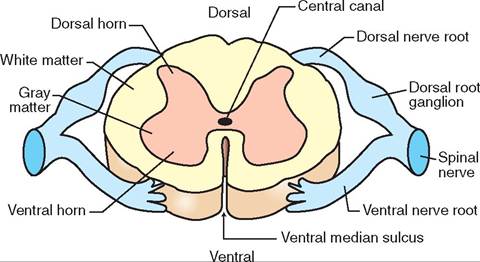
FIGURE 9-1 1 Cross section of the spinal cord.
contains many neuron cell bodies (in the gray matter) and extensive synapses (connections) between ascending nerve fibers conducting sensory information toward the brain and descending nerve fibers conducting motor information to muscles and other organs. A considerable amount of processing and modification of nerve impulses between the brain and the peripheral nerves takes place in the spinal cord. This is important to remember when we are trying to prevent or treat severe pain in our patients. (See the Clinical Application on Preventing Windup in Chapter 10.)
The positions of the gray matter and white matter of the spinal cord are reversed from the arrangement in the cerebrum and cerebellum of the brain. In cross section (Figure 9-11), the gray matter of the spinal cord is located in the medulla (inner part) and takes the shape of a butterfly with the CSF-containing central canal running its length in the center. The white matter of the spinal cord forms the cortex (outer part) that surrounds the gray matter. As in the brain, the gray matter contains many neuron cell bodies, and the white matter consists mainly of myelinated nerve fibers.
Between each pair of adjacent vertebrae, the spinal cord sends off dorsal and ventral nerve roots from each side that combine to form left and right spinal nerves; these link the spinal cord with peripheral nerves. The dorsal nerve roots contain sensory (afferent) fibers, and the ventral nerve roots contain motor (efferent) fibers. So, sensory information comes into the spinal cord via the dorsal nerve roots, and motor instructions go out to the body via the ventral nerve roots. The neurons that process and carry sensory (afferent) nerve impulses to the brain or other parts of the spinal cord are located in what are called the dorsal horns of the spinal cord's gray matter “butterfly” (see Figure 9-11). The neurons that process and carry motor (efferent) nerve impulses to the spinal nerves are located in the ventral horns of the gray matter.
THE AUTONOMIC NERVOUS SYSTEM
As mentioned previously, the autonomic nervous system controls many functions of the body at a subconscious level. These automatic functions are performed by two divisions of the autonomic nervous system: the sympathetic nervous system and the parasympathetic nervous system. These two systems generally have opposite effects on organs or tissues, and whichever system dominates at any given moment determines how excited or relaxed things are in the body.
STRUCTURE
The first anatomic difference between these two systems is where the peripheral nerves of each system emerge from the CNS. The nerves for the sympathetic nervous system emerge from the thoracic and lumbar vertebral regions in the back. Thus, the sympathetic system is often referred to as the thoracolumbar system. In contrast, the parasympathetic system emerges from the brain and the sacral vertebral regions and therefore is called the cranial-sacral system.
The efferent motor nerves of both the sympathetic and parasympathetic nervous systems are composed of a sequence of two neurons. The first neuron has its cell body in the brain or spinal cord and extends its axon out from the CNS to a cluster of neuronal cell bodies outside of the CNS called an autonomic ganglion. Here the first neuron synapses with one or more second neurons, which then connect to the target organ (e.g., endocrine gland, smooth muscle). The first neuron is called the preganglionic neuron, because it is before the ganglion. The second neuron is the postganglionic neuron, because it carries the impulse from the ganglia to the target organ.
The sympathetic and parasympathetic systems also differ anatomically in the length of the preganglionic and postganglionic neurons (Figure 9-12). The sympathetic preganglionic neuron originates in the thoracic and lumbar segments of the spinal column. Outside the thoracolumbar area of the spinal column are a series of autonomic ganglia (many ganglions) that form a chain called the sympathetic ganglion chain. The sympathetic preganglionic neuron extends out from the spinal cord and either synapses with a neuron within the ganglion chain or passes through the ganglionic chain and synapses with a neuron located beyond the sympathetic chain. Each sympathetic preganglionic neuron usually synapses with many postganglionic neurons in a wide variety of locations in the sympathetic chain or in ganglia outside the sympathetic chain. This helps explain why sympathetic nervous system responses are usually spread throughout the body and involve several organs simultaneously.
The sympathetic postganglionic neuron extends the remaining distance to the target organ. Therefore, the sympathetic postganglionic neuron is much longer than its corresponding preganglionic neuron.
Unlike the short sympathetic preganglionic neuron, the parasympathetic preganglionic neuron is quite long and originates from the nuclei (clusters of neurons in the brain) of several cranial nerves and from the sacral region of the spinal cord. Thus the parasympathetic system is called the cranial-sacral division of the autonomic nervous system.
In contrast to the sympathetic preganglionic neuron, which terminates close to the spinal cord, the parasympathetic preganglionic neuron travels directly from the
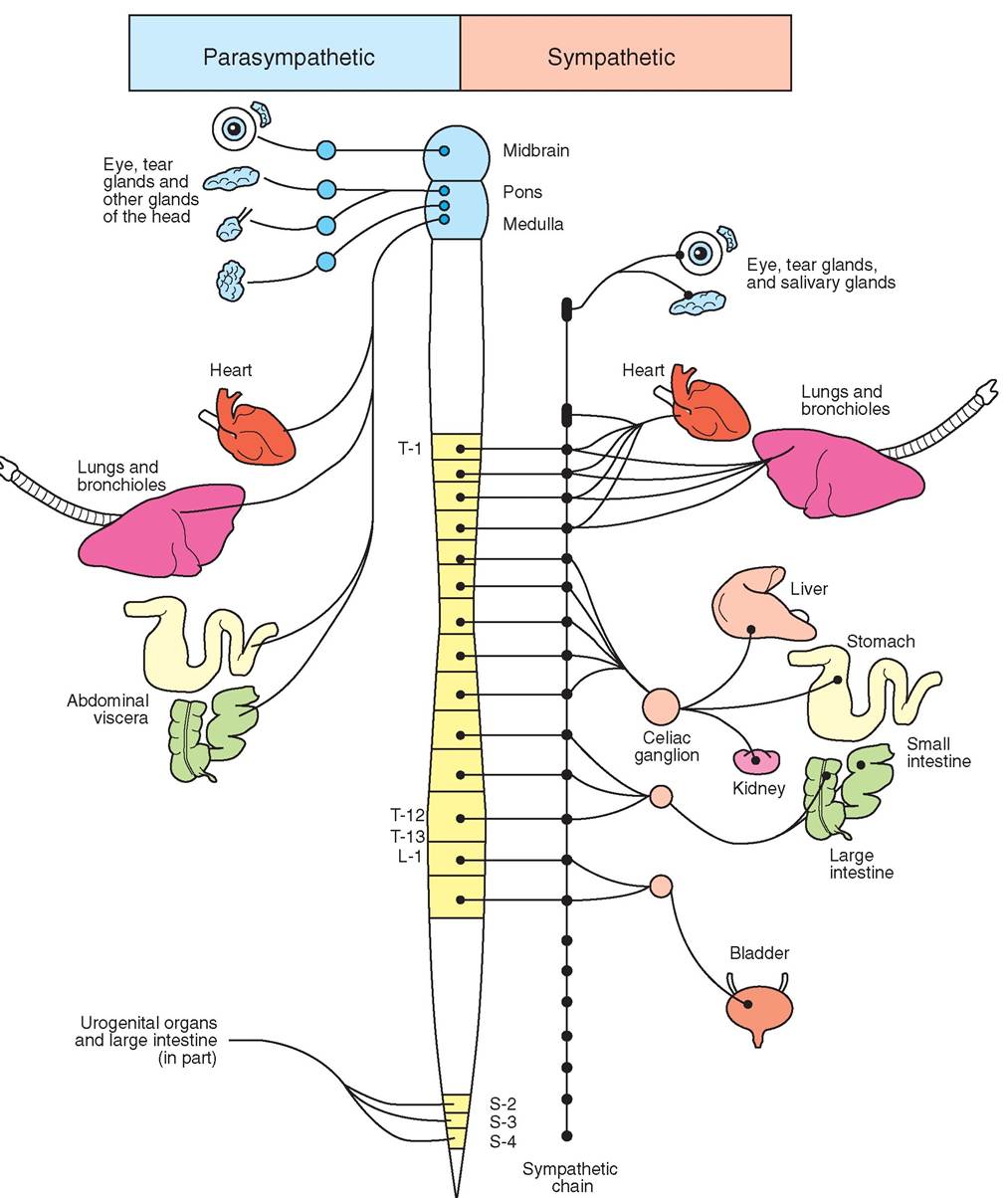
FIGURE 9-1 2 Structure of parasympathetic and sympathetic nervous systems.
CNS to its target organ, where it synapses with a short postganglionic neuron in the target organ. Thus the parasympathetic preganglionic neuron is relatively long compared with the short postganglionic neuron.
GENERAL FUNCTIONS
The sympathetic nervous system is often called the fight-or- flight system, meaning that this is the system that helps the body cope with emergency situations in which an animal might have to defend itself (fight) or escape (flight). In contrast, the parasympathetic nervous system could be called the rest-and-restore system because of its ability to decrease the strong excitatory effects of the fight-or-flight system (bring the body back to resting state) and its ability to facilitate all the processes that will replace those body resources used up during the emergency (restore).
Using this simple model of fight or flight or rest and restore, we can make sense of the clinical signs observed with stimulation of either part of the autonomic nervous system. Table 9-3 summarizes these effects. In a fight-or-flight situation, the animal needs to move rapidly, therefore the muscles are going to be working vigorously. To meet the needs of the muscles, the bronchioles (airway passages) increase in diameter (bronchodilation) to allow a greater exchange of oxygen and carbon dioxide. Once the oxygen leaves the lungs and enters the blood, it must be delivered rapidly. Therefore heart rate and the force of cardiac contractions increase, which increases the rate at which blood is moved around the body. Finally, to deliver more blood into the working muscles, the small blood vessels (arterioles) supplying the muscles dilate (vasodilation).
In contrast to the dilation of skeletal muscle blood vessels under sympathetic stimulation, the small blood vessels supplying the skin, gastrointestinal tract (GI tract), and the kidney constrict, thereby reducing blood flow to these areas
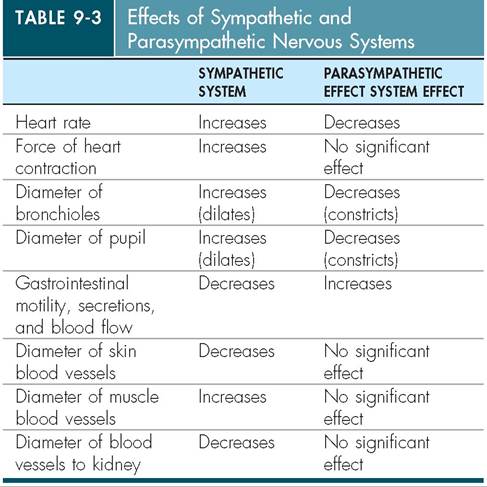
so that blood can be redirected to the muscles. During this time, when blood is redirected away from these organs, the digestive and absorptive functions of the GI tract and the filtering function of the kidney are temporarily suspended or decreased until the crisis passes. The decreased blood supply to the skin also means that superficial wounds will bleed less.
Why would eye pupil dilation be of benefit to an animal in a fight-or-flight situation? Opening up the pupil admits more light, but an additional benefit may be an increase in peripheral vision (you can see more “out of the corner of the eye”). Thus the sympathetic nervous system may allow the animal to take in a wider field of vision and see better under low-light conditions.
It’s pretty easy to see how the parasympathetic nervous system antagonizes (works against) many but not all of the sympathetic nervous system effects. The parasympathetic system causes the GI tract to increase its activity, thus digesting and absorbing nutrients that are needed to replenish the energy resources used during the fight-or-flight situation. The parasympathetic system also reduces the heart rate and reduces the sympathetic system’s dilation of the bronchioles. Except for the GI tract, the parasympathetic system has little effect on the blood vessels in most parts of the body, so the return to normal vessel diameter is handled by other regulatory mechanism.
CLINICAL APPLICATION
The Sympathetic Nervous System Response to Shock or Blood Loss
One of the key roles of the sympathetic nervous system is to maintain arterial blood pressure. The arterial blood pressure can drop to a point where the brain may not be receiving adequate blood flow in situations involving a loss of blood volume; a loss of fluid in the blood, as with dehydration; or a large-scale dilation of blood vessels throughout the body, as in the case of shock. The body responds to this loss of arterial blood pressure by causing massive stimulation of the sympathetic nervous system. The heart pounds rapidly and fiercely in the chest to increase the output of blood into the arteries; simultaneously, the small blood vessels, or arterioles, in the skin, GI tract, kidney, and other areas constrict. The increased cardiac output and arteriole vasoconstriction result in an increased arterial blood pressure and more blood directed to the brain.
The increase in blood arterial pressure from the increased cardiac output and vasoconstriction would be like the increase in air pressure within a rubber tube if you blew forcefully into the tube while simultaneously pinching the far end. The narrowing of the small blood vessels supplying the skin also explains why the skin and mucous membranes of people or animals with a sudden decrease in blood pressure appear so pale.
Drugs or diseases that imitate, stimulate, or inhibit either the parasympathetic or sympathetic nervous system produce physiologic changes that mimic one of these branches of the autonomic system. Therefore the veterinary professional should know how and why the sympathetic and parasympathetic nervous systems affect the body.
NEUROTRANSMITTERS AND RECEPTORS
The sympathetic nervous system primarily uses norepinephrine as its neurotransmitter. As stated previously, norepinephrine, epinephrine, and dopamine are all part of a group of neurotransmitters and hormones called catecholamines. Because epinephrine and norepinephrine used to be called adrenaline and noradrenaline, respectively, the neurons that release norepinephrine are said to be adrenergic neurons. Note: The terms adrenaline and noradrenaline are still used in some countries. In addition to the release of norepinephrine from postganglionic adrenergic neurons, the body's sympathetic response also comes from the release of epinephrine and norepinephrine from the medulla (inner part) of the adrenal gland. The adrenal medulla acts like a cluster of adrenergic neurons releasing the epinephrine and norepinephrine directly into the bloodstream, where they are quickly distributed to receptors throughout the body.
Neurotransmitters can only work on cells that contain specific receptors capable of binding to the particular neurotransmitter molecules. So, blood vessels in the skin, the GI tract, and the skeletal muscle must have adrenergic receptors for epinephrine or norepinephrine. What we find is that there are actually different types of adrenergic receptors in the tissues or organs affected by the sympathetic nervous system. Alpha1-adrenergic receptors typically are found on blood vessels and cause the vasoconstriction of the skin, GI tract, and kidney associated with sympathetic stimulation. The increases in heart rate and force of contraction are the result of stimulation of beta1-adrenergic receptors, and the bronchodilation associated with sympathetic stimulation results from beta2-adrenergic receptor stimulation.
The effects and side effects of drugs that mimic the sympathetic nervous system can be explained by the specificity (selectiveness) of the drug molecule for alpha1, beta1, or beta2 receptors. Therefore it is important for the veterinary professional to remember the different adrenergic receptors and the organs or tissues with which they are associated.
The neurons associated with the parasympathetic nervous system secrete acetylcholine as their neurotransmitter. Therefore these neurons often are referred to as cholinergic neurons. Even though norepinephrine is the neurotransmitter associated with sympathetic nervous system effects, the preganglionic neuron (the neuron that emerges from the CNS and synapses with the postganglionic neuron) in both the sympathetic and parasympathetic nervous system is a cholinergic neuron that releases acetylcholine. That may seem a little strange, but remember the ultimate effect of the sympathetic and parasympathetic systems is determined by their postganglionic neurons.
Like the adrenergic receptors of the sympathetic nervous system, the cholinergic receptors for acetylcholine also come in different types. The two types of acetylcholine receptor are called nicotinic and muscarinic. The nicotinic
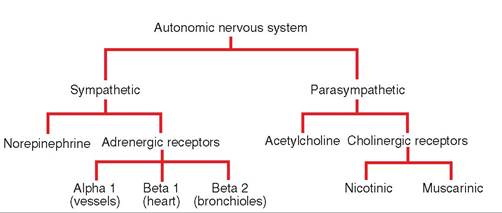
FIGURE 9-13 Simplified concept map for the autonomic nervous system.
receptors are found primarily on the postganglionic neurons of both the sympathetic and parasympathetic nervous system, as well as between motor neurons and muscle in the somatic (voluntary) motor system (Figure 9-13). The muscarinic receptors are found on the target organs and tissues supplied by the postganglionic neurons of the parasympathetic nervous system. Certain toxins (e.g., nicotine or plants containing nicotine-like compounds) may selectively stimulate the nicotinic receptors more than the muscarinic receptors, whereas others may selectively affect the muscarinic receptors. The clinical signs of the toxicities reflect which receptors have been stimulated.
TEST YOURSELF 9-7
1.
2.
3.
4.
Which part of the autonomic nervous system is responsible for the fight-or-flight response and which is responsible for the rest-and-restore system?
Compare and contrast the sympathetic and parasympathetic nervous systems. Include in your comparison the preganglionic and postganglionic neurons; the origin of preganglionic neurons; the neurotransmitters; and each system's impact on the heart, GI tract, blood vessels, bronchiole diameters, and the size of the pupil of the eye.
With which branch of the autonomic nervous system are the alpha1, beta1, and beta2 receptors associated? What happens to the body when these particular receptors are stimulated?
With which branch of the autonomic nervous system are muscarinic and nicotinic receptors associated?
REFLEXES AND THE REFLEX ARC
Reflexes are rapid, automatic responses to stimuli designed to protect the body and maintain homeostasis. Reflexes can be somatic reflexes, which involve contraction of skeletal muscles, or autonomic reflexes, which regulate smooth muscle, cardiac muscle, and endocrine glands.
Regardless of whether the reflex is somatic or autonomic, all reflexes have the same basic structure, called the reflex arc. The reflex arc originates from a sensory receptor, which detects a change either in the external environment or within the body itself. Once stimulated to threshold, the sensory receptor sends an action potential (nerve impulse) along the sensory neuron to the gray matter of the spinal cord or brainstem. In the CNS gray matter, the sensory neuron synapses with other interneurons, which serve to integrate the incoming sensory impulse with other impulses from other sensory neurons. Finally, the integrated response of the reflex is sent out from the spinal cord or brainstem by the motor neuron, which ends at the target organ (muscle or endocrine gland). If the motor sonmeuartoicn i a s neuron, the reflex arc ends in contraction or inhibition of skeletal muscle. If it is an autonomic neuron, the reflex arc ends in smooth muscle within an organ or blood vessel, cardiac muscle, or endocrine gland.
Stimulation of skeletal muscle (somatic) reflex arcs is commonly used in veterinary medicine to aid in the diagnosis of spinal cord trauma, peripheral nerve damage, or muscle disease. Various types of somatic reflex are evaluated by the veterinarian, including the stretch reflex, the withdrawal reflex, and the extensor reflex. Understanding how these somatic reflex arcs function and are regulated is important for comprehending how normal and abnormal reflexes can indicate disease or damage in the nervous system or musculoskeletal system.
STRETCH REFLEX
The stretch reflex is considered a simple, monosynaptic or two-neuron reflex arc, because it involves only a sensory neuron and a motor neuron (with only one synapse between them) without any interneurons (Figure 9-14). The sensory receptor in the stretch reflex arc is a specialized structure within the muscle called the muscle spindle. If a muscle is stretched, the muscle spindle also stretches and sends impulses via the somatic sensory neuron to the spinal cord. At the spinal cord, the sensory neuron synapses with the motor neuron that innervates the same muscle. Stimulation of the motor neuron causes that muscle to contract in response to the stretching of the muscle. In this way, the body can maintain the tension or tone of a muscle to meet an increased force applied to stretch the muscle, or it can prevent an overstretching of a muscle caused by contraction of opposing muscles.
You may have had this reflex tested in your own doctor's office, when your patellar ligament (located just below your kneecap) was tapped. In this situation, the force against the ligament slightly stretched the large quadriceps femoris muscle on your thigh, stimulating a reflex arc that resulted in contraction of the quadriceps and the subsequent extension—a slight kick—of the lower leg.
When stimulated by the stretch receptor, the afferent somatic sensory neuron from the muscle spindle does more than just cause the stretched muscle to contract. Branches off the sensory neuron will synapse with another reflex arc to cause the opposing muscles to relax. Therefore, when the stretch receptor in your quadriceps femoris muscle initiated the stretch reflex arc, a branch of the sensory neuron also synapsed with an inhibitory interneuron in the spinal cord. This inhibitory interneuron released inhibitory neurotransmitters at a synapse with the motor neuron to the semimembranosus and semitendinosus (hamstring) muscles, the muscles that normally oppose the quadriceps femoris muscle. By inhibiting stimulation of these opposing muscles, the quadriceps femoris muscle could contract without antagonism, or without being worked against by the opposing muscle.
Finally, another branch off the stretch receptor's sensory neuron enters the spinal cord and goes up to the brain so that the cerebellum can coordinate the movement and the conscious sensory centers in the cerebral cortex can be informed of the stimulation and subsequent limb movement. Note: The patellar tap reflex is a pure reflex that cannot be overridden by conscious effort. Even if you tighten up your leg muscles and do your best not to kick when the patellar ligament is tapped, you cannot completely prevent that kick from occurring.
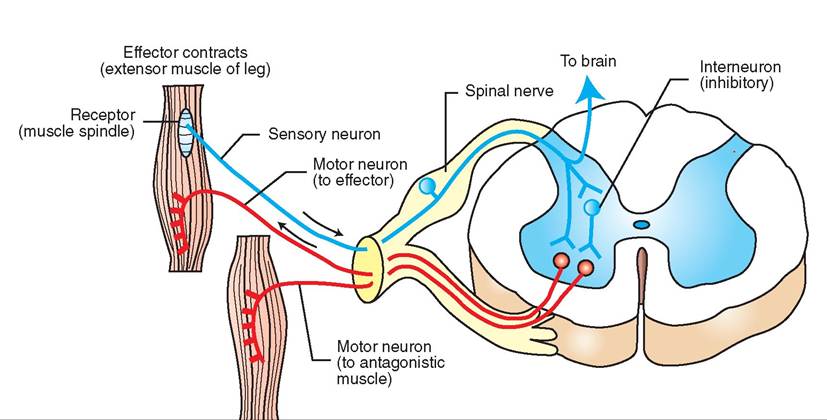
Antagonistic
muscle relaxes
(flexor muscle of leg)
FIGURE 9-14 Basic reflex arc as illustrated by simple stretch reflex.
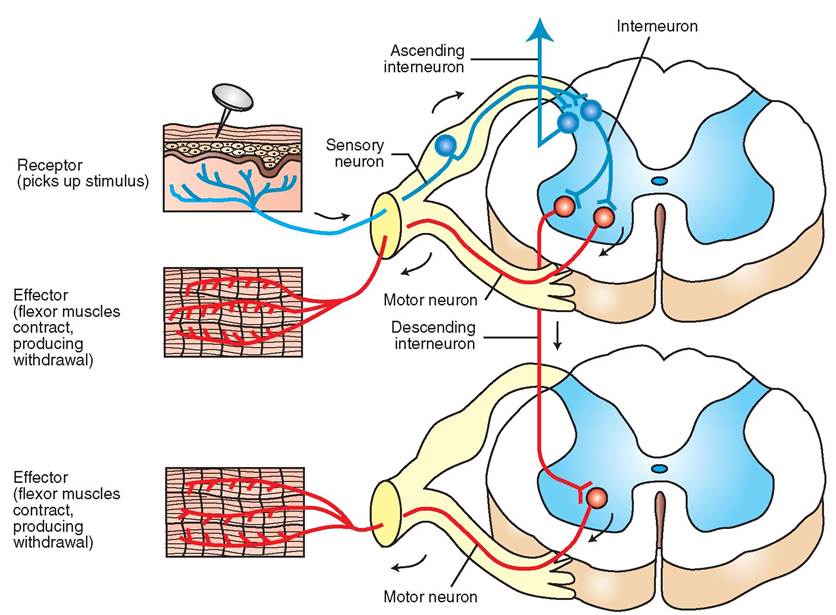
FIGURE 9-15 Flexor (withdrawal) reflex. The reflex arc involves several spinal segments. Some branches of the spinal reflex arc extend to motor nerves in other spinal segments, resulting in relatively complex reflex movement.
WITHDRAWAL REFLEX
The withdrawal reflex, also called the flexor reflex, happens when you rapidly withdraw a limb or flex the joints after accidentally touching a hot stove or stepping on a sharp object with your bare foot (Figure 9-15). In both of these situations, a strong stimulus to a receptor causes the sensory somatic neuron to send impulses to the spinal cord. Unlike the simpler stretch reflex, the withdrawal reflex involves synapses with several interneurons. Some of these interneurons will synapse with motor neurons that will cause contraction of a specific set of muscles responsible for pulling the limb away from the painful stimulus. Other interneurons will inhibit those opposing muscle groups so that the withdrawal of the limb is rapid and complete. This reflex can be complicated, because many different muscles may be involved in the withdrawal from the painful stimulus.
Even though the reflex arc may involve several interneurons, several motor neurons, and several different segments of the spinal cord, the reflex occurs without the brain being aware of the incident. In other words, the limb has been withdrawn from the painful stimulus before the brain becomes consciously aware of the painful stimulus.
CROSSED extensor reflex
If you step on a sharp tack, which results in the leg being withdrawn rapidly before the brain is aware of what is going on, you should fall over. The reason you don't is that when the withdrawal reflex arc is stimulated, the afferent somatic sensory neuron also synapses with another set of interneurons that cause extensor muscles in the opposite leg to contract and support the full weight of your body when the other leg flexes. This reflex is called the crossed extensor reflex, because the afferent sensory impulse crosses to the other side of the spinal cord and stimulates the muscles that extend the opposite limb (Figure 9-16).
Reflexes that start on one side and travel to the opposite side of the body are said to be contralateral reflexes; the crossed extensor reflex is one example. Reflexes like the stretch reflex are called ipsilateral reflexes, because the stimulus and response are on the same side of the body. The veterinary technician needs to be familiar with the terms Contralateral and ipsilateral because they are used for a variety of applications in veterinary medicine and do not pertain only to reflexes.
THE ROLE OF THE UPPER CNS IN MODERATING REFLEXES
An animal with severe spinal cord damage in the area of the first or second lumbar vertebrae (L1 or L2) can still have reflexes in the hind limbs. Even though sensory impulses from the hind limbs are blocked from reaching the brain, and conscious motor impulses from the brain to the hind limbs are blocked by the damaged section of the spinal cord, the reflex arcs in the lumbar spinal cord segments caudal to
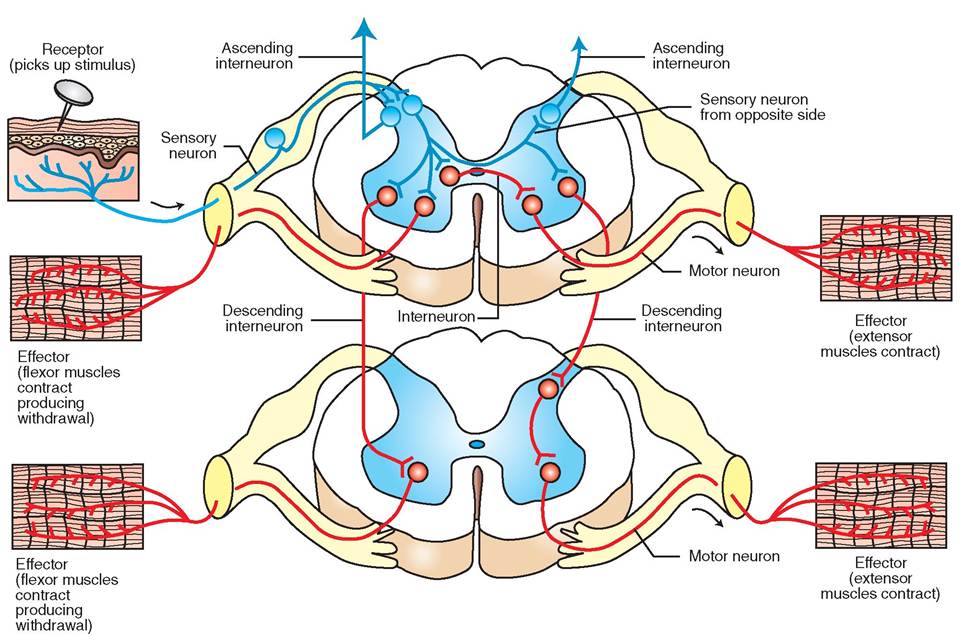
FIGURE 9-16 Crossed extensor reflex.
the trauma are still intact and can still function. The animal, however, is unaware that the hind limbs are moving, because it cannot receive any sensory information as a result of the spinal cord damage at L1 to L2.
Not only are the reflex arcs functioning in the hind limbs despite the damage to the spinal cord at L1 to L2, but the reflex: responses are hyperreflexive. This means when the veterinarian taps on the patellar ligament, the quadriceps omnutsrcalcets c with more force and produce more limb movement than normal. This illustrates an important cdoncept use by veterinarians to diagnose location of spinal cord injuries. The upper CNS normally produces a dampening yr inhibitory effect on the reflex arcs. Thus, under normal conditions, the patellar ligament stretching usually only produces a relatively small kick because of the dampening effect of the upper CNS. With spinal cord trauma, this dampening eofmfect fr the upper CNS is blocked, and any intact reflex arcs caudal to the spinal cord trauma are now exaggerated, or hyperreflexive.
If spinal cord trauma occurs where the reflex arc enters voerslea the spinal cord, or if the sensory nerve or motor fnerve o the reflex arc is damaged, the reflex will be less than normal (hyporeflexive) or absent altogether. In other words, icf the ar is damaged or broken anywhere, the reflex will either not function, or it will function weakly.
By stimulating different somatic reflex arcs, knowing thhicrohugh w spinal cord segments the reflex arc travels, and observing whether the response is normal, hyperreflexive, or hyporeflexive, the veterinarian can localize ftehae ar o the spinal cord in which trauma or disease has occurred.
OTHER CLINICALLY SIGNIFICANT REFLEXES
Many reflexes are used to assess the clinical condition of patients in veterinary medicine. Most are complex reflexes relating to posture, gait, the animal's ability to right itself, caenmd etnhte pla of limbs. Two other reflexes that the veterinary technician should understand are the palpebral reflex and the pupillary light reflex (PLR). These two reeflexes ar routinely used when assessing an animal for fdepth o anesthesia and when performing a physical examination.
eTbhrealpalp reflex arc originates from receptors on the eyelid margins, travels via sensory neurons in CN V to the pons (brainstem), synapses with neurons in the pons, then itaravels v CN VII to the muscles that blink the eyelids. If the rcetiflvex, is a a light tap on the medial canthus of the eye
(dtihael me corner of the eye, where the top and bottom etyaelids me produces a blink of the eyelids. When an animal is anesthetized, the neurons in the pons become less responsive. Therefore, as anesthesia deepens, the palpebral reflex (also called the palpebral blink reflex) bmomes less responsive and provides an indication of the animal's depth of anesthesia.
The PLR is a test for a reflex arc that includes the retina (light-sensing layer of the eye), the optic nerve (CN II),
neuron cl listers in the diencephalon of the brain, and motor neurons of CN III (which supply the muscle of the iris that constricts the size of the pupil). A normal response to shining light in the eye of an animal is for the iris in both eyes to constrict, thus making the pupil smaller. Normally, this opnusptirllicatriyonc would reduce the amount of light
entering the eye and protect the retina against bright light. Because shining the light in one eye causes a constriction in
yoth eyes, the reflex arc must cross over to the other side of the body. By examining the PLRs in both eyes, a veterinarian can eveluate each reflex arc and identify whether an animal ihsausala v problem in the retina, the optic nerve, or the motor nerve. Note that the PLR does not assess vnision. A animal with damage to the visual centers in the coerrtexbral c can be blind and still have normal PLRs in
ybeost.h e
CLINICAL APPLICATION
Dachshund With Intervertebral Disc Disease
Gretchen was an 8-year-old, spayed, female Miniature Dachshund brought in when it became apparent that she was suddenly unable to move her hind legs. On examination, the veterinarian observed that Gretchen could not stand on her rear legs and did not yelp when the skin on her rear toes was pinched. She did, however, react to firm squeezing of the toes themselves. The veterinarian concluded that spinal cord trauma was interfering with the sensory impulses reaching the brain and with conscious motor impulses from the brain reaching the muscles of the hind limb.
Given the history and Gretchen’s breed, spinal trauma from tuhpet urre of an intervertebral disc seemed the most likely cause. hr these situations, the material from the disc between vertebrae can rupture forcibly against the adjacent spinal cord, producing severe trauma and swelling that renders the spinal cord at that segment nonfunctional.
Before ordering radiographs, the veterinarian must localize ftehae ar o the spinal cord where the trauma is most likely to have occurred. In this way, the radiograph can be focused directly ∞^ the suspected site with the vertebrae lined up to aeltltoerw a b view of disc material in the spinal canal and any fnarrowing o the affected disc space.
Gretchen’s veterinarian tested some somatic reflexes— withdrawal and crossed extensor among them—in the hind limb and determined that the reflexes that travel through segments of spinal cord caudal to L3 were all hyperreflexive. This indicated that the reflex arcs were functional but were not being dampened by the upper CNS. Therefore the veterinarian could conclude that these spinal cord segments were caudal to tohred spinal c trauma. The reflex arcs that traveled through L1, L2, and L3 were all hyporeflexive or nonexistent. The reflex arcs at these spinal cord segments were broken because of the torauma t the spinal cord segments. The reflexes that involved soprdinal c segments cranial to T13 were normal, so these soprdinal c segments were unaffected and still had communi- ictahtion w the upper CNS.
eTthereinvarian ordered the radiographs centered over rLne2t choen. G The radiographs indicated narrowed
ecdetiwsceesnpa b vertebrae L1 and L2. Because Gretchen’s ouwshneedr r her in quickly, the veterinarian could amrreargnegnecye surgery to relieve the pressure on the
soprdinal c by removing part of the vertebral bone over the eafafse.cted ar Gretchen slowly recovered over several days, until she con Id walk with 90% of her normal strength and control.
TEST YOURSELF 9-8
1. Whot αrethe differences betweenan Outonomicreflexand asomatic reflex?
2.Whet rol255
Pain, 256
Proprioception, 256
SPECIAL SENSES, 257
Taste, 257
Smell, 258
Hearing, 258
Equilibrium, 264
Vision, 266
LEARNING OBJECTIVES
When you have completed this chapter you will be able to:
1. List the four general types of stimulus that can trigger a response from sensory receptors.
2. List and describe the visceral senses.
3. Differentiate between superficial and central temperature sensors.
4. List and describe the processes that contribute to nociception.
5. Describe the structure of the taste buds.
6. List and describe the special senses.
7. Describe the structure and function of the components of the ear.
8. Describe the processes that contribute to the sense of equilibrium.
9. Describe the structure and function of the components of the eyeball.
10. Describe the structure of the conjunctiva and eyelids.
11. Describe the origin of tears and explain how tears flow onto and drain from the eye.
VOCABULARY FUNDAMENTALS
Accommodation uh-kohm-uh-da-shuhn
Ampulla ahm-pyoo-luh
Analgesia ahn-ahl-je-zhuh
Anesthesia ahn-uhs-the-zhuh
Anterior chamber ahn-teer-e-ar cham-bar
Aqueous compartment a-kwe-uhs kuhm-pahrt-mehnt
Aqueous humor a-kwe-uhs hyoo-mar
Bulbar conjunctiva buhl-bar kohn-juhnck-tih-vah
Canal of Schlemm kuh-nahl of shlehm
Canthus kahn-thuhs
Choroid kohr-oyd
Ciliary body sihl-e-ahr-e boh-de
Ciliary muscle sihl-e-ahr-e muhs-uhl
Cochlea kok-le-ah
Cochlear duct kok-le-ahr duhckt
Cone kon
Conjunctiva kohn-juhnck-tih-vah
Conjunctival sac kohn-juhnck-tih-vahl sahck
Cornea kohr-ne-uh
Crista krihs-tuh
Cupula kuh-pu-luh
Eardrum er-druhm
Endolymph ehn-do-lihmf
Equilibrium e-kwuh-lihb-re-uhm
Eustachian tube yoo-sta-shehn toob
External auditory canal ehcks-tar-nahl ahw-dih-tohr-e kuh-nahl
External ear ehcks-tar-nahl er
Extraocular eye muscle ehcks-trah-ah-kyoo-lahr ι muhs-uhl
Eyelid ι-lihd
Fundus fuhn-duhs
General anesthesia jehn-ar-ahl ahn-uhs-the-zhuh
General sense jehn-ar-ahl sehns
Gustatory sense guhs-tuh-tohr-e sehns
Hearing hihr-ihng
Heatstroke het-strok
Hyperthermia hι-par-thuhr-me-uh
Hypothermia hι-po-thuhr-me-uh
Incus ihn-kuhs
Inner ear ihn-ar er
Lacrimal apparatus lah-kreh-mahl ahp-uh-raht-uhs
Lacrimal^ιd lah-kreh-mahl glahnd
Lncrimal puncta lah-kreh-mahl puhngk-tuh
Limbus lihm-buhs
Local anesthesia lo-kuhl ahn-uhs-the-zhuh
Macula mahck-u-lah
Malleus mahl-e-uhs
Meibomian glanr mih-bo-me-ahn glahnd
Mirrle ear mihd-uhl er
Morulation mohd-u-la-shuhn
Nasolacrimal duct na-zo-lahck-rihm-ahl duhckt
Nictitating membrane nihck-tih-ta-tihng mehm-bran
Nociception no-sih-sehp-shuhn
Nociceptor no-sih-sehp-tuhr
Olfactory ^se ohl-fahck-tuh-re sehns
Optic disc ohp-tihck dihsk
Organ of Corti ohr-gahn of kor-te
Ossicle ohs-eh-kuhl
Otolith o-to-lihth
Oval window o-vahl wihn-do
Pain pan
Palpebral conjunctiva pahl-pe-brahl Mn-juhnck-tih-vah
Perception puhr-sehp-shuhn
Perilymph pear-ih-lihmf
Photoreceptor fo-to-reh-sehpt-ar
Pinna pihn-nuh
Post∣erior chamber po-steer-e-ar cham-bar
Pressure prehsh-ar
Proprioception pro-pre-o-sehp-shuhn
Pupil pu-puhl
Retina reh-tih-nuh
Ror rohd
Rounr window round wihn-do
Saccule sahck-ul
Sclera sklear-uh
Semicircular canal seh-me-suhr-kyoo-lar kuh-nahl
Sensory receptor sehn-sar-e reh-sehpt-ar
Smell smehl
eSnpseec ial s spehsh-hunhzl se
Stapes sta-pes
Suspensory ligament suh-spehn-suh-re lihg-ah-mehnt
Tactile ^se tahck-tihl sehns
Tapetum tah-pe-duhm
Tapetum lucirum tah-pe-duhm loo-sihd-uhm
lTaanrrsa l g tahrl-ashahnldg
Taste tast
Tectorial membrane tehck-tohr-e-uhl mehm-bran
Temperature sense tehm-par-uh-char sehns
Thirr eyelid thihrd ι-lihd
Touch tuhch
Transruction trahnz-duhck-shuhn
Transmission trahnz-mihsh-uhn
Tympanic membrane tihm-pahn-ihck
mehm-bran
Utricle u-trih-kuhl
Uvea u-ve-uh
Vestibule vehs-tuh-byool
Visceral sensation vih-sar-ahl s⅛ι-sa-shuhn
Vitreoum compartment vih-tre-uhs kuhm-pahrt-mehnt
Vitreous humor vih-tre-uhs hyoo-mar
Winr-up Wnd-uhp
INTRODUCTION
The old expression "perception is reality” is nowhere as true as it is for the sensory system. The world that animals, including us humans, live in exists only as it is perceived in their (our) brains. Colors are merely different wavelengths of electromagnetic radiation that stimulate photoreceptors in the eye to send nerve impulses to the brain that are perceived as what we call "colors.” If there is no brain to interpret nerve impulses from the appropriate photoreceptors, colors do not independently exist. So the correct answer to the old question, "If a tree falls in a forest and there is no one (or nothing) present with a sense of hearing, does it make a sound?” would be "No.” What the brain perceives as sound is vibrations of air molecules. If there are no receptors present to detect those vibrations and send impulses to a brain that can perceive them as sounds, sound does not independently exist. This may seem like a strange concept, but it is an important one. The world that the animals we work with live in is determined by the sensory receptors they are equipped with, and how their brains interpret the nerve impulses created by those receptors. The world they perceive is different from the world we perceive — sometimes slightly and sometimes greatly. Awareness of those differences makes it much easier for us to work with them in a useful manner.
Okay, let's get down to basics. How many senses do animals have? Tradition says five: hearing, seeing, feeling, smelling, and tasting. Anatomy and physiology concepts, however, say they have more than that. Coming up with a precise number is difficult, because so many different kinds of sensations can be identified, and the total number depends on how we separate or group them. However, the total is definitely more than five.
Before we sort through the sensory numbers game, let's look at what sense organs are. In simplest terms, they are extensions of the central nervous system (CNS) that allow it to monitor what is going on inside and outside the animal. At the heart of all sense organs are various kinds of specially modified nerve endings (dendrites), called sensory receptors. When triggered by an appropriate stimulus, a sensory receptor generates nerve impulses that travel to the CNS and are interpreted (perceived) as a particular sensation.
The sensory receptors of common domestic animals are sensitive to only four general types of stimulus:
1. Mechanical stimuli (e.g., touch, hearing, balance).
2. Thermal stimuli (e.g., hot and cold).
3. Electromagnetic stimuli (e.g., vision).
4. Chemical stimuli (e.g., taste and smell).
All of the sensations that an animal can perceive start with one or more of these four types of stimuli. Therefore the CNS has to do a lot of work to interpret the resulting sensory nerve impulses correctly. For instance, one type of mechanical stimulus is a pesky cat rubbing against a dog's leg. Another type of mechanical stimulus is the sound of a can opener being used to open a can of dog food. The distinction is important to a hungry dog. Fortunately, its CNS is preprogrammed to perceive the many sensory nerve impulses it receives correctly.
So how many senses are there? Would you believe 10? That's right; we are going to discuss 10 senses, or categories of sensation, in this chapter-five general senses and five special senses. These are listed in Table 10-1.
| I TABLE 10-1 General and Special Senses | ||
| SENSE | WHAT IS SENSED | bgcolor=white>TYPE OF STIMULUS|
| General Senses | ||
| Visceral sensations | Hunger, thirst, hollow-organ fullness | Chemical, mechanical |
| Touch | Touch and pressure | Mechanical |
| Temperature | Heat and cold | Thermal |
| Pain | Intense stimuli of any type | Mechanical, chemical, or thermal |
| Proprioception | Body position and movement | Mechanical |
| Special Senses | ||
| Taste | Tastes | Chemical |
| Smell | Odors | Chemical |
| Hearing | Sounds | Mechanical |
| Equilibrium | Balance and head position | Mechanical |
| Vision | Light | Electromagnetic |
GENERAL SENSES
The general senses are visceral sensations, touch, temperature, pain, and proprioception. Some of them are not exactly household terms, but as their category name implies, general senses are distributed generally throughout the body. Their receptors tend to be fairly simple structures, and they transmit sensory information to the CNS through peripheral and autonomic nerve fibers. Because their receptors tend to be widespread on the inside and outside of the body, the general senses keep the CNS informed about the overall prevailing conditions both inside and outside the body.
Although they are important to the well-being of an animal, with the exception of pain the general senses are rarely involved in clinical disease or treatment. So we will give them a quick once-over, but we will not discuss them in detail. Additional information about the general senses can be found in more in-depth anatomy and physiology references.
VISCERAL SENSATIONS
Visceral sensations make up a somewhat miscellaneous category of interior body sensations. Most are vague and poorly localized. They include the sensations of hunger and thirst, which indicate deficiencies of nutrients and water. The result of such sensations is the initiation of actions designed to secure the needed substances and restore nutrient and fluid balance (homeostasis) in the body.
Other visceral sensations originate in internal organs, particularly hollow organs such as the gastrointestinal tract and portions of the urinary system. Interestingly, these organs have only certain, specific kinds of receptors, particularly stretch receptors. Anything that stretches the wall of the organ, such as a bubble of gas in the intestine or a stone (calculus) in the ureter, can be intensely painful. On the other hand, these same organs can be handled, cut, or crushed without any apparent pain. This can be a problem on those rare occasions when an abdominal surgical incision breaks down (called surgical wound dehiscence) and internal organs come out through the defect. The animal actually can traumatize its own organs without feeling any apparent discomfort.
The urinary bladder is an exception to the rule that stretching causes severe pain, which holds true for most other hollow internal organs. The job of the urinary bladder is to store urine as it is produced by the kidneys and release it periodically to the outside. The sensation of a filling bladder reaches the conscious mind, but it is not an acutely painful event. Rather, it triggers reflex centers in the spinal cord that cause the smooth muscle in the wall of the bladder to contract. Urination may then take place, or it can be delayed if the animal contracts the voluntary sphincter muscle that surrounds the neck of the bladder.
The pleura and peritoneum (membranes that line and cover the contents of the thorax and abdomen, respectively) are well supplied with sensory receptors. As long as conditions are normal and the pleural and peritoneal surfaces slide over each other smoothly, no sensation is felt. However, if the surfaces become roughened by inflammation and/or infection, the resulting pleuritis or peritonitis is very painful. Pleuritis and peritonitis most often result from penetrating wounds from the outside or from ruptures or perforations of internal organs. When opening thoracic or abdominal organs surgically, we must be very careful to suture them closed securely to prevent leakage, which could lead to pleuritis or peritonitis.
TOUCH
We include touch and pressure together, even though they are sometimes classified as separate senses. Touch, also known as the tactile sense, is the sensation of something being in contact with the surface of the body. It can be difficult to differentiate touch from pressure, which is the sense of something pressing on the body surface. Different kinds of specific touch and pressure receptor produce sensations of light contact, deep pressure, vibration, or hair movement. The overall effect is to give the CNS a picture of what, where, and to what extent objects from the outside environment are physically in contact with the surface of the body. The touch and pressure sensations operate almost at an unconscious level, unless the contact is abrupt or the pressure severe. Once physical contact or pressure is initially sensed, it quickly fades into the sensory background unless it changes or is extreme.
If you are sitting down while reading this, take a moment to think about where the chair is contacting your body— probably in the areas of your lower back, your buttocks, and the backs of your thighs. Until you thought about those areas, you probably were not consciously aware of the pressure on them, unless you are sitting in a very uncomfortable chair. The CNS has too much information to process to keep track of everything that is, or is not, in contact with every square millimeter of the skin's surface. Usually, only when things change, or are extreme, do the sensations of touch and pressure rise to the level of the conscious mind.
TEMPERATURE
The temperature sense is the monitoring half of the body's temperature control (temperature homeostasis) system. Temperature receptors detect increases or decreases in body temperature and transmit the information to the CNS. The CNS can activate mechanisms to correct conditions of hypothermia (too low a body temperature) or hyperthermia (too high a body temperature).
Temperature receptors fall into two categories: superficial and central. Superficial temperature receptors are located in the skin and detect upward or downward changes in skin temperature. Heat receptors increase their generation of nerve impulses when the temperature increases, and cold receptors increase theirs when the temperature falls. These increased impulses get the attention of the conscious mind and let it know that things are out of balance. At constant temperatures, the receptors generate steady, low-level streams of nerve impulses that are sensed at the subconscious level and do not intrude on the conscious mind.
Central temperature receptors keep track of the core (interior) temperature of the body by monitoring the temperature of the blood. Central temperature receptors are located in the hypothalamus, a small but very important area of the brain. (See Chapters 9 and 11 on the nervous and endocrine systems for information on other important functions of the hypothalamus.) An animal's rectal temperature indicates its core temperature.
By monitoring the temperature of the body both superficially and centrally, the nervous system can initiate corrective actions if things become too hot or cold. By controlling functions such as blood flow in and beneath the skin, sweating, piloerection (hairs standing on end to increase insulation by trapping air), shivering, and even thyroid hormone production, the nervous system can set in motion mechanisms that can help bring body temperature back into balance. It can also initiate actions to help heat or cool the animal as needed. If the animal is too hot, it may seek out shade or cool water. If it is too cold, it might seek out warmth or increase its muscular activity to generate more heat.
CLINICAL APPLICATION
Heatstroke and Hypothermia
Normal cellular functions in warm-blooded animals depend on the core body temperature remaining fairly constant. This is because chemical reactions, including all the metabolic reactions that occur in the body, are temperature dependent. Higher temperatures speed up chemical reactions, and lower temperatures slow them down. Significant variations in the core temperature of the body, such as might occur in heatstroke (significantly elevated body temperature) or hypothermia (significantly decreased body temperature), can have serious consequences and endanger the life of the animal.
Heatstroke can result from prolonged exposure to high environmental temperatures. The core temperature of the affected animal climbs to dangerously high levels. Early on, the animal typically appears weak and confused; as things progress, it may lapse into unconsciousness that can lead to convulsions and even death. The very rapid heart and respiratory rates that occur in affected animals are signs of the abnormally accelerated metabolic reactions in the body. If the animal is not cooled in time, the distorted metabolic reactions, particularly in the brain, can reach a point at which brain damage and possibly death can result. The maximum body temperature compatible with life is about 10° F (5° C) above the animal's normal body temperature level.
Hypothermia results from an abnormally low body temperature that slows all the metabolic processes. The heart and respiratory rates of affected animals slow as their core temperatures drop. If not warmed, affected animals can lose consciousness and die. Hypothermia can result from prolonged exposure to cold environmental temperatures, but it can also occur in the veterinary hospital in animals under general anesthesia. Most general anesthetic drugs anesthetize the temperature control centers in the brain along with the conscious mind. This often results in a slow fall in body temperature during anesthetic procedures that can be accelerated by contact with cold environmental surfaces, such as metal surgery tables. The falling core temperature slows metabolic reactions in the animal's body, including those that metabolize or eliminate the anesthetic agent at the end of the procedure and allow the animal to wake up. This can prolong the time it takes for the animal to recover from the anesthetic. For this reason, we generally try to keep anesthetized and recovering animals warm through means such as table and cage warmers, towels, blankets, and hot water bottles.
PAIN
Pain receptors, also called nociceptors, are the most common and widely distributed sensory receptors inside and on the surface of the body. They are found almost everywhere. They range from simple, free nerve endings that respond to intense stimuli of all types, to more specialized structures that detect mechanical forces, temperature, and so on. Their purpose is to protect the body from damage by alerting the CNS to potentially harmful stimuli. Interestingly, the only place in the body where pain receptors are not found is the brain. It is not uncommon for certain types of human brain surgery to be performed on wide-awake patients who have had local anesthesia to allow the brain to be exposed.
The process of experiencing pain is called nociception. It seems like it should be a straightforward, simple process. A painful stimulus should generate a sensory nerve impulse that goes to the brain and is perceived as pain, right? Actually the process is more complicated than that, and our growing understanding of the processes involved is enabling us to provide more effective pain management to veterinary patients.
Four processes contribute to nociception (Figure 10-1). Some fit our logical preconceptions, and at least one adds some new wrinkles to our thinking. The first step in nociception has the strange name of transduction. This is the conversion of the painful stimulus to a nerve impulse, which occurs at the sensory nerve ending. Transmission of the nerve impulse up the sensory nerve fibers to the spinal cord is the next step. So far, everything is as we expected. The next step in the process, however, has only recently been fully appreciated. The spinal cord does not simply relay the sensory nerve impulses up to the brain. Modulation (changing) of the sensory nerve impulses can occur in the spinal cord, and this can significantly influence the information the brain receives, particularly in cases of chronic or severe pain. This modulation process can amplify (make more severe)
FIGURE 10-1 Pain pathways.
or suppress (make less severe) sensory impulses through synapses between neurons in the dorsal horns of the spinal cord. We can influence this modulation process through therapy with several different classes of drugs. This can enhance our ability to prevent and treat chronic and severe pain in our patients.
The last step in the nociception process is perception of the painful impulses by several areas of the brain. Conscious perception occurs in the cerebral cortex, but other areas of the brain are involved also. These include areas involved with the autonomic nervous system (fight or flight), fear and anxiety, memory, arousal, and behavior and emotion. Severe or chronic pain can have far-reaching negative effects on an animal's well-being that can extend beyond just the unpleasant conscious discomfort of pain.
Pain can be classified in a number of ways. One useful system classifies pain as superficial (affecting the skin and subcutaneous areas), deep (involving muscles and joints), and visceral (relating to the internal organs). Another system classifies pain as acute (sharp and intense) or chronic (dull and aching).
We cannot get inside the head of a dog or a horse to find out precisely how they perceive pain, but behavioral and physiologic responses suggest that they experience pain much the same way we do. The difference is often in how they react to the pain. We humans often tend to dwell on pain we are experiencing, which can make the overall experience even more stressful, whereas nonhuman animals seem to accept the current situation as how things are supposed to be. This is not to say that they do not suffer pain; rather, they just do not seem to have the same kind of emotional reaction to it that we humans do. They often seem to hide it well, which can be a problem for us clinically when we are trying to assess the degree of pain a patient is experiencing. Actually, hiding signs of pain is a survival instinct for most animals. An animal that shows signs of pain is showing signs of weakness that might encourage other animals, including predators, to attack it.
Visible reactions to pain vary greatly among species, breeds, and individual animals. Some animals are very sensitive to pain and become stressed by even mildly painful stimuli. Others hardly seem to react at all until pain becomes severe. Prevention and relief of pain are becoming increasingly important in veterinary medicine, as we learn more about its harmful effects and how to prevent them. Research studies are providing the knowledge necessary to detect the subtle signs of pain that many animals show. This information is allowing us to become more proactive in our efforts to prevent and treat pain in ill and injured patients.
PROPRIOCEPTION
Without looking at them, can you tell what positions your arms and legs are in? Of course you can, although precisely how you do it may not be clear. You just seem to know where all your body parts are. Actually, you are making use of your sense of proprioception, which is the sense of body position and movement. This sense operates largely at the subconscious level and is very important in allowing an animal to stand upright and make accurate, purposeful movements as it interacts with its environment. During the examination of an animal with suspected nervous system damage, a veterinarian may evaluate proprioception by curling the animal's foot under so that it is upside down, and seeing how long it takes the animal to right it. Animals with normal pri^o^^^^^ti011 almost immediately bring the foot oup int a normal standing position.
The heart of the proprioception sense is the variety of stretch roeccaetpedtors l in skeletal muscles, tendons, ligaments, and joint capsules. These receptors keep the CNS informed about the move iτιents of limbs, the positions of joints, the state of contraction of muscles, and the amount of tension being exerted on tendons and ligaments. This information is important to the CNS so that it can send out the right combination eorftvomer n impulses, which are appropriate in range and
sotrength, t produce smooth body movements.
SPECIAL SENSES
The special senses include four of the traditional senses— taste, smdl, hearing, and vision—plus the important sense of equilibrium. In contrast with the general senses, the special senses are organized into specific, often complex rsegnansosry o and structures that are all located in the head. Because of their locations, structures, and functions, the snpsecial se organs often are involved in clinical illnesses aunridesi.nj
TASTE
The sense of tastel also called the gustatory sense, is a dιem- itncssael. se I receptors are located in the mouth in structures called taste buds. Wfen they detect chemical substances dissolved in the saliva, the taste receptors generate nerve impulses that travel to the brain and are interpreted as tastes.
∕j CLINICAL APPLICATION
Anesthesia and Analgesia
The aMty to perceive sensations, or to feel things, is known as esthesia. (The study of the sensory system is called esthesiology.) Anesthesia is the loss of eerheeia, or the complete loss of sensation. In clinical veterinary medicine, we use two basic types of aoeerheeia to carry out procedures that would be painful for patients: general anesthesia and local anesthesia.
General anesthesia involves a complete loss of sensory perception accompanied by loss of consciousness. The animal icosedpla int a controlled sleep that prevents it from feeling painful procedures. We produce this sleep by administering general anesthetic drugs either by injecting them or by having raenaitmheals b them from an inhalant anesthesia machine. Animals under general anesthesia must be monitored closely, because the drugs depress cardiovascular and respiratory functions along with the CNS.
Local anesthesia parises loss of sensation from a specific, localized area of the body without affecting consciousness. It irsodpuced by injecting a local anesthetic drug into an area thhicrohugh w sensory nerve fibers pass. The drug blocks the transmission of nerve impulses through the site, which pnrseovreynts se information from reaching the central nervous system. This can allow potentially painful procedures to bm performed without having to render the animal unconscious.
Analgesia is a related state in which the perception of pain iescrdeased but not completely absent. The pain is dulled but onmotpcletely gone. A drug that produces analgesia is called an analgesic drun, Aspirin, carprofen (Rimadyl1'), and morphine are all examples. Analgesic drugs are often used to make animals with severe pain (postsurgical patients for example) more comeortable.
∕j CLINICAL APPLICATION
Preventing Wind-Up
seoWelidevue t b that potentially painful procedures we carried
nut on an animal under general anesthesia, such as surgery, hnd no lgng-lasting effects. After all, properly administered gnenstehreatlicas effectively block the perception of painful
iympulses b the conscious part of the brain. We now know that the other taee nociceptive processes—transduction, transmission ano modu Iation—are still operating full-bore in an anesthetized animal during surgery, and they can significantly affect the animal's Wel of pain once they wake up from the anesthetic.
Mosn important, we now understand that the neurons of tohred spinal c are bombarded with painful stimuli during srourcgedicuarleps despite the fact that the conscious mind iesmtporarily disconnected from the process by the general anesthetic. Because the spinal cord is capable of changing (modulating) the information it forwards on to the brain, this sensory assault during surgery can cause the pain signals going troainthe b to be amplified once the animal wakes up from
tehne rgal anesthesia. This can make the animal's postopera-
tvievle pain le even more severe than the tissue damage caused buyrgtehrey s would seem to warrant. This exaggerated pain response is referred to as wind-up, and it can cause significant sntress o a postsurgical patient.
sWind-up i much easier to prevent than it is to treat. It is odifficult t bring the exaggerated pain response produced by wind-up under control with drugs. On the other hand, if we ceacrneadse the painful stimuli received by the spinal cord duurgreinryg, s we can often prevent wind-up from developing lirhansectet. fi p This can be done by administering analgesic aonssdibply local anesthetic drugs before, and even during, surgery. Using small amounts of several drugs with different analgesic/anesthetic mechanisms is often more effective than aursgineg a l dose of a single drug. This strategy of heading off the effects of severe surgical pain before it occurs can make pain cuorinntgrol d the postoperative period much more effective, tactnudalilyc a speed a patient's recovery from surgery.
TEST YOURSELF 10-1
1. Why are visceral sensations important to the survival of an animal?
2. Why do touch and pressure sensations fade so rapidly from the conscious mind unless they change or are severe?
3. If a dog walks out of an air-conditioned house and lies in the sun on a hot summer day, which of its temperature receptors will signal the brain first that the dog is getting hot: superficial receptors or central receptors?
4. Which category of temperature receptors is most critical to the long-term survival of an animal in very hot or very cold environmental conditions: the superficial receptors or the central receptors? (Hint: Which is more critical to an animal's survival-keeping its skin and extremities from getting too hot or cold or keeping the core of its body from getting too hot or cold?)
5. What is nociception and what roles do transduction, transmission, modulation, and perception play in it?
6. Why do you suppose mild to moderate pain often does not appear to affect the mood or behavior of domestic animals significantly?
7. Why is the proprioceptive sense so important to the maintenance of balance and an upright posture? a huge amount of information from sniffing the air or an object such as a fire hydrant (this can be thought of as reading their “pee-mail”). For reasons we will discuss shortly, dogs probably do not see colors like we do, but they can smell “colors” that we cannot even imagine. So the next time you are walking a dog and it decides to stop and do a thorough sniff-examination of something, do not immediately yank on its leash to get it moving. It is busy reading messages left for it by every other dog that has visited there and left a pee-mail message. Take a moment to appreciate the amazing communication process you are witnessing. The dog will appreciate it, and may add a “reply to all” message of its own by leaving a small amount of urine!
The sense of smell is organized in two patches of olfactory epithelium located up high in both nasal passages. Figure 10-3 shows the location and some of the structure of the olfactory epithelium. Sensory (olfactory) cells are mixed with supporting cells in these epithelial patches. Hairlike processes (modified dendrites) from the surfaces of the olfactory cells project up into the mucous layer that covers the nasal epithelium. When odor molecules dissolve in the mucus and contact the sensory processes, nerve impulses are generated that travel to the brain and are interpreted as particular smells.
The majority of the taste buds are located on the sides of certain small, elevated structures on the tongue called papillae, although a few can be found in the lining of the mouth and throat (pharynx). As seen in Figure 10-2, taste buds are tiny, rounded structures made up of gustatory (sensory) cells and supporting cells. Tiny openings on the surface of each taste bud, the taste pores, allow dissolved substances to enter the taste buds and contact the sensory receptors. The sensory receptors are tiny, hairlike processes (modified dendrites) from the gustatory cells that project up into the taste pores. When appropriate chemical substances dissolved in the saliva come in contact with the sensory processes, nerve impulses are generated that travel to the brain and are interpreted as particular tastes.
In humans the four primary taste sensations are sweet, sour, salty, and bitter. Each has a particular area of the tongue that responds most strongly to that taste. The ability to detect many different kinds of taste is due to combinations of the four basic taste sensations and interactions with the sense of smell. We cannot know if nonhuman animals experience tastes in the same way, but taste sensations most likely vary among animal species.
SMELL
The sense of smell is also called the olfactory sense. It is a chemical sense very similar to taste. The sense of smell is more important in most nonhuman animals than it is in humans. We live in a sight-oriented world. Although we have a decent sense of smell, our keen vision is more important to us as we interact with our environment. In contrast, many nonhuman animals have less sensitive eyesight but a highly sensitive sense of smell. They live in more of a smell-oriented world. This can be difficult for us to relate to, but dogs obtain
TEST YOURSELF 10-2
1. How are the senses of taste and smell similar? How are they different?
2. Why do nonhuman animals often greet others by sniffing them?
HEARING
Hearing, also called the auditory sense, is a mechanical sense that converts vibrations of air molecules into nerve impulses that are interpreted by the brain as sound. The ear, the organ of hearing, can be divided into three physical and functional areas: the external ear, the middle ear, and the inner ear. The external ear acts as a funnel to collect sound wave vibrations and direct them to the eardrum. The middle ear amplifies and transmits the vibrations from the eardrum to the inner ear. The inner ear contains the actual sensory receptors that convert the mechanical vibrations to nerve impulses, along with receptors for the equilibrium sense.
Most of the ear structures are housed within the temporal bones of the skull. The external ear canal, the middle ear cavity, and the inner ear structures all occupy hollowed-out areas in the temporal bones that are lined with soft tissue membranes. The processes of collecting, transmitting, and converting sound wave vibrations all take place within these membrane-lined bony cavities. Figure 10-4 shows the main structures of the external, middle, and inner portions of the ear.
EXTERNAL EAR
The external ear consists of structures that collect sound waves and transmit them to the middle ear. Its main parts
FIGURE 10-2 Taste buds onthe tongue. A, One magnified papilla showing locations of taste buds. B, Taste bud structure. C, Tongue showing a few of the many papillae containing taste buds.
∕j CLINICAL APPLICATION
Upper Respiratory Tract Infections
Upper respiratory tract infections are caused by disease-causing organisms, such as viruses and bacteria. They affect primarily the nasal passages and pharynx (throat) and result in coughing, sneezing, sore throat, and discharges from the eyes and nose. The common cold is a human upper respiratory tract infection. What: sometimes makes upper respiratory tract infections dan- orrous for domestic animals is their effect on the animals’ sense of sme 11—these infections effectively eliminate it. Think back to your last cold. You probably did not have much of a sense of smell, which rendered most foods bland and tasteless.
The effect on animals that live in a smell-oriented world is reeven mo drastic. They often stop eating and drinking completely, because they cannot smell anything. If this continues fenorgry, v lo they can be in real danger from dehydration. Therefore we often have to administer fluids, either orally or by injection, to animals with upper respiratory tract infections. By keeping them properly hydrated while we provide coetshsearryne medical and nursing care, we often can help them fight off the infection.
are the pinna, the external auditory canal, and the tympanic membrane (eardrum).
The pinna is the part of the ear that we can see from the outside. It is a funnel-like structure composed mainly of elastic cartilage and skin that collects sound wave vibrations and directs them into the external auditory canal. In many animals, the pinna is very mobile and can be aimed in the direction of a sound. For instance, watch ftshe ear o a horse in a strange environment. Its ears will eprobably b scanning the area like a couple of little radar
FIGURE 10-3 Olfactory region of the dog. A, Location of olfactory region high ln nasal passage. B, Olfactory epithelium. C, Detail of olfactory epithelium showing olfactory cilia projecting up into overlying mucous layer.
dishes. This ability is particularly useful in animals with erect ears.
The external auditory canal is a soft, membrane-lined tube that begins at the base of the pinna and carries sound waves to the tympanic membrane (eardrum). In most domestic animal species, it is somewhat L-shaped with an outer, vertical portion and an inner, horizontal portion. It ends blindly at the tympanic membrane.
The tympanic membrane is commonly called the eardrum. I- is a paper-thin connective tissue membrane that is tightly stretched across the opening between the external auditory canal and the middle ear cavity. When sound wave vibrations strike it, the tympanic membrane vibrates at the same frequency through a process called sympathetic
vibration. (Author’s note: Another example of sympathetic vibration is the annoying audible vibration of a snare drum tchcuatr so when certain notes are played by a band or
forchestra. I you have ever been in a band or orchestra you raorebapbly familiar with that sound.)
MIDDLE EAR
The middle ear cavity is a hollowed-out area of the temporal bone that is lined by soft tissue membranes. It is filled with air and contains three small bones called ossicles and the opening of the Eustachian tube, which connects it with the pharynx (throat). Laterally it is separated from the external ear by the tympanic membrane, and medially it is separated
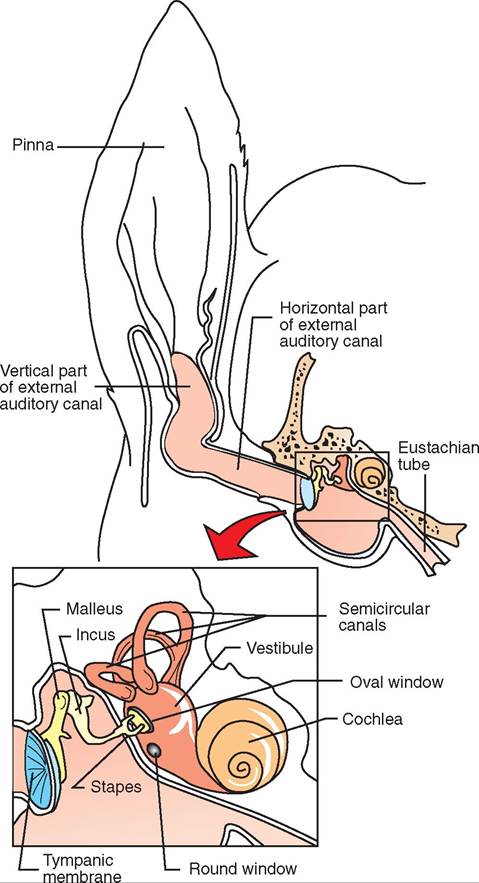
FIGURE 10-4 Cross section ofdog'rscsar structures with middle and inner ear regions enlarged.
from the inner ear by the membranes that cover the oval and round windows of the cochlea.
Three small bones called ossicles link the tympanic membrane with the cochlea of the inner ear, where the receptors for hearing are located. The individual ossicles are named for some past anatomist's fanciful impression of their shapes. The outermost bone, the malleus IhammCT), is attached to the tympanic membrane. The malleus forms a complete synovial joint with the middle bone, the incus (anvil), which in turn forms a joint with the medial-most bone, the stapes (stirrup). ⅞e other end of the stapes is attached to the membrane that covers the oval window of the cochlea. How this attachment to the cochlea contributes to hearing is described further in the section on the inner ear.
lTehs e ossic act as a system of levers that transmit the svoeund wa vibrations from the tympanic membrane to the cochlea. hi doing so, they decrease the amplitude (size) of itbhreatvions but amplify their force. This helps transmit itbhreavtions accurately to the supersensitive receptor structures in the cochlea, ideally without causing any damage. Another mechanism that helps prevent damage to the hearing receptors is a tiny muscle, the tensor tympani, that attaches to the malleus. It adjusts the tension of the tympanic membrane and helps deaden the transmission of extremely loud sound vibrations to the cochlea. Another tiny muscle, the stapedius, assists the damage-control process by restricting the movement of the stapes in response to loud sounds.
The Eustachian tube, also called the auditory tube, connects the middle ear cavity with the pharynx. Its purpose is to eqnelize the air pressure on the two sides of the tympanic membrane. Without this structure, every time the atmospheric (barometric) pressure changed (as it often does; watch the next weather forecast), the tympanic membrane uwlogue ld b in or out, depending on whether the pressure
irncreased o decreased. This would be painful, because the tympanic membrane is liberally supplied with pain receptors, and it would decrease sound wave transmission. Fortunately, the Eustachian tube comes to the rescue. The slitlike fopening o the tube in the pharynx is stretched open whenever the animal swallows or yawns. This allows air to enter or leave as necessary to equalize the pressure in the middle ear cavity with that of the outside air. (Note how this system itos pu t the test when we engage in activities that rapidly change the pressure on the outside of the tympanic membrane, such as flying or scuba diving. Consciously swallowing on yawning often helps correct the resulting pressure imbalance.)
CLINICAL APPLICATION
Ear Hematomas
The pinna of the ear consists of elastic cartilage and small blood vessels covered by skin. Sometimes irritation in the ear canal (such as with otitis externa) will cause an animal to shake its head vigorously (see the Clinical Application on otitis externa). Io some animals, particularly floppy-eared dogs, this movement can rupture small blood vessels under fthe skin o the pinna, usually on the inside surface. The resulting bleeding between the cartilage and skin can cause an scchs^o of blood, called an ear hematoma, which is
anno rambal accumulation of free blood between the carti
lage and skin of the pinna.
Amnatoemara he usually is not dangerous or even
painful, but it is heavy and swollen and often seems to bother the animal. If left untreated, the blood will be reabsorbed sultowly, b the ear probably will be permanently deformed by scar tissue, resulting in what is called a cauliflower ear. Treatment of ear hematomas usually involves surgically draining the material from the hematoma and placing sutures through the pinna to hold the skin tight against the cartilage and prevent fluid from reaccumulating. The underlying cause of the head shaking needs to be determined and treated also to prevent recurrence.
CLINICAL APPLICATION
Otitis Externa
Otitis externa is an inflammation of the skin of the external ear canal that occurs most commonly in dogs, cats, and rabbits. It is often caused by parasites such as ear mites, foreign bodies, or microorganisms such as bacteria and yeasts. The irritation in the ear canal causes redness, pain, itching, and fluid accumulation. The owner usually notices that the animal shakes its head a lot and spends time scratching at its ears. When the affected ear is examined, the ear canal is usually red, moist, swollen, and painful, and often has a characteristic pungent odor.
The basic anatomy of the external ear canal adds a challenge to the treatment of otitis externa. Gravity makes inflammatory fluids drain downward and accumulate in the horizontal portion of the canal next to the tympanic membrane. Because topical medications are usually part of the treatment of otitis externa, the ear canals must be cleaned thoroughly and carefully to remove discharges before the medications are instilled. Because the ear canals are often swollen and painful, this process can be difficult for at least the first few days. Therapy for otitis externa often must be continued for many weeks to bring the condition under control.
INNER EAR
The inner ear is made up of structures that contribute to both hearing and equilibrium. The hearing portion of the inner ear is contained in a snail shell-shaped spiral cavity in the temporal bone called the cochlea (Figures 10-4 and 10-5). Within the hollowed-out bony cavity of the cochlea is a soft, multilayered, fluid-filled portion that contains the receptor organ of hearing—the organ of Corti. The organ of Corti runs the length of the cochlea in a long tube called the cochlear duct, which is filled with a fluid called endolymph. A U-shaped tube containing another fluid, perilymph, lies on either side of the cochlear duct. Membrane-covered openings at the ends of the “U,” called the oval window and the round window, are located at the base of the cochlea. The bottom of the U is located at the tip of the cochlea. Nothing lies against the round window, but the stapes (one of the ossicles) is attached to the oval window.
The organ of Corti runs along the cochlear duct, on a shelf called the basilar membrane, like a long ribbon. Its main parts are hair cells, supporting cells, and the tectorial membrane. The hair cells are the receptor cells of hearing. They have tiny, hairlike projections (modified dendrites) on their surfaces. The gelatin-like tectorial membrane lies gently on top of the hairs like a long, soft strip lying on top of a broad series of brush bristles. As their name implies, the supporting cells provide physical support to the hair cells.
Sound wave vibrations cause the tympanic membrane and the ossicles in the middle ear to vibrate. As the stapes vibrates back and forth, it alternately pushes and pulls on the membrane covering the oval window of the cochlea. This causes the perilymph around the cochlear duct to vibrate back and forth (the membrane covering the round window acts as a pressure relief mechanism by alternately bulging in and out as the fluid moves back and forth). This process is summarized in Figure 10-6. The movement of the perilymph causes the cochlear duct to move, which causes the tectorial membrane and the hair cells of the organ of Corti to rub against each other. This bends the sensory hairs, which generate nerve impulses that travel to the brain and are interpreted as sound.
Different frequencies of sound wave vibration stimulate different areas along the length of the organ of Corti. Areas near the oval window respond best to high-frequency (high-pitched) sounds, and areas at the tip of the cochlea respond best to low- frequency (low-pitched sounds). This physical process generates nerve impulses in different parts of the organ and helps the brain differentiate high- and low-pitched sounds.
TEST YOURSELF 10-3
1. Which skull bone houses the middle and inner ear structures?
2. Why would keen hearing be important to the survival of a potential prey animal? Why would it be important to a predator?
3. How would the rupture or perforation of an eardrum affect hearing?
4. How would arthritis in the tiny joints of the ossicles affect hearing? Could this possibly affect the hearing of older animals?
5. How would an animal probably feel if they had a middle ear infection that caused the opening of the Eustachian tube to swell closed?
6. How might repeated exposure to loud sounds lead to progressive hearing loss?
CLINICAL APPLICATION
Recovery from General Anesthesia
The importance of head position to overall body posture and balance can be seen in an animal that is recovering from general anesthesia. One of the first things that a recovering animal tries to do as it regains consciousness is to raise its head into an upright position and steady it there. It has to get its head into that position before it can start trying to raise its body. Primitive instincts for survival prompt many animals to try and stand before they are steady and coordinated enough to support themselves. This can lead to stumbling and falling, which can cause injury.
By taking advantage of our knowledge of the sense of equilibrium, we can prevent an animal from trying to get up prematurely by gently holding its head down in a horizontal position until it has enough strength and coordination to rise. This can be particularly important for larger animals, such as horses. By holding a recovering horse’s head down with a gentle hand or knee on the dorsal part of its neck just behind the skull, we can keep the animal from trying to get up too soon. Once we feel it has regained enough strength, we can allow it to raise its head and prepare to help steady it as it rises to its feet.
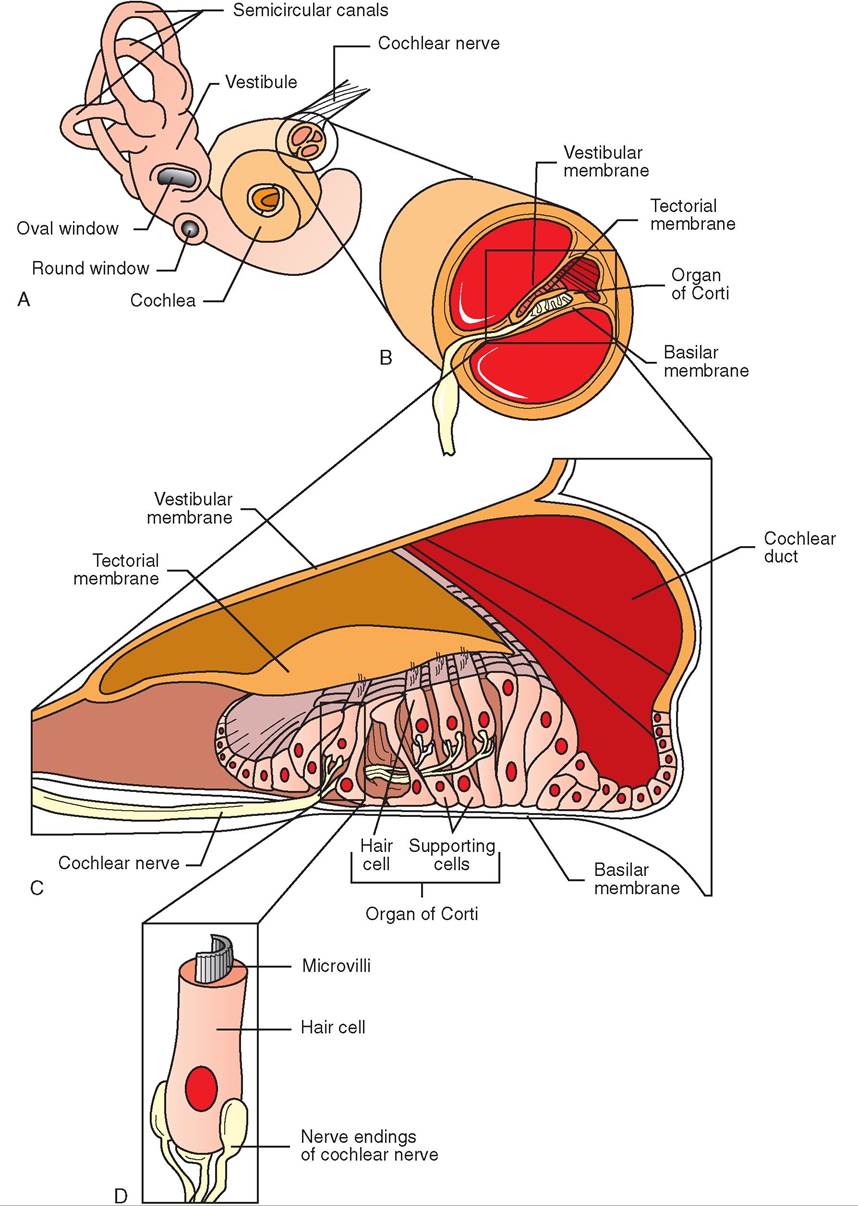
FIGURE 10-5 Inner ear strudures,∖t^ith detailofcocHear an atomy. A, Inner ear structures. B, Enlarged cross section of cochlea. C, Detail of organ of Corti. D, Greatly enlarged, single sensory hair cell.
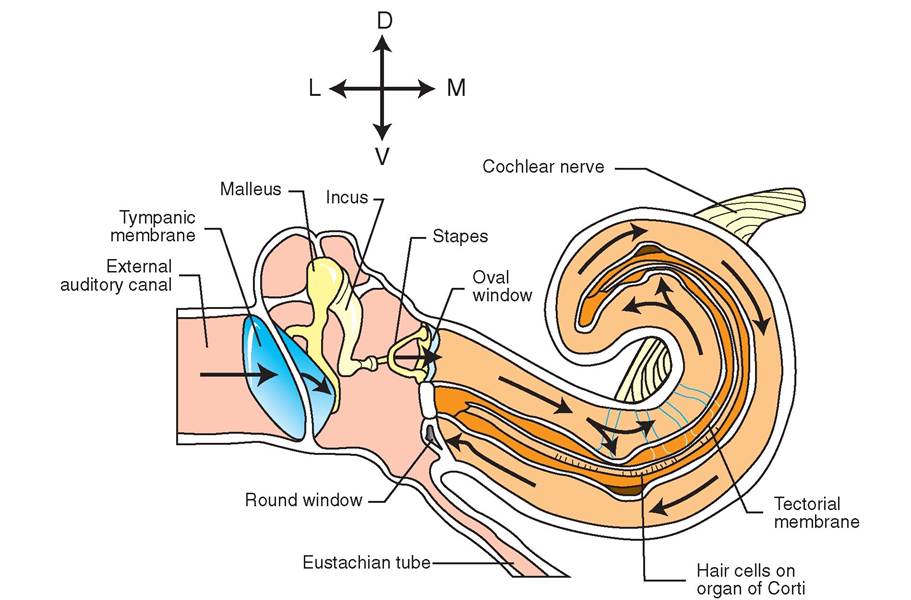
FIGURE 1 0-6 Effect of sound waves on cochlear structures. Sound waves cause the tympanic membrane and ossicles to vibrate. As the stapes vibrates back and forth, it pushes and pulls on the membrane covering the oval window of the cochlea. This sets the fluid in the cochlea in motion, which causes the hair cells and tectorial membrane to rub against each other. This bends sensory hairs, generating nerve impulses that are transmitted to the brain and interpreted as sound.
EQUILIBRIUM
As the head goes, so goes the rest of the body. At least that is the principle behind the sense of equilibrium. This mechanical sense helps the animal maintain its balance by keeping track of the position and movements of the head. The receptors are located in portions of the inner ear called the vestibule and the semicircular canals.
Actually, maintaining balance is a complicated process that involves information from the equilibrium receptors, as well as from the eyes and the proprioceptors around the body. (See the Clinical Application on motion sickness for an explanation of what happens when the information from these various sources does not agree.)
VESTIBULE
The vestibule is the portion of the inner ear that is located between the cochlea and the semicircular canals. It is made up of two saclike spaces, called the utricle and the saccule, that are continuous with the cochlear duct of the cochlea and are filled with the same endolymph fluid. Like the cochlear duct, the utricle and saccule are surrounded by perilymph.
In each utricle and saccule is a patch of sensory epithelium called the macula (Figure 10-7). It consists of hair cells and supporting cells covered by a gelatinous matrix that contains tiny crystals of calcium carbonate called otoliths. (The word otolith literally means “ear stone.”) The hair cells here are similar to the hair cells of the organ of Corti in the cochlea. They have hairlike processes (modified dendrites) on their surfaces that the gelatinous matrix sits on. Gravity causes the otoliths and the gelatinous matrix to put constant pressure on the hairs as long as the head stays still. Movement of the head bends the sensory hairs, which generates nerve impulses that give the brain information about the position of the head.
Semicircular canals
The semicircular canals are located on the other side of the vestibule from the cochlea. Each canal is semicircular and oriented in a different plane at right angles to the other two. If you are indoors in a room with square walls, look up in a corner where two walls and the ceiling come together. Each of the walls and the ceiling is in a different plane, at right angles to the other two. This is the basic arrangement of the semicircular canals.
Within each bony semicircular canal is an endolymph- filled membranous tube that is surrounded by perilymph. (Each of the endolymph-filled structures in the inner ear is continuous with the others, as are the perilymph- filled structures.) Near the utricle end of each semicircular canal is an enlargement, called the ampulla, that contains the receptor structure, called the crista ampullaris, or simply the crista.
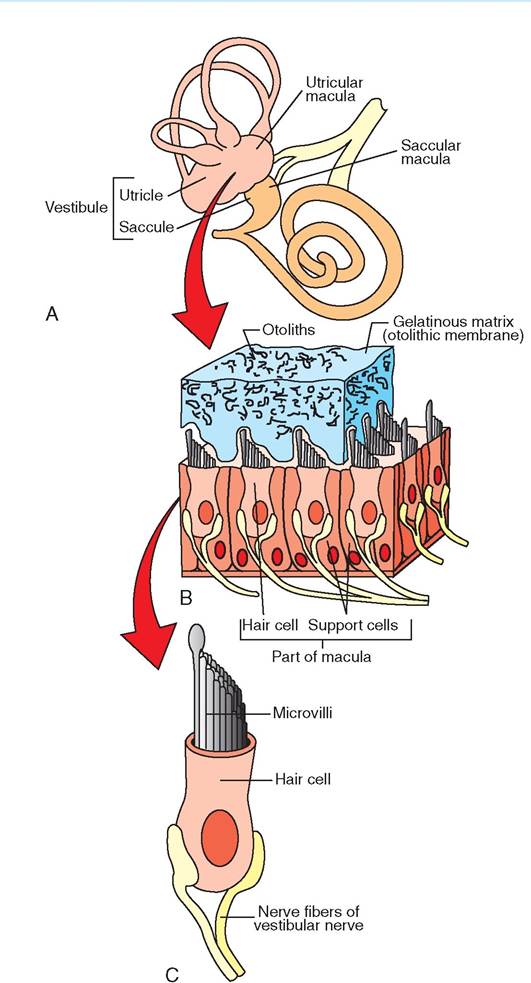
FIGURE 10-7 Inner ear structures, with detail of vestibular anatomy.
A, Inner ear structures. Note the utricle and saccule, which make up the vestibule, and locations of the macula (sensory epithelium) in each.
B, Enlarged view of macula showing sensory hair cells, gelatinous matrix, and otoliths. C, Greatly enlarged sensory hair cell.
The crista is similar to the macula of the vestibule. It consists of a cone-shaped area of supporting cells and hair cells with their processes (modified dendrites) sticking up into a gelatinous structure called the cupula (Figure 10-8). However, there are no otoliths to weigh down the cupula. It functions as a float that moves with the endolymph in the membranous canal.
When the head moves in the plane of one of the semicircular canals, inertia causes the endolymph to lag behind the movement of the canal itself. The same principle is at work if you quickly rotate a glass containing liquid and ice cubes.
FIGURE 10-8 Semicircular canals, with detail of ampulla and crista ampullaris. A, Semicircular canals, showing locations of ampullae. B, Enlarged view of crista ampullaris in an ampulla, showing sensory hair cells and cupula. C, Greatly enlarged sensory hair cell.
The glass turns, but the liquid and ice cubes lag behind. The relative movement of the endolymph pulls on the cupula, which bends the hairs. This generates nerve impulses that give the brain information about motion of the head, particularly rotary motion.
So the vestibular system senses rotary motion of the head with the semicircular canals, and linear motion and the position of the head with the vestibule. By integrating this information, the brain forms a picture of what is happening to the animal's head and, by extension, to its body as a whole.
TEST YOURSELF 10-4
1. How is the functioning of the vestibule and the semicircular canals similar? How is it different?
2. What are otoliths and why are they important to the equilibrium sense?
3. What is the difference between ossicles and otoliths?
4. How is the physical concept of inertia important to the functioning of the semicircular canals?
5. What is the basic cause of motion sickness?
VISION
The eyes have a lot in common with digital cameras. They have lens covers (the eyelids), a “window” on the front to let light in (the cornea), an adjustable diaphragm to control the amount of light that enters (the iris), a lens that can be focused, an image sensor on which the image is formed (the retina), and connections (the optic nerves) to carry the images to a memory device (the brain) for storage.
As complicated as the eye seems, most of its components exist to help form an accurate visual image, not to detect one. The actual photoreceptors that detect the image and generate visual nerve impulses are in a single layer of cells in the retina (the structure that lines the back of the eyeball). In our discussion of vision, we'll deal mostly with the imageformation structures of the eye.
Terminology
Two general terms are used to refer to the eye: the word ocular and the combining form ophthalm/o. For example, ocular anatomy refers to the anatomy of the eye, ophthalmic medications are used to treat the eye, and ophthalmology is the study of the eye.
MAJOR LAYERS OF THE EYEBALL
The eyeball consists of three major layers: the outer fibrous layer, the middle vascular layer, and the inner nervous layer. Figure 10-9 illustrates the main structures of each layer.
FIBROUS LAYER. The outer, fibrous layer of the eye admits light to its interior and gives strength and shape to the eyeball. Its two components are the cornea and the sclera. The cornea is the transparent “window” that admits light to the interior of the eye. It consists of an orderly arrangement of collagen fibers and contains no blood vessels. Its transparency is maintained by careful control of the amount of water it contains. Too much water (corneal edema) or too little water (corneal dehydration) causes the cornea to become cloudy and opaque. The cornea is richly supplied with pain receptors, making it one of the most sensitive tissues of the body.
CLINICAL APPLICATION
Motion Sickness
The brain relies on the vestibular system, the eyes, and the proprioceptors around the body for information about how and where the body is moving so that it can keep the animal upright and maintain balance. Usually the information the brain receives from these sources agrees—the animal is either moving, or it is not. This is fine as long as it is standing or moving around on the ground (assuming the ground is not moving). But when we put an animal into a moving car, boat, airplane, or space vehicle, things become more complicated. The eyes look around the interior of the vehicle and see that nothing is apparently moving, but the equilibrium receptors and proprioceptors detect motion (or, in the case of space travel, the absence of gravity).
This disagreement between the sensory receptors can result in the unpleasant sensations of motion sickness, such as headache, nausea, and vomiting. This often occurs in animals that travel by car, truck, boat, or plane. Medications often can be used to prevent motion sickness, but they usually must be administered before traveling. Motion sickness is easier to prevent than it is to treat in mid journey. Veterinary clinics may dispense anti-motion-sickness medications to clients who must travel with animals that are prone to motion sickness.
A form of motion sickness is common in astronauts as well. Space adaptation syndrome occurs in a high percentage of astronauts, particularly during the first few days of a space flight. It causes symptoms similar to normal motion sickness, such as headache, nausea, and cold sweats. The symptoms often disappear after a day or two, indicating that the brain has adjusted to the conflicting signals. Ongoing research is focused on finding ways to prevent this annoying and sometimes debilitating disorder. It seems more complicated to deal with than traditional earth-bound motion sickness.
The sclera is the “white” of the eye. Like the cornea, it consists mainly of collagen fibers and makes up the majority of the outer fibrous layer of the eye. The junction of the cornea and the sclera is called the limbus. It can be used as a landmark to describe the position of lesions (abnormalities) on the cornea or sclera.
VASCULAR LAYER. The middle, vascular layer is also called the uvea. It has several parts, including the choroid, the iris, and the ciliary body.
The choroid is sandwiched between the sclera and the retina. It consists mainly of pigment and blood vessels that supply blood to the retina. Most of the pigment is dark melanin, but in most domestic animals, except swine, the choroid forms a highly reflective area in the rear of the eye called the tapetum lucidum or, more commonly, the tapetum. (See Figure 10-12.) The tapetum is like a brightly colored mirror, and it is responsible for the bright light that seems to shine from an animal's eyes in the dark, when a light is directed into them. Its purpose seems to be to act as
FIGURE 10-9 Sagittal section of an eye.
for vision, the rods and cones. (We discuss the retina in more detail shortly.) The term fundus is sometimes used to describe the caudal interior surface of the eye. The retina is the main component of the fundus, along with the optic disc, which we discuss below.
TEST YOURSELF 10-5
1. An animal has an area of inflammation located at the dorsal limbus of its right eye. Where is the lesion located?
2. The cornea and sclera are both made up substantially of collagen fibers. Why is the cornea transparent, but the sclera opaque?
3. Which layer of the eyeball are the iris and ciliary body part of?
4. How does the tapetum aid low-light vision?
5. What is the function of the iris?
6. Which layer of the eyeball contains the photoreceptors?
a light amplifier to aid low-light vision. After light has passed through the photoreceptors (the rods and cones in the retina), it reflects off the tapetum and passes back through the photoreceptors again, stimulating them a second time. Therefore most animals can see better in dim light than we can. Humans and pigs do not have a tapetum.
At the front of the eye, the middle vascular layer is modified into the iris and the ciliary body. The iris is the colored part of the eye. If we speak of an animal as having blue eyes, it is the iris color we are talking about. The iris is a pigmented muscular diaphragm that controls the amount of light that enters the posterior part of the eyeball. The opening at its center is called the pupil. The pupil enlarges in low-light conditions and gets smaller in bright light. Two types of multiunit smooth muscle fiber make up the iris: radially arranged fibers (oriented like the spokes of a wheel) that enlarge the pupil when they contract, and circularly arranged fibers that constrict the pupil when they contract. The nerve supply for the smooth muscle cells of the iris comes from the autonomic nervous system.
The ciliary body is a ring-shaped structure located immediately behind the iris. It contains the tiny muscles that adjust the shape of the lens to allow near and far vision. The muscles of the ciliary body (the ciliary muscles) are also multiunit smooth muscles. They are contained within small processes (called the ciliary processes) that are attached to the periphery of the lens by tiny suspensory ligaments. The ciliary muscle fibers are oriented so that when they are relaxed the suspensory ligaments pull on the periphery of the lens, stretching it into a flattened shape. When they contract, they move the ciliary body forward and inward. This action takes tension off the suspensory ligaments, allowing the lens to assume its natural, more rounded shape. (We discuss this focusing process more in the section on the lens.)
NERVOUS LAYER. The inner, nervous layer is the retina, which lines the back of the eye. It is like the image sensor in the camera of the eye. It contains the actual sensory receptors
MAJOR COMPARTMENTS OF THE EYEBALL
The interior of the eyeball is made up of two fluid-filled compartments: one in front of the lens and ciliary body and the other behind it. The aqueous compartment is in front of the lens and ciliary body and contains a clear, watery fluid called aqueous humor. The vitreous compartment is behind the lens and ciliary body and contains a clear fluid with the consistency of soft gelatin, called vitreous humor. The term humor is an old anatomical term meaning fluid. The eye is the only part of the body for which this archaic term is still commonly used.
When we look into an animal's eye, we are looking into its aqueous compartment. Actually, to be more precise, we are looking into the anterior chamber of its aqueous compartment. The aqueous compartment is subdivided into two parts by the iris. The space in front of the iris is the anterior chamber, and the space behind the iris and in front of the lens is the posterior chamber. The anterior chamber is the only portion of the eye's interior that we can see clearly without special instruments.
Within the aqueous compartment, aqueous humor is constantly being produced and drained. It is produced in the posterior chamber by cells of the ciliary body. It then passes very slowly through the pupil into the anterior chamber, where it is drained by the canal of Schlemm and the fluid is returned to the bloodstream. The canal of Schlemm is a ringlike structure located way out at the periphery of the anterior chamber at the angle where the iris and the cornea meet (Figure 10-10).
The vitreous compartment of the eye is considerably larger than the aqueous compartment. It fills the whole back of the eyeball behind the lens and ciliary body. It contains vitreous humor (sometimes called the vitreous body), which is a clear fluid that has a soft, gelatinous consistency.
LENS
The lens of the eye is a soft, transparent structure made up of layers of microscopic fibers that are arranged like the
FIGURE 10-10 Formation of aqueous humor. Aqueous humor (large arrows) is formed in the posterior chamber by cells of the ciliary body. It slowly circulates through the pupil into the anterior chamber and is drained by the canal of Schlemm, located at the junction between the cornea and the iris. The pressure of the aqueous humor is indicated by small arrows.
The main role of the lens is to help focus a clear image on the retina regardless of whether the object being viewed is close up or far away. It does this with the help of the muscles of the ciliary body through a process called accommodation.
Accommodation is the process whereby the shape of the lens is changed to allow close-up and distant vision. When the muscle fibers of the ciliary body are relaxed, the suspensory ligaments that attach it to the periphery of the lens exert tension on the lens, pulling it into a flattened shape that allows clear distant vision (greater than about 20 feet). For close-up vision, the ciliary muscles must contract to take tension off the suspensory ligaments. This allows the lens to assume its natural, more rounded shape. So close-up vision requires muscle contractions in the ciliary body, but distant vision does not. This explains why we often suffer eyestrain when we do close-up work for long periods (like reading this book, for example). Fortunately, we can take advantage of our knowledge of how accommodation works to relieve the eyestrain; that is, we just need to look off into the distance periodically to give our ciliary muscles a break.
CLINICAL APPLICATION
CLINICAL APPLICATION
Glaucoma
Glaucoma is not one simple disease. Rather, it is a group of diseases characterized by increased intraocular pressure (pressure within the eye) that causes pain and can lead to blindness. In domestic animals, glaucoma is most often diagnosed in dogs, partly because it is seen fairly often in that species but partly because other species are not commonly tested for it. An instrument called a tonometer is used to test for glaucoma by measuring the intraocular pressure.
Glaucoma can have many origins, but the basic mechanism is that aqueous humor is being produced at a faster rate than it is being drained from the eye. This causes the intraocular pressure to rise. Most often the problem is due to insufficient drainage of aqueous humor, rather than overproduction. Therapy for glaucoma usually involves medical or surgical treatments designed to increase the rate at which aqueous humor drains from the anterior chamber.
The treatment of glaucoma usually is not as successful in domestic animals as it is in humans, because glaucoma is often in a very advanced stage when veterinary patients are presented for treatment. Unfortunately, glaucoma comes on very slowly and gradually with few, if any, clinical signs noticed early in the process, when treatment would be most beneficial. By the late stages, when the eyeball becomes noticeably enlarged and painful, it is often too late to save the animal’s vision. In many cases, the affected eye must be surgically removed.
layers of an onion. It is elastic and biconvex (meaning it bulges out on both sides). Its normal shape is fairly rounded, but it can be pulled into a flatter shape if tension is applied equally around its equator. The front (rostral) surface of the lens is in contact with aqueous humor, and its back (caudal) surface is in contact with vitreous humor.
Cataracts
A cataract is an abnormal condition of the eye whereby the lens becomes opaque. Instead of having a normal transparent appearance, a cataract lens appears milky. This impairs vision, particularly in dim light. Cataracts can be a normal part of the aging process. As a result, they are often seen in older animals, in which they can lead to total or near-total blindness. In younger animals, they can be genetically inherited or develop secondary to conditions such as infection, trauma, diabetes mellitus, or excessive exposure of the eyes to ionizing radiation (such as x-rays). Usually the only effective treatment for a cataract is surgical removal of the affected lens.
RETINA
The retina is the business end of the eye—the image sensor in the camera, so to speak. It is where the visual image is formed, sensed, and converted to nerve impulses that are decoded in the brain to re-form the image in the conscious mind. The whole reason the rest of the eye structures exist is to produce as accurate and clear an image as possible on the retina.
The retina is a complex, multilayered structure that lines most of the vitreous compartment of the eye. Its main components are three layers of neurons, the outermost (closest to the outside of the eyeball) of which is the actual layer of sensory cells. Figure 10-11 is a diagram of the major layers of the retina. From outside in, the layers of the retina are a thin pigment layer, the photoreceptor layer, the bipolar cell layer, the ganglion cell layer, and a nerve fiber layer that proceeds to the optic nerve. The bipolar cells and ganglion cells are neurons that integrate and relay nerve impulses from the photoreceptor
FIGURE 10-1 1 Cell layers of retina, with detail of rod and cone (photoreceptor) cells.
cells to the optic nerve. Because the photoreceptor cells form the outermost (deepest) layer of neurons, light rays (which approach the retina from the inside of the eye, having entered through the cornea and lens) must pass through the other, more superficial layers before reaching them. Fortunately, these layers are basically transparent to light.
The nerve fibers on the inside surface of the retina all converge at the optic disc. This is where they leave the eye to form the optic nerve that carries visual information to the brain. The optic disc contains only nerve fibers and a few blood vessels but no photoreceptor cells (so no visual images are formed there). It is the blind spot of the eye. See Figure 10-12.
The resolution of a digital photograph is determined by how many tiny dots (aka picture elements or pixels) make it up. In the digital camera of the eye, the photoreceptors form the pixels that make up the image that is perceived by the brain. Photoreceptor cells are neurons, but their dendrites have been modified into the actual sensory receptors for light. Two receptors, with different shapes and characteristics, are found among the photoreceptor cells: thin, rodshaped receptors and thicker, cone-shaped receptors. They are usually referred to as rods and cones and actually have different sensory roles (Table 10-2).
Rods are more sensitive to light than are the cones, but the rods produce a somewhat coarse image in shades of gray. The cones are more sensitive to color and detail than are the rods, but they do not function well in dim light. So rods are the main receptors for low-light vision, and cones perceive color and detail. (The next time you are in dimly lit
FIGURE 10-12 Normal fundus of canine eye. Note the ivory-colored optic disc, the blue tapetum, and the dark red retinal blood vessels. (From Dziezyc J: Color atlas of canine and feline ophthalmology, Philadelphia, 2005, WB Saunders Company.)
| TABLE 10-2 | Photoreceptor Characteristics | ||
| SENSITIVITY | SENSITIVITY | SENSITIVITY | |
| RECEPTOR | TO LIGHT | TO DETAIL | TO COLOR |
| Rods | High | Low | Absent |
| Cones | Low | High | High |
conditions, notice that you see little or no color and that things do not appear very sharp. Your rods are doing most of the work.)
Domestic animals are often said to be color blind. This is not really an accurate way to describe their vision, because color blindness implies a defect in color reception that sometimes occurs in humans. Most domestic animals can see colors to some extent, but because most have a lot of rods and not as many cones, colors probably appear washed out to them. Their color vision probably looks like an old color photograph that has been exposed to too much direct sunlight; the colors are there, but are they are pale and faded. Recent research has shown that different animal species may perceive certain colors more intensely than others.
Domestic animals also do not perceive detail as sharply as we do for another physical reason. Humans and other primates have a dense accumulation of cones in a small depression called the fovea centralis in the center of the retina. This is the area of clearest vision and the one you are using to read these words. Domestic animals do not have a fovea, so their vision is apparently less sharp. Focus your eyes on the center of this page. The way the top, bottom, and sides of the page appear to you is probably as sharp as the world appears to many animals. That might seem like a severe handicap to us sight-oriented humans, but that's not a good way to think of it. Remember that animals are equipped with sense organs that are appropriate for them. To other animals, our limited sense of smell would seem as much of a handicap as their visual limitations seem to us.
CLINICAL APPLICATION
The Blind Spot of the Eye
To detect the blind spot of your eye, mark an X on a piece of paper, and about 2 inches to the right of that mark an O. Now close or cover your left eye, focus your right eye on the X, and hold the paper about 12 inches from your face. Slowly move the page toward your eye. At some point, the O will disappear. When that happens, it has fallen on your optic disc, where there are no photoreceptors. So its image disappears! Interestingly, the brain normally fills in the blind spot area, so the conscious mind is not usually aware of that “hole” in the visual field.
FORMATION OF A VISUAL IMAGE
For the eye to transmit a clear visual image to the brain, a clear image must be formed on the retina. This is done by structures in the eye that refract (bend) light rays so they come into focus on the retina. Refraction is the bending of light rays that occurs as the rays pass into a medium of a different optical density, which affects the speed of light transmission. The more oblique the angle, the greater the degree of refraction. Because eye structures are curved, most light rays (except those directly in the center) strike the ocular surfaces at an angle.
Four refractive media in the eye help form a clear image on the retina: the cornea, the aqueous humor, the lens, and the vitreous humor. All contribute to the creation of a clear visual image, but the cornea does the majority of the refractive work. Its curved shape and the extreme difference between its optical density and that of the air in front of it result in significant refraction of light rays as they pass through it. The other refractive media, even the adjustable lens, only fine-tune the image that the cornea has formed. Interestingly, the image formed on the retina by these refractive structures is upside down. The brain somehow inverts the image so the conscious mind sees everything right side up.
TEST YOURSELF 10-6
1. Where is aqueous humor produced? Where is it drained from the aqueous compartment of the eye?
2. Which type of vision requires more muscular effort: close-up vision or far-away vision? Why?
3. Why is the optic disc the blind spot of the eye?
4. What kind of vision do the rods in the retina perceive? What do cones perceive?
5. What is the main refractive structure of the eye? Why?
EXTRAOCULAR STRUCTURES
Extraocular structures are not part of the eye itself, but they play important roles in its protection and functioning. They include the conjunctivae, the eyelids, the tear-production and drainage system, and the muscles that delicately move and position the eyeballs. Figure 10-13 shows many of the externally visible extraocular structures.
FIGURE 10-13 External view of a dog's left eye.
CONJUNCTIVA. The conjunctiva is a thin, transparent membrane that covers the front portion of the eyeball and lines the interior surfaces of the eyelids. The portion covering the front of the eyeball is called the bulbar conjunctiva (bulbar refers to the eyeball), and the portion lining the eyelids is called the palpebral conjunctiva (palpebral refers to the eyelids). The transparency of the conjunctiva allows the underlying tissues to show through, so it can be used as a window to see the blood vessels that are hidden elsewhere in the body by opaque structures, such as the skin. By looking through the conjunctiva at the lining of the eyelid, for example, we can often detect abnormalities such as anemia (paleness caused by decreased blood flow), jaundice (yellowish color), and cyanosis (dark purplish color).
The space between the bulbar and palpebral portions of the conjunctiva (between the eyelid and the eyeball) is called the conjunctival sac. It is normally only a potential space moistened by tears, but the ventral (bottom) conjunctival sac can be a useful place to deposit ophthalmic medications if the bottom lid is gently pulled out away from the eye before the medication is administered.
EYELIDS. The eyelids consist of upper and lower folds of skin that are lined by the thin, moist conjunctiva. The lateral and medial corners, where the eyelids come together, are called the lateral and medial canthi (singular, canthus). Along the margin of each eyelid are the tiny openings of the tarsal glands, also known as meibomian glands. These can be seen as a line of little dots along the eyelid margin. They produce a waxy substance that helps prevent tears
CLINICAL APPLICATION
Conjunctivitis
Conjunctivitis is inflammation of the conjunctiva of the eye. It is one of the most common eye diseases and occurs in all common domestic animal species. Signs include redness (hyperemia) and swelling (chemosis) of the conjunctiva, accompanied by ocular discharge and discomfort. The ocular discharge seen with conjunctivitis can range from watery (serous) to bloody (sanguineous) or pus-like (purulent). Causes include bacterial and viral infections, as well as physical and chemical irritants. Conjunctivitis is often seen in dogs that have been swimming in pools containing chlorinated water or hanging their heads out of car windows. Often the precise cause (etiologic factor) is not known. In mild cases of conjunctivitis, medical treatment may not be necessary. In more severe cases, antibiotic ointments or drops, which often contain corticosteroids to lessen the inflammation, are administered topically. Most cases of conjunctivitis respond well to treatment.
from overflowing onto the face. Eyelashes (cilia) are most prominent on the upper lid of most animals. Lower eyelashes are usually more sparse and thin, if they are present at all.
Domestic animals also have a third eyelid (also called the nictitating membrane) located medially between the eyelids and the eyeball. It consists mainly of a T-shaped plate of cartilage covered by conjunctiva. On its ocular surface (the surface in contact with the eyeball) are lymph nodules and an accessory lacrimal (tear-producing) gland. No muscles attach to the third eyelid. Its movements are entirely passive.
LACRIMAL APPARATUS. The lacrimal apparatus includes the structures that produce and secrete tears and the structures that drain them away from the surface of the eye. Tears are an important part of the overall liquid film that moistens and protects the surface of the eye. They are produced by the lacrimal glands and the accessory lacrimal glands of the third eyelids. The lacrimal glands are the primary source of tears. They are located dorsal and lateral to each eye inside the bony orbits that protect the eyeballs. Several small ducts from each gland deposit tears in the dorsal conjunctival sacs; from there, tears wash down over the surface of the eyes, aided by blinking movements of the eyelids. Figure 10-14 illustrates the position of the lacrimal glands.
The overall liquid film that moistens and protects the surface of the eye actually is made up of three main layers, only one of which comes from the lacrimal glands. These are an inner mucous layer, a middle tear layer, and an outer oily layer. The inner mucous layer comes from cells in the conjunctiva. It contains antibacterial substances that help protect the eye from infection. The middle tear layer comes from the lacrimal glands and the accessory lacrimal glands of the third eyelids. It serves to keep the cornea moist. The outer oily layer comes from the tarsal (meibomian) glands. It helps
FIGURE 10-14 Lacrimal apparatus. Tears produced by the lacrimal glands flow down over the surface of eye and drain into the lacrimal puncta. From there, they pass into the lacrimal sac and down the nasolacrimal duct to the nasal passage.
reduce evaporation of the underlying tear layer and prevents tears from flowing over the lid margin.
Tears are constantly being produced, so they must constantly be drained from the surface of the eye to prevent their spilling down the animal's face. The tear-drainage system is illustrated in Figure 10-14. Two small openings, one each in the upper and lower eyelid margins, drain the tears away from the surface of each eye. The openings are called the lacrimal puncta (singular, punctum), and they are located near the medial canthus of each eye. From the lacrimal puncta on each side, the tears flow down two small ducts to the lacrimal sacs and then down single ducts, called the nasolacrimal ducts, to the nasal cavity. This is why you get the sniffles when your eyes water: those excess tears have to go somewhere, and they drain down into your nasal cavity.
EYE MUSCLES. The extraocular eye muscles attach to the sclera of the eye (Figure 10-15). They are the small
FIGURE 10-15 Extraocular muscles of the right eye.
skeletal muscles that hold the eyeballs in place and delicately and accurately move them. They are capable of a wide range of movements that allow the eyes to track and examine objects with great precision. As you are reading this, note that your eyes track along the lines of words. If you concentrate, you can cause your eyes to move from letter to letter instead of from word to word. That's pretty delicate muscle control!
Humans share six extraocular muscles with domestic animals: four straight muscles and two oblique muscles. The four straight muscles are called the rectus muscles. (The word rectus means “straight.”) They include the dorsal, ventral, medial, and lateral rectus muscles. Their names indicate where they attach to the eyeball. The two oblique muscles are the dorsal and ventral oblique muscles. Many animals also have a seventh extraocular muscle—the retractor bulbi muscle. It retracts the eyeball slightly deeper into the orbit when it contracts. This may assist the activity of the other muscles by enhancing their mechanical advantage. Most movements of the eye involve a combination of several of these muscles contracting together.
TEST YOURSELF 10-7
1. How might examination of the bulbar and palpebral portions of the conjunctiva be useful as part of the overall physical examination of an animal?
2. An animal can intentionally blink its eyelids. Can it intentionally cover its eye with its third eyelid? Why or why not?
3. How would an animal with a plugged nasolacrimal system appear? Why?
4. If the medial rectus muscle of an animal's eye was damaged and lost its ability to contract, what would the effect be on the positioning of the affected eye? Why?