Organization of the brain
The cerebral hemispheres
The telencephalic vesicles form the telencephalon, which consists of two cerebral hemispheres. As the brain develops, the telencephalic vesicles grow posteriorly and laterally until they encase the diencephalon.
There is a proliferation of neurons resulting in the formation of three major white matter systems, including the cortical white matter, the corpus callosum, and the internal capsule. The cortical white matter includes neurons that run to and from the cerebral cortex. The corpus callosum includes neurons that connect the two cerebral hemispheres, and the internal capsule connects the cortex with the brain stem.The surface of the brain is marked by many convolutions. The grooves are called sulci (singular: sulcus); the ridges are called gyri (singular: gyrus). The outer six layers of neurons constitute the cerebral cortex. The convolutions greatly increase the surface area, thus increasing the amount of cortex. Whereas the brain of a chicken has a relatively flat surface, the brain of most domestic animals is considerably convoluted (Fig. 9.4).
The larger grooves that separate brain regions are called fissures. The longitudinal fissure separates the two
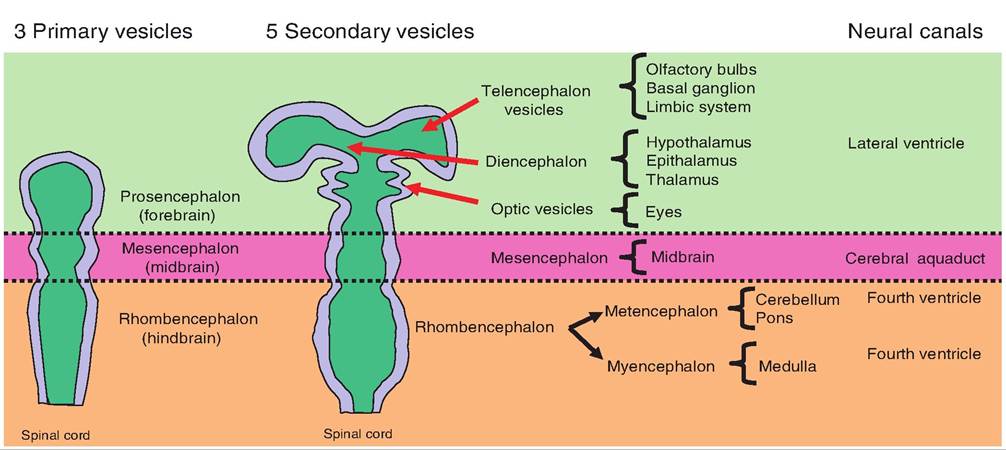
Fig. 9.3. Brain development. During embryonic development, the brain begins as three vesicles. The most rostral is the prosencephalon, which gives rise to the telencephalon and diencephalon vesicles. The most caudal, the rhombencephalon, gives rise to the metencephalon and myelencephalon.
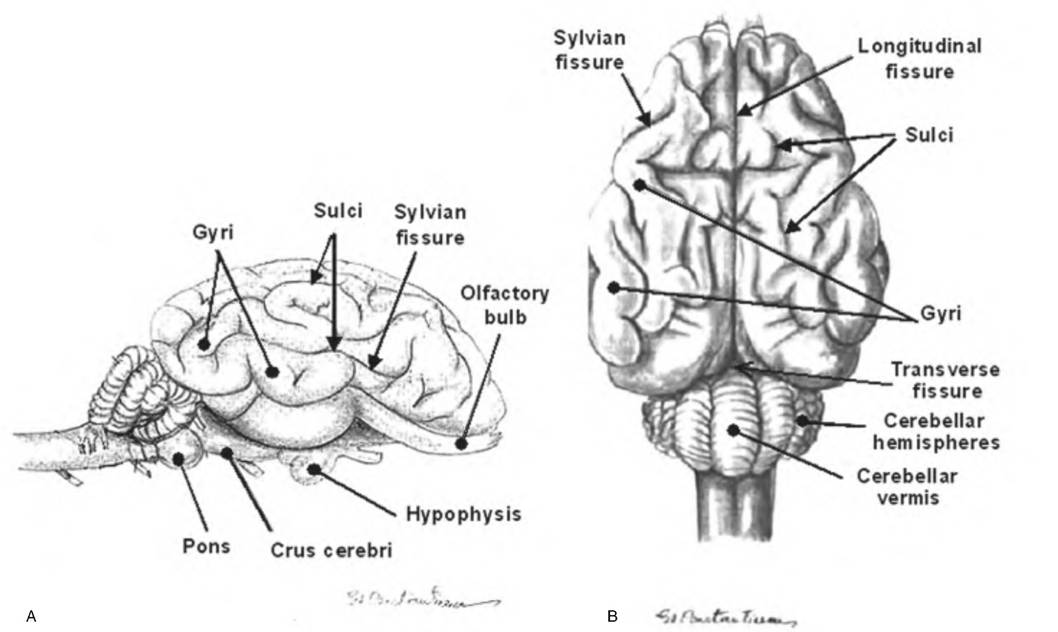
Fig. 9.4. Cytoarchitecture of the goat brain surface. (A) Lateral view; (B) dorsal view. Cephalization has resulted in the formation of ridges, called gyri, and valleys, called sulci.
Fissures are larger grooves separating major brain areas, such as the longitudinal fissure that separates the two cerebral hemispheres. (Reprinted from Constantinescu, 2001. Used by permission of the publisher.)cerebral hemispheres, whereas the cerebral hemispheres are separated from the cerebellum by the transverse fissure, which runs perpendicular to the longitudinal fissure.
When viewing an intact brain, four lobes—frontal, parietal, temporal, and occipital—are visible in the cerebral hemispheres. In most animals, the lobes are not delineated by sulci but are named for the cranial bones under which they lie (Fig. 9.5). In the human brain, various sulci separate the lobes, as will be described further. The frontal lobe lies just under the frontal bone making up the most rostral lobe of the cerebral hemispheres. Immediately caudal to the frontal lobe is the parietal lobe, which is separated from the frontal lobe by the central sulcus.
Two important gyri are also bordered by the central sulcus: the precentral gyrus anterior and the postcentral gyrus posterior to the central sulcus.
Below the parietal lobe is the temporal lobe, which is found under the temporal bone. The temporal lobe is separated from the frontal and parietal lobes by the lateral sulcus (lateral fissure, Sylvian fissure). Finally, at the back surface of the cerebral hemispheres is the
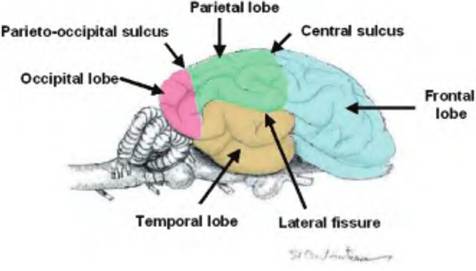
Fig. 9.5. Lobes of the brain. Four lobes are visible on the surface of the brain. The frontal lobe is most rostral. Caudad to the frontal lobe is the parietal lobe. Below the lateral fissure is the temporal lobe. The most caudal lobe is the occipital lobe. (Modified from Constantinescu, 2001.)
occipital lobe, which is separated from the parietal lobe by the parieto-occipital sulcus.
Although not visible on the surface, there is a fifth lobe called the insula, which is found by spreading the brain apart at the lateral fissure.
It is covered by parts of the temporal, parietal, and frontal lobes.Ventricles of the brain
As development progresses, the neurocoel expands to produce four chambers, called Cerebroventricles. Ependymal cells line the Cerebroventricles. Each cerebral hemisphere contains a lateral ventricle, also called the first and second ventricle. A thin layer of tissue called the septum pellucidum separates the two lateral ventricles from each other. The third ventricle is found in the diencephalon, and the fourth ventricle extends from the posterior surface of the pons and the anterior surface of the cerebellum to the superior portion of the medulla oblongata (Fig. 9.6). The fourth ventricle is continuous with the central canal of the spinal cord.
The lateral ventricles are connected to the third ventricle via the interventricular foramen of Monroe (Fig. 9.7). The third ventricle connects to the fourth ventricle via the mesencephalic aqueduct, also called the aqueduct of Sylvius or cerebral aqueduct. CSF flows by bulk flow from the lateral ventricles to the third ventricle to the fourth ventricle.
CSF can leave the fourth ventricle through the medial aperture called the foramen of Magendie and the two lateral apertures called the foramina of Luschka and enter the subarachnoid space (Fig. 9.8). This space is found between the arachnoid and pia mater, which, along with the dura mater, form the three meningeal layers covering the brain. The fluid in the subarachnoid space bathes the surface of the brain and spinal cord. Should the CSF not be able to flow through the ventricular system, it will back up in the ventricles causing hydrocephalus, or swelling of the ventricles. Since the skull cannot expand, increased pressure in the ventricles causes the soft tissue of the brain to be compressed, which leads to impaired brain function and death if untreated.
The adult human contains about 150 mF of CSF, and it is estimated that 430-450 mF are produced daily. Therefore, CSF is turned over every 6-7 hours.
After entering the subarachnoid space, CSF moves through one-way valves called arachnoid villi that project into the superior sagittal sinus formed by the dura mater located in the longitudinal fissure. The superior sagittal sinus is filled with venous blood. Therefore, once the CSF enters the superior sagittal sinus, it returns to the circulatory system.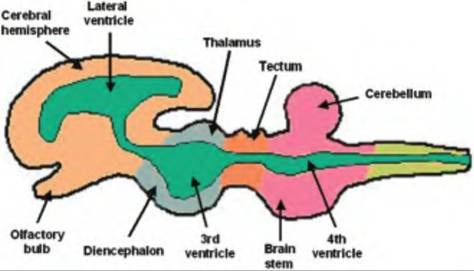
Fig. 9.6. Cerebroventricles. The neurocoele, or central cavity, in the neural tube eventually enlarges into four Cerebroventricles filled with cerebrospinal fluid. Two lateral ventricles, one in each cerebral hemisphere, empty into the medially located third ventricle located in the diencephalon. This empties into the fourth ventricle located between the cerebellum and the brain stem.
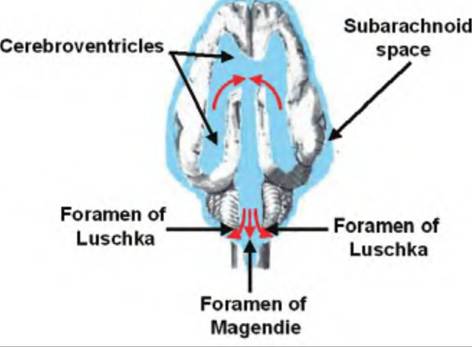
Fig. 9.8. Cerebrospinal fluid (CSF) movement out of ventricles. CSF travels from the fourth ventricle to the subarachnoid space via the medially located foramen of Magendie and the two lateral foramina of Luschka.
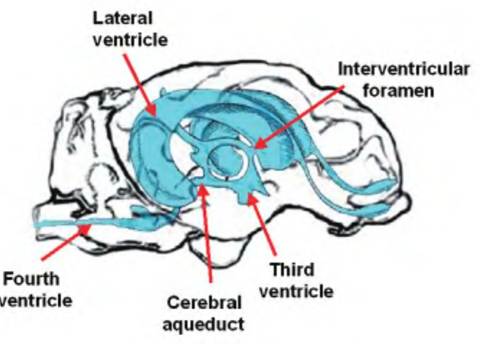
Fig. 9.7. Cerebroventricles of the sheep. Cerebrospinal fluid (CSF) is formed in the choroid plexus located in each lateral ventricle.
Via bulk flow, the CSF flows from the lateral ventricles through the interventricular foramen of Monroe to the third ventricle. It then flows through the cerebral aqueduct to the fourth ventricle.
Cerebral cortex
The cerebral cortex is arranged as layers of cells that lie parallel to the surface of the brain. The layer closest to the surface of the brain is separated from the pia mater by a zone called the molecular layer or layer I that lacks neurons. Furthermore, at least one layer contains pyramidal cells that project large dendrites, called apical dendrites, that project to layer I where they form multiple synapses.
The neocortex, which is found only in mammals and is associated with higher brain functions such as conscious behavior, is found over most of the surface of the cerebral hemisphere and consists of six layers of cells (Fig. 9.9). Medial to the lateral ventricles is an area of cortex named for its unique shape called the hippocampus (Greek for "seahorse"). It is only a single cell layer. Ventral and lateral to the hippocampus is the third area of cortex called the olfactory cortex (piriform, or pyriform cortex) that consists of two cell layers. The olfactory cortex connects with the olfactory bulbs. The olfactory cortex is separated from the neocortex by the rhinal fissure.
In humans, the neocortex was extensively mapped by Korbinian Brodmann in 1906 in which he numbered 52 different cortical areas each having a common Cytoarchitecture. Such an extensive mapping has not been done in other animals. The cortex can be designated into three areas: (1) motor areas responsible for the control of voluntary motor functions, (2) sensory areas responsible for perception of various sensations, and (3) association areas that integrate the motor and sensory signals.
The motor and sensory areas can be grouped together into projection areas, thus allowing the cortex to be subdivided into three components including the projection areas, rhinencephalon (olfactory and limbic) areas, and association areas. The projection and association areas comprise the neocortex. Association areas receive sensory information, process that information, develop a response, and predict its consequences. As animals became more evolutionarily advanced, the association areas became more developed (Fig. 9.10).
The white matter, or fibers, in the cerebral cortex form three types of fibers:
1. Association fibers. Association areas allow complex problem solving and creative thinking. There are association areas found in the frontal, temporal, parietal, and occipital lobes. These fibers course within a cerebral hemisphere, thus connecting various areas of the cortex.
The parietal and occipital lobe association areas are involved with cognitive functions, whereas the frontal association areas are involved with general alertness, intelligence, and temperament. The temporal association area is involved with learning and memory. Although these areas are not well developed in cats and dogs, lesions to the frontal association area result in changes in behavior and personality.2. Projection fibers. Neurons that leave the cerebrum and enter the brain stem via the internal capsule are called projection fibers. They therefore connect the cortex with subcortical structures, as well as the remainder of the nervous system.
3. Commissural fibers. Each cerebral hemisphere generally controls the contralateral side of the body; therefore, fibers need to cross between hemispheres. When crossing from one side of the cerebral cortex to the other, they do so as commissural fibers.
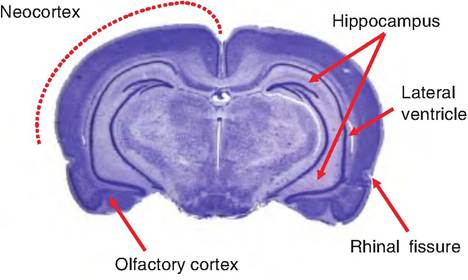
Fig. 9.9. Three types of mammalian cortex. The neocortex is found on the outer surface of the cerebral hemispheres. Medial to the lateral ventricles is a second type of cortex called the hippocampus, part of the limbic system. The third type of cortical tissue is the olfactory cortex, located ventrally and laterally to the hippocampus.
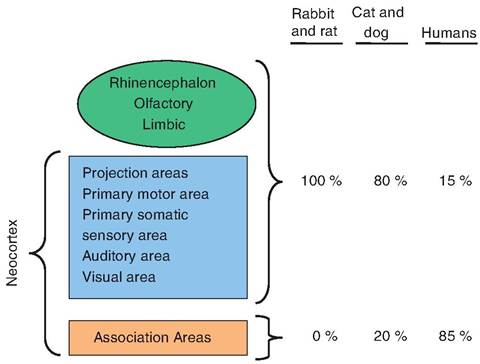
Fig. 9.10. Evolutionary changes in the Cerebrocortex. Through evolutionary development, the ability to display higher-order thinking is associated with enhanced development of the association areas. (Modified from King, 1987.)
Motor areas
These are cortical areas responsible for motor functions. The primary motor cortex is the final site for cortical processing of motor commands before messages are then sent to the somatic muscles. While in humans the primary motor area lies in the posterior part of the frontal lobes just anterior to the central sulcus, in mammals, this area lays in the rostral region of the frontal lobes (Fig. 9.11). Unlike birds, reptiles, amphibians, and fish, mammals possess a pyramidal system. The pyramidal system consists of corticospinal fibers that travel from the primary motor area through the medullary pyramids located at the base of the medulla oblongata, to the somatic motor neurons found in the spinal cord. It also includes the corticonuclear fibers that project to the nuclei of those cranial nerves that innervate striated muscles in the head. The pyramidal fibers decussate, or cross over, in the pyramids of the medulla.
The extrapyramidal system includes all the descending somatic motor pathways excluding those described earlier that constitute the pyramidal system. The extrapyramidal system is phylogenetically old and found in all but the lowest vertebrates. It consists of nine main motor areas located in the forebrain, midbrain, and hindbrain (Fig. 9.12). These will be discussed in depth later in the chapter.
Sensory areas
Unlike the motor areas that are located in the frontal lobe, sensory areas are located throughout the cortex. The primary somatosensory cortex receives information from sensory receptors located in the skin and proprioceptors in skeletal muscles.
Cerebral white matter
Basal ganglion
In addition to the cerebral cortex discussed earlier, and its commissures, and association and projection fibers, there are some deep subcortical nuclei called the basal ganglia or nuclei (Fig. 9.13). Although the definition of the structures included in the basal ganglion varies, it generally includes the caudate nucleus, putamen, globus pallidus, substantia nigra (consisting of the pars reticulata and pars compacts), and subthalamic nucleus. The putamen and globus pallidus (or pallidum) together form the Ientiform nucleus that laterally borders the internal capsule. The Ientiform nucleus and caudate are collectively called the corpus striatum because the fibers of the internal capsule, a collection of fibers that runs between the neocortex and thalamus, pass through them, giving them a striated appearance.
The corpus striatum receives most of the inputs to the basal ganglion from the cerebral cortex, thalamus, and brain stem. The corpus striatum sends projections to the globus pallidus and substantia nigra that provide the major output projections from the basal ganglion. These projection fibers travel through the thalamus to the premotor and prefrontal cortex, and therefore affect motor movement.
Limbic system
The limbic system consists of a group of structures located in the medial region of each cerebral hemisphere
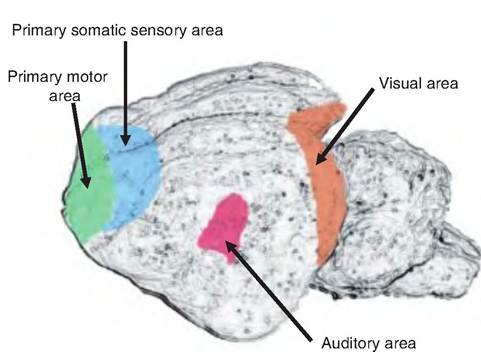
Fig. 9.11. Primary projection areas of the cat. The primary motor and somatic sensory areas spread over the medial surface of the frontal lobes. The visual area is located in the occipital lobe; the auditory area is located in the temporal lobe.
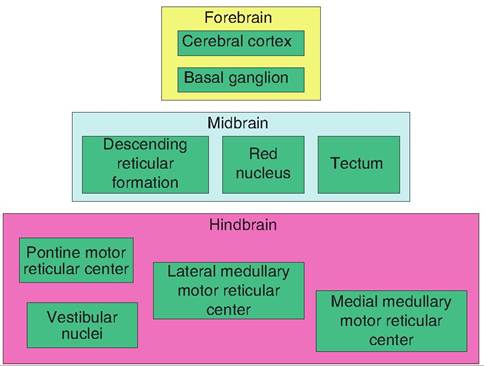
Fig. 9.12. Extrapyramidal system. The extrapyramidal system is a multisynaptic motor pathway that includes all those motor neurons not part of the pyramidal system. It consists of nine motor centers scattered throughout the brain.
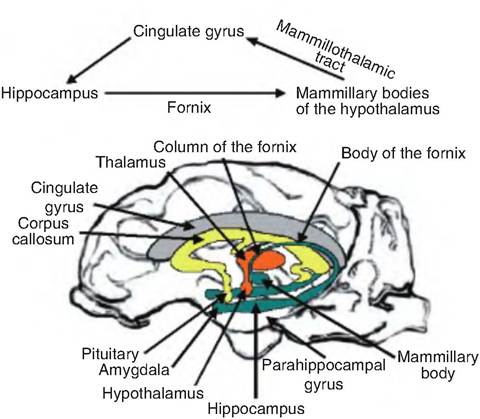
Fig. 9.14. Limbic system. The limbic lobe includes the cingulate gyrus, parahippocampal gyrus, and the hippocampal formation that lies deep in the parahippocampal gyrus. The hippocampal formation includes the hippocampus, dentate gyrus (deep to the parahippocampus gyrus), and subiculum. The limbic system additionally includes parts of the rhinencephalon, amygdala, hypothalamus, and anterior nucleus of the thalamus. The fornix helps connect parts of the limbic system.
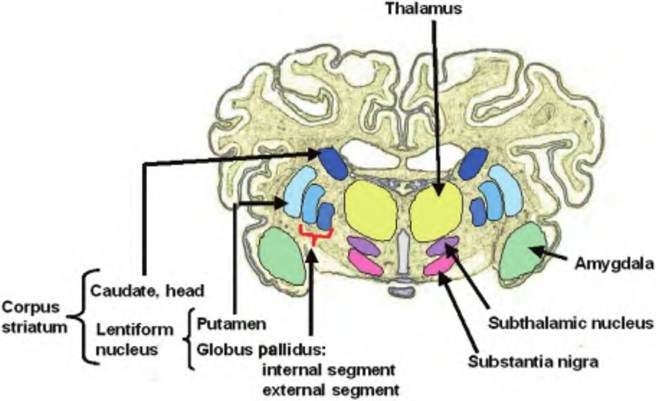
Fig. 9.13. Basal ganglion. The basal ganglion, or nucleus, is a collection of brain nuclei that function in motor movement as part of the extrapyramiclal system. The basal ganglion includes the globus pallidus and putamen, which together constitute the Ientiform nucleus. Also included is the caudate, which, combined with the Ientiform nucleus, forms the corpus striatum. Finally, also included in the basal ganglion are the amygdala, subthalamic nucleus, and substantia nigra.
(Fig. 9.14). These structures encircle (limbus = ring or border) the brain stem. The limbic lobe of the cerebral hemisphere includes gyri surrounding the diencephalon, as well as other underlying structures. Specifically, it consists of three gyri. The cingulate gyrus is dorsal to the corpus callosum. The dentate gyrus and parahippocampal gyrus form the inferior and posterior portions of the limbic lobe. These later two gyri conceal the hippocampus, which is a nucleus lying inferior to the lateral ventricle. The fornix is a fiber tract running inferior to the corpus callosum and connecting the hippocampus with the hypothalamus where it ends in the mammillary bodies. Also included in the limbic system is the anterior nucleus of the thalamus, which relays information from the mammillary bodies to the cingulate gyrus. The amygdaloid body (nucleus) serves as the interface between the limbic system, cerebrum, and various sensory systems.
The limbic system is involved in emotional and behavioral patterns. The functions of the limbic system include (1) establishment of emotional states; (2) linking of conscious functions with unconscious, autonomic functions; and (3) long-term memory storage and retrieval. The rabies virus generally attacks the hippocampus and results in emotional changes including bouts of terror and rage. The amygdala is believed to be the major component of the limbic system involved in emotion since electrical stimulation of this region produces feelings of fear and apprehension, whereas damage to this region causes tameness. Removal of the amygdala will allow a cat to wander through a colony of monkeys ignoring the monkey's hoots and threats.
It is believed that the hippocampal formation processes information from the Cingulated gyrus. This information is sent to the mammillary bodies of the hypothalamus via the fornix (Fig. 9.14). The hypothalamus gives feedback to the cingulate gyrus by a pathway from the mammillary bodies to the anterior thalamus via the Inammillofhalamic tract, and then to the cingulate gyrus.
Diencephalon
The telencephalon and diencephalon make up the forebrain and are derived from the rostral-most vesicle called the prosencephalon. Surrounded by the cerebral hemispheres, the diencephalons consist of three paired structures—the thalamus, hypothalamus, and epithalamus.
Thalamus
The thalamus lies dorsal to the hypothalamus and is bordered by the caudate nucleus dorsally and the internal capsule laterally. Its two halves are separated by the third ventricle. There is a collection of nuclei found in each half of the thalamus (Fig. 9.15). The intermediate mass of the thalamus extends through the third ventricle connecting the two halves.
The thalamus is the major relay station for sensory information generated in the periphery and transferred for processing to the cerebral hemispheres. It also integrates motor information from the cerebellum and basal ganglion and transfers such information to the motor regions of the cortex.
The nuclei of the thalamus are generally classified into four groups—anterior, ventrolateral, medial, and posterior. The ventrolateral and posterior groups are located lateral to the medullary lamina, a fiber tract that runs the rostrocaudal length of the thalamus. The medial group of nuclei is located medial to the medullary lamina. The medullary lamina splits at the rostral end of the thalamus and encases the anterior group.
The posterior nuclear group consists of the pulvinar and medial and lateral geniculate bodies, whereas the anterior group consists only of the anterior nucleus. The anterior nucleus receives input from the mammillary nuclei of the hypothalamus and is thought to participate in memory and emotion. It is also interconnected with the Cingulated and frontal cortices and is therefore part of the limbic system.
The medial group, consisting mostly of the medial nuclei, receives inputs from the basal ganglia, amygdala, and midbrain. It projects to the frontal cortex and has been implicated in memory. The ventrolateral group consists of the remaining nuclei shown in Figure 9.15. The ventral anterior and ventral lateral nuclei carry information from the basal ganglia and cerebellum to the motor cortex. The ventral posterior lateral nucleus carries somatosensory information to the neocortex.
Hypothalamus
The hypothalamus (hypo = below) is found ventral to the thalamus and forms the inferolateral walls of the third ventricle (Fig. 9.16). It extends from the optic chiasm, the site where the optic nerves cross to the contralateral sides, to the posterior border of the mammillary bodies. The infundibulum, which connects the hypothalamus to the pituitary, lies between the optic chiasm and mammillary bodies. It arises from the tuber cinereum, which is an oval protuberance located on the floor of the third ventricle.
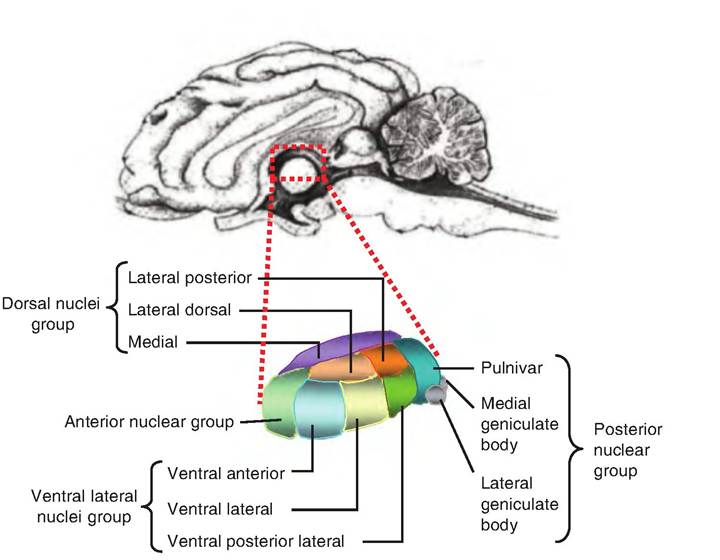
Fig. 9.15. The thalamus. The thalamus consists of four groups of nuclei. The anterior nucleus and pulvinar constitute the anterior and posterior groups, respectively. The medial group consists mainly of the mediodorsal nuclei (not shown).
The hypothalamus serves an essential role in the control of homeostasis and reproduction. Once termed the "head ganglion" of the autonomic nervous system, the hypothalamus was thought to control the autonomic nervous system. This was due to the observations that electrical stimulation of the hypothalamus altered autonomic functions. However, it is now believed that the hypothalamus plays more of an integrative role in regulating the autonomic nervous
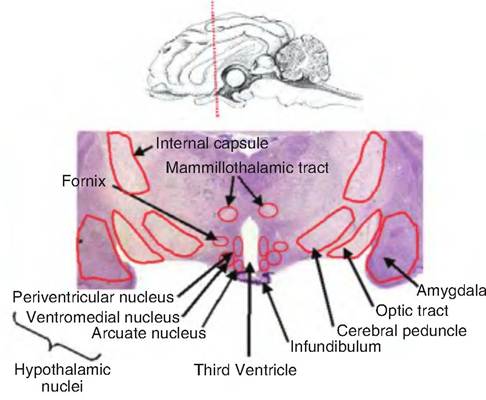
Fig. 9.16. Diencephalon. A coronal section through the sheep brain at the level of the diencephalon is shown. The hypothalamus is that region surrounding the third ventricle.
system, and that many of the early effects attributed to the hypothalamus were a result of stimulation of fiber pathways that course through the area.
While relatively small, the hypothalamus is a complex structure containing many nuclei (Fig. 9.17). It can be divided into three general areas: anterior, middle, and posterior. The anterior region lies dorsal to the optic chiasm and is called the preoptic region. It contains the Suprachiasmatic nucleus, which is involved in circadian rhythms, the anterior hypothalamic nucleus, and the lateral and medial preoptic nuclei. This area also controls blood pressure, body temperature, and reproductive activity.
The middle region lies dorsal to the pituitary and contains various nuclei including the dorsomedial, ventromedial, paraventricular, supraoptic, and arcuate nuclei. The paraventricular nucleus contains magno- cellular and parvocellular neuroendocrine regions. The arcuate nucleus and parvocellular region of the paraventricular nucleus produce hormones that control the anterior pituitary. The magnocellular area of the paraventricular nucleus, consisting of large neurons, produces oxytocin that is secreted in the neurohypophysis. The paraventricular nucleus also has connections to both parasympathetic and sympathetic preganglionic neurons in the medulla and spinal cord, and can thus control the autonomic nervous system. The supraoptic nucleus produces antidiuretic hormone, which is also secreted from the neurohypophysis.
The posterior region of the hypothalamus contains the mammillary bodies, Iuberomammillary nucleus, and the overlying posterior hypothalamic region. This latter nucleus contains histamine and appears involved in wakefulness and arousal.
The hypothalamus can also be divided longitudinally into the periventricular, medial, and lateral zones that each run the rostral-caudal length of the hypothalamus. The periventricular zone lies immediately adjacent to the third ventricle. There are two major fiber systems running through the hypothalamus. The medial forebrain bundle runs through the lateral hypothalamus connecting the hypothalamus with the brain stem, basal forebrain, amygdala, and cerebral cortex. The second fiber system is located medial to the major hypothalamic nuclei along the wall of the third ventricle, and it links the periaqueductal gray matter in the midbrain with the hypothalamus. This latter fiber system is involved in stereotyped behavioral patterns such as posturing during sexual behavior.
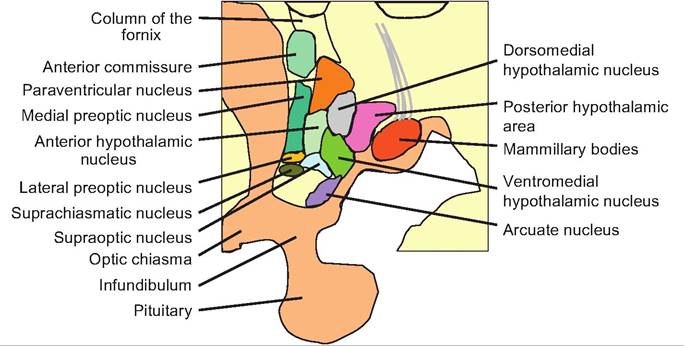
Fig. 9.17. Hypothalamus. Saggital view of the hypothalamus showing the various hypothalamic nuclei.
The hypothalamus integrates the regulation of the autonomic nervous system and thereby controls growth, feeding, drinking, circadian rhythms, and maternal behavior. Specifically, the hypothalamus plays a key role in six physiological areas:
1. It controls blood pressure and electrolyte composition by affecting water consumption and salt appetite as well as controlling blood osmolarity and vasomotor tone. When the blood becomes hyperosmotic, osmoreceptors in the hypothalamus are stimulated and result in the release of antidiuretic hormone (vasopressin) from the posterior pituitary that causes the collecting tubules in the kidneys to reabsorb water. This results in a decrease in blood osmolarity.
2. It regulates body temperature by controlling metabolic rate, vasomotor tone, and thermoregulatory behavior.
3. It regulates food and water intake. For example, an increase in white adipose levels in the body causes increased plasma leptin levels. Leptin circulates to the hypothalamus where it decreases the production and release of neuropeptide Y by neurons in the arcuate nucleus, thus resulting in decreased food intake.
4. It regulates reproduction through the hormonal control of estrus, pregnancy, and lactation. Releasing factors produced in the hypothalamus and released into the hypothalamic-hypophyseal portal system control the release of hormones from the anterior pituitary. Conversely, the hormones released from the posterior pituitary (i.e., oxytocin and antidiuretic hormone) are produced in supraoptic and paraventricular nuclei of the hypothalamus and then travel down the axons and are released from the posterior pituitary.
5. It controls the stress response. This includes the physiological and immunological responses such as increased blood pressure and decreased humoral immunity.
6. It controls circadian rhythms and sleep-wake cycles. The Suprachiasmatic nucleus acts as a major internal clock in mammals. This nucleus is entrained by light via the Tetinohypothalamic tract that runs from the retina to the Suprachiasmatic nucleus. Lesioning the Suprachiasmatic nucleus attenuates sleep and other circadian rhythms. These rhythms can be restored by transplanting a fetal suprachi- asmatic nucleus. Furthermore, stimulation of the posterior hypothalamus produces arousal that is mediated by histaminergic neurons, whereas destruction of this area decreases histaminergic output, which causes sleep, much like that caused by antihistamines. In contrast, stimulation of the anterior hypothalamus induces sleep. This appears to be mediated by GABAnergic inhibitory neurons.
Epithalamus
The epithalamus lies superiorly, caudally, and medially relative to the other parts of the diencephalon and thus represents a cephalad extension of the pretectum (just rostral of where the midbrain fuses with the thalamus) of the mesencephalon. The epithalamus includes the pineal body (epiphysis), the habenula, the habenular commissure, and the striae medullares.
The pineal body serves a role in sensing light and thereby controlling the sleep-wake cycle via the production of melatonin. It also has an important role in seasonal breeding animals such as sheep in which a decrease in day length, and therefore a decrease in melatonin levels, results in estrus.
Mesencephalon
The mesencephalon lies between the diencephalon and pons (Fig. 9.18). The tectum (Latin for "roof") forms the roof of the mesencephalon and contains two pairs of prominent bulges known as the corpora quadrigemina. Consisting of the superior and inferior colliculi, these nuclei process visual and auditory stimuli, respectively. The tegmentum forms the floor of the midbrain.
The superior colliculus receives visual input from the lateral geniculate nucleus of the thalamus from the ipsilateral side. The inferior colliculus receives auditory information from the medulla oblongata and pons, and relays that information to the medial geniculate on the same side. The inferior colliculi control the orientation of movements in response to auditory stimuli. The superior colliculi control the orientation of movements of the eyes, head, and neck in response to visual stimulation such as bright light.
The cerebral aqueduct connecting the third ventricle with the fourth ventricle runs through the mesencephalon. The tegmentum forms the part of the midbrain lying ventral to the cerebral aqueduct. On the walls and floor of the mesencephalon are found the red
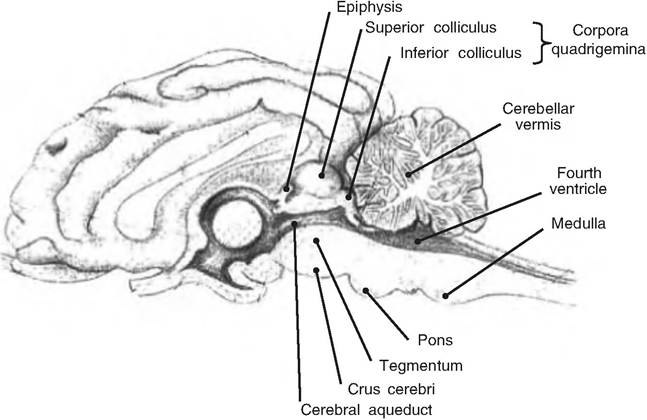
Fig. 9.18. Mesencephalon. Saggital view of goat brain showing the mesencephalon. (Modified from Constantinescu, 2001.)
nuclei, substantia nigra, reticular formation, and nuclei for the third (occulomotor) and fourth (trochlear) cranial nerves. The red nucleus is named for its red color caused by the extensive blood supply it receives and the iron pigment found in the cell bodies. It is the largest nucleus of the reticular formation discussed further. It receives information from the cerebrum and cerebellum, and is involved in subconscious motor control of the forelimb positions and background muscle tone. The substantia nigra (nigra = black), named for its black pigment, lies lateral of the red nucleus.
Visible on the sides of the mesencephalon are the cerebral peduncles (little feet). They contain descending fibers connecting to the cerebellum via the pons, and descending fibers carrying voluntary motor signals from the cerebral hemispheres. The large ventral bundle of efferent fibers in the cerebral peduncles are referred to as the crus cerebri.
Metencephalon
The metencephalon consists of the pons and cerebellum. The pons links the spinal cord with the forebrain, as well as the cerebellum with the forebrain and spinal cord. It forms part of the anterior surface of the fourth ventricle.
The pons has four major components:
1. Cranial nerves. Cranial nerves V (trigeminal), VI (abducens), VII (facial), and VIII (vestibulocochlear) are found in the pons.
2. Nuclei controlling respiration. Located bilaterally are two respiratory centers called the apneustic center and the pneumotaxic center.
3. Fiber tracts and nuclei connecting the cerebellum to the brain stem, forebrain, and spinal cord.
4. Transverse tracts. Found on the anterior surface of the pons are fiber tracts that link nuclei found in the pons with the contralateral (opposite) cerebral hemisphere.
The cerebellum (Latin for "little brain") is the second largest region of the brain, accounting for 10% of its total mass, but containing half of all neurons in the brain. It is located dorsal to the pons and medulla oblongata, with the fourth ventricle in between, and is separated from the cerebral hemispheres by the transverse fissure.
The cerebellum is important in coordinating muscle movement and maintaining balance. For example, when a horse jumps over an obstacle, it requires the coordinated movement of all the limbs, as well as a good sense of where the horse is in space. The cerebellum compares what movement was intended to what movement happened. The cerebellum can then institute adjustments to the intended movement while they are in progress, or during repetitions.
Resembling a piece of cauliflower, the cerebellum is connected to the brain stem by the inferior, middle, and superior cerebellar peduncles, also called the res- tiform body, brachium pontis, and brachium conjucti- vum, respectively (Fig. 9.19). The surface of the cerebellum consists of small convolutions similar to small gyri, called folia (leaves), running side to side. The anterior and posterior lobes of the cerebellum are separated by the primary fissure. The anterior and posterior lobes are further subdivided into two and five lobules, respectively. The two lateral cerebellar hemispheres are separated by a narrow ridge of cortex
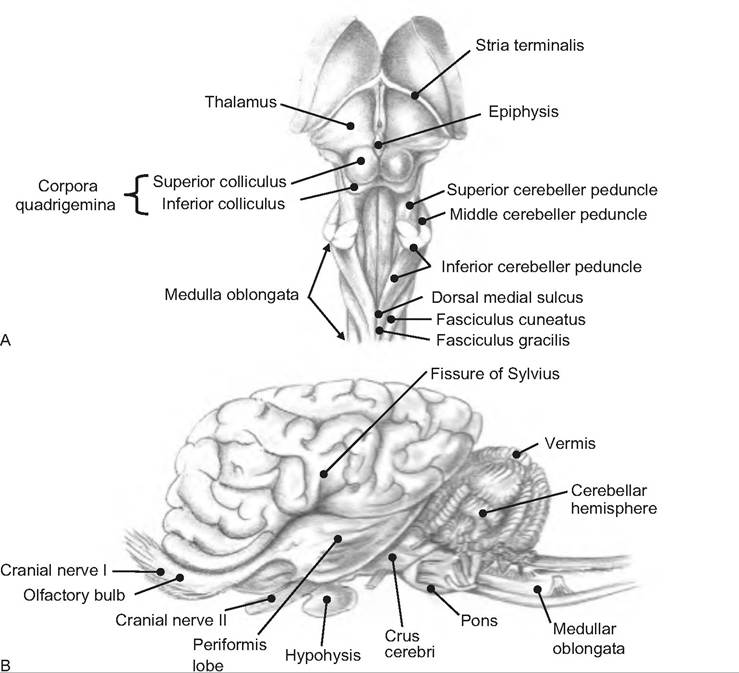
Fig. 9.19. B rain of the horse. (A) Dorsal view of brain stem. (B) Lateral view of brain. (Reprinted from Constantinescu and Constantinescu, 2004. Used by permission of the publisher.)
called the vermis (Latin for "worm"). When viewing a saggital cut of the cerebellum, the flocculonodular lobe is seen between the cerebellar hemispheres, vermis, and the roof of the fourth ventricle. This lobe is separated from the body of the cerebellum by the posterolateral fissure (Fig. 9.19).
The surface of the cerebellum is covered with a cortex containing large, highly branched neurons called Purkinje cells. These cells contain an extensive dendritic tree and receive input from up to 200,000 synapses. The importance of this input is evidenced by the fact that 40 times as many axons project to the cerebellum as leave it. Inside the cerebellum, forming the shape of a tree, is white matter called the arbor vitae, or "tree of life." In addition, there are three pairs of deep cerebellar nuclei.
The cerebellum monitors all proprioceptive, visual, tactile, balance, and auditory sensory information. Most of this information is carried to the Purkinje cells, thus bypassing deep cerebellar nuclei. The motor information traveling to the cerebellum from the pyramidal and extrapyramidal system is relayed from nuclei in the pons or cerebellar nuclei found in the arbor vitae.
Information leaves the pons via the cerebellar peduncles. The superior cerebellar peduncle carries signals to the midbrain, diencephalons, and cerebrum while the middle cerebellar peduncle transmits signals to the sensory and motor nuclei in the pons. The inferior cerebellar peduncle relays information between the cerebellum and nuclei in the medulla oblongata and cerebellar tracts from the spinal cord.
Signals generated in the sensorimotor areas of the cortex project to nuclei in the pons, and then to the cerebellum. This is a large pathway containing approximately two times the axons found in the pyramidal tract. The lateral cerebellum then sends signals back to the motor cortex via the ventral lateral nucleus of the thalamus.
Medulla oblongata
Originating from the myelencephalon, the medulla oblongata is continuous with the spinal cord. The rostral portion of the medulla contains part of the fourth ventricle; the caudad portion contains a central canal.
The medulla contains three major groups of nuclei. The first is part of the reticular formation, which extends from the medulla to the mesencephalon, and was described earlier. The portion of the reticular formation in the medulla is responsible for regulating autonomic functions. The two major reflex centers are the cardiovascular centers and the respiratory rhyth- micity center. The cardiovascular centers regulate heart rate, the strength of cardiac contractions, and the flow of blood through peripheral tissues. The cardiovascular centers include the cardiac center and vasomotor center. The respiratory rhythms center regulates the pace of respiratory movements. These are controlled by inputs from the apneustic and pneumotaxic centers in the pons.
The second group of nuclei is for the cranial nerves (see Fig. 9.22, later in the chapter). Five cranial nerves originate in the medulla including the VIII (vestibulocochlear), IX (glossopharyngeal), X (vagus), XI (spinal accessory), and XII (hypoglossal) nerves. Note that cranial nerve VIII carries sensory information from the inner ear to both the vestibular and cochlear nuclei that extend from the pons into the medulla.
The third group of nuclei acts as relay stations from sensory and motor pathways. They include the nucleus gracilis and nucleus cuneatus that carry somatic sensory information to the thalamus. The solitary nucleus, located bilaterally, receives visceral sensory information coming from the spinal cord and cranial nerves. This information is relayed to the autonomic centers in the medulla and elsewhere. The olivary nuclei, which form olive-shaped bulges along the ventrolateral surface of the medulla, transmit somatic motor commands from higher brain centers to the cerebellar cortex.
Functional systems
The reticular activating system (RAS)
The reticular formation forms the core of the brain stem tegmentum extending from the medulla oblongata through the pons and midbrain. It consists of loosely clustered neurons and is homologous to the central gray area of the spinal cord that contains interneurons. It is termed an area since it lacks cell clusterforming nuclei, and is spanned by long bundles of ascending and descending fibers, giving it a reticulated appearance.
The area can be divided into lateral and medial regions. The lateral region generally consists of parvo- Cellular interneurons found close to motor nuclei of the cranial nerves, and is involved in coordinated reflexes and simple stereotyped behaviors. These are generally related to the vagus nerve and include gastrointestinal responses such as swallowing and vomiting; respiratory activities such as initiation of respiratory rhythm, coughing, hiccupping, and sneezing; and cardiovascular functions including the baroreceptor reflexes. The main descending lateral pathway is the rubrospinal tract originating in the magnocel- Iular portion of the red nucleus and traveling to the lateral column of the spinal cord. The lateral reticulospinal pathway inhibits antigravitational muscles and can lead to atonia.
Neurons in the medial region are larger (magno- cellular) and have long ascending and descending axons (e.g., pontine reticulospinal tract) that modulate neurons involved in movement and posture, pain, autonomic functions, and arousal. In particular, this pathway enhances antigravitational reflexes by facilitating extensors and helping standing posture.
Within the reticular formation, certain neurons send continuous impulses via the thalamus to the cerebral cortex. These impulses cause excitation, thus causing arousal. This system is called the ascending RAS. This system receives input from ascending sensory tracts. It is responsible for filtering out selected sensory signals after they become familiar and weak.
The RAS is inhibited during sleep and is thus involved in the sleep-wake cycle. Studies in the 1940s and 1950s revealed that electrical stimulation of the midbrain reticular formation (i.e., midline tegmental area) promoted wakefulness, whereas damage to this region caused a state similar to that in non-REM sleep. This area was found to be inhibited by a system in the medulla. Making a cut just caudad to the midbrain (i.e., midpontine-pretrigeminal transection) prevented an animal from sleeping.
Stimulating the RAS increases the firing rate of various neurons including the locus coeruleus in the pons, which contains norepinephrine, the raphe nuclei, which produces serotonin, acetylchonine-containing neurons in the brain stem and forebrain, as well as histamine-containing neurons in the midbrain. These neurons synapse in the thalamus, cerebral cortex, and other brain regions.
Sleep
Sleep can be defined as a readily reversible state of reduced responsiveness to and interaction with the environment. Therefore, behaviorally, sleep has four characteristics: (1) reduced motor activity, (2) decreased responsiveness, (3) stereotypic postures, and (4) ready reversibility. Coma and anesthesia do not qualify as sleep since they are not readily reversible.
Sleep is categorized into two categories. Rapid eye movement (REM) sleep, is noted for the movement of the eyes under the eyelids, and the almost complete inhibition of skeletal muscle tone. This stage of sleep
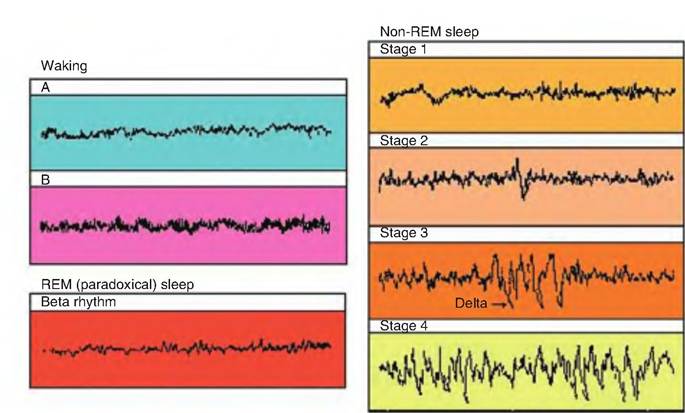
Fig. 9.20. EEG. Various EEG wave patterns seen during wakefulness and various stages of sleep. (A) Beta rhythm (alert and awake). (B) Alpha rhythm (relaxed, eyes closed).
is when most dreaming occurs. Non-REM sleep is characterized by a change in the electroencephalogram (EEG). Non-REM sleep is sometimes called slow-wave sleep because of the large, slow EEG rhythms (Fig. 9.20). All mammals sleep, although the length of sleep varies. Birds also show both REM and non-REM sleep, although their sleep episodes are short, with REM sleep lasting maybe only a few seconds. In addition, muscle atonia during REM sleep is rare in birds.
Non-REM sleep appears designed for rest. Neuronal activity is low, muscle tension is reduced, and movement is minimal, although movement is possible. Body temperature and metabolic rate are at their lowest. Sympathetic outflow decreases while the activity of the parasympathetic nervous system increases. Therefore, heart rate, blood pressure, respiration rate, and kidney function decrease, and digestive function increases. The slow, large-amplitude EEG rhythms indicate that the cortex is oscillating in high synchrony while receiving little, if any, sensory input.
In contrast, the EEG during REM sleep is nearly indistinguishable from that of the awake brain. It displays fast, low-voltage fluctuations and is therefore sometimes called paradoxical sleep. Oxygen consumption of the brain is higher during REM sleep than when an animal is awake and concentrating. While most of the muscles of the body are in atonia, the muscles controlling eye movement and those in the inner ear are active.
Roughly 75% of the sleep cycle is spent in non-REM sleep and 25% in REM sleep. An animal cycles between these two stages of sleep going from non-REM to REM sleep, and back approximately every 90 minutes. This is an example of an ultradian rhythm that is shorter than circadian rhythm.
Non-REM sleep is divided into four stages. The lightest stage of sleep, stage 1, is the transition period from wakefulness to onset of sleep and may last 5-10 minutes. While awake, people show low-voltage EEG activity of 10-30 μV and 16-25Hz (Fig. 9.20). As they relax, they show alpha activity of 20M0μV and IOHz. During stage 1, the EEG shows a low-voltage mixed frequency pattern known as theta waves. Stage 2 contains theta waves plus K complexes and sleep spindles. Sleep spindles are sudden bursts of electrical activity, 12-14 Hz, lasting 1-2 seconds, whereas K complexes are sudden and sharp waves of electrical activity of longer frequency, often triggered by noise. Eye movements almost cease. During stage 3, the EEG contains large-amplitude, slow delta rhythms (0.5-2 Hz), and eye and body movements are absent. Delta waves are the slowest and strongest waves produced. Stage 4, the deepest stage of sleep, is characterized by large EEG rhythms of 2 Hz or less (slow wave sleep). Growth hormone, prolactin, and gonadotrophins are released during these last two stages, leading some to speculate that these stages of sleep are restorative. If awakened during stages 3 and 4, an animal will appear confused.
Throughout the night, there is a reduction in the duration of non-REM sleep, particularly stages 3 and 4, while there is an increase in the duration of REM sleep. During REM sleep, brain temperature and metabolic rate rise, although almost all skeletal muscle activity is lost.
During non-REM sleep, gamma-aminobutyric acid (GABA)-Containing neurons in the ventrolateral preoptic (anterior) area of the hypothalamus send a signal to the posterior hypothalamus, particularly the Iuberomammillary nucleus, which contains histamin- ergic neurons. These latter neurons normally project to the thalamus and cortex causing activation. The reticularis nucleus of the thalamus and thalamocortical neurons are reciprocally interconnected, and they shift into a non-REM sleep spindle wave when deactivated by the hypothalamus and brain stem. These cells have a unique property; when they are hyperpolarized, voltage-sensitive calcium channels open, causing a burst of action potentials. Following the calcium spike, the membrane potential returns to the hyperpolarized state. The action potential in the reticularis nucleus leads to the release of gamma aminobutyric acid (GABA) that hyperpolarizes thalamic cells projecting to the cortex (thalamocortical neurons), and such hyperpolarization causes a rebound low- threshold calcium spike in these thalamocortical neurons. The firing of the thalamocortical neurons causes synchronized postsynaptic potentials in cortical neurons that cause spindle waves. When the thalamocortical neurons become progressively hyperpolarized, and spindling becomes reduced, cortical neurons spontaneously develop delta waves.
Using brain transection and stimulation experiments, Francois Michel and Michel Jouvet showed that the pons is the source of EEG activation and rapid eye movement during REM sleep. The pons is also the site of inhibitory signals that lead to atonia. Signals called pontine-geniculate-occipital (PGO) waves are sent from the pons, specifically from the reticularis pontis oralis/caudalis region, to both the forebrain and the spinal cord (Fig. 9.21). These signals cause bursts of firing in cortical neurons while sensory input is suppressed via presynaptic inhibition. The cholinergic PGO-on cells fire in bursts, and this leads to the PGO spikes observed in the thalamus during REM sleep. Thus, the brain is blocked from external stimuli. During waking periods, serotonergic neurons from the raphe nucleus called REM-off cells hyperpolarize and block the burst firing of PGO-on cells. Noradrenergic neurons in the locus ceruleus, as well as hista- minergic neurons in the posterior hypothalamus, also are involved in inhibition of the PGO cells during waking periods.
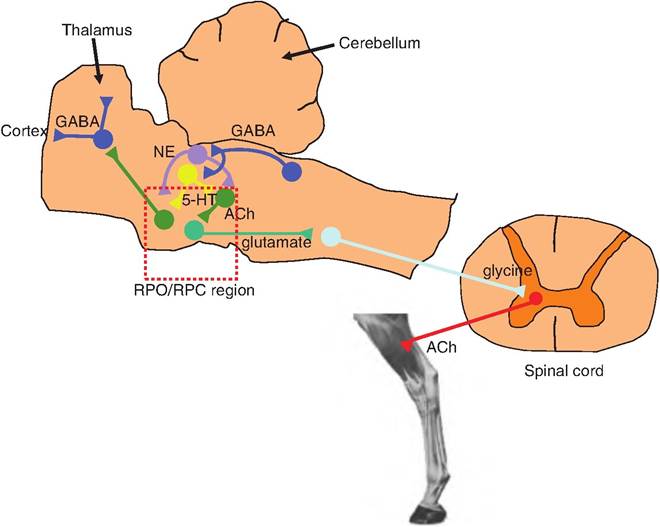
Fig. 9.21. Possible neurochemical model for the control of REM sleep. During the transition to REM sleep, the slow wave EEG associated with stage 4 of non-REM sleep is replaced by a low-voltage EEG. In addition, skeletal muscle tone is reduced. During this transition, GABA containing neurons in the pons become activated. They cause an inhibition of NE- and 5-HT-containing neurons found in the locus ceruleus and raphe nucleus, respectively. This causes disinhibition of the cholinergic neurons in the reticularis pontis oralis/caudalis (RPO/RPC) region of the pons. The ascending pontine neurons cause the reduction in EEG voltage associated with REM sleep by blocking the burst firing mode of the GABAergic neurons in the thalamus. The cholinergic neurons in the pons also stimulate descending glutamatergic neurons that project to the medulla, synapsing on glycine-containing neurons. These latter neurons project to the central gray area of the spinal cord and synapse on α-motor neurons causing inhibition. GABA, gamma aminobutyric acid; NE, norepinephrine; 5-HT, 5-hydroytryptamine.
Another class of neurons in the nucleus reticularis pontis oralis, the REM-on cells, shows little firing during waking and non-REM sleep, but has high activity during REM sleep.
Cranial nerves
Cranial nerves connect directly to the brain rather than the spinal cord (Fig. 9.22). Most are part of the peripheral nervous system, although the first two (olfactory nerve and optic nerve) are considered part of the CNS. There are 12 pairs of cranial nerves, and they arise from the ventrolateral surface of the brain. These nerves mostly innervate the head.
The cranial nerves attach to the brain near their associated sensory and/or motor nuclei. As shown in Table 9.1, the Cranialnerves can be classified as sensory, special sensory, motor, or mixed. Sensory nerves carry somatic sensory information such as touch, pressure, vibration, temperature, and pain input to the brain. Special sensory nerves carry signals associated with special senses including smell, sight, hearing, taste, or balance. Motor nerves carry somatic motor input to their respective muscles, while mixed nerves are both afferent and efferent, carrying sensory and motor information.
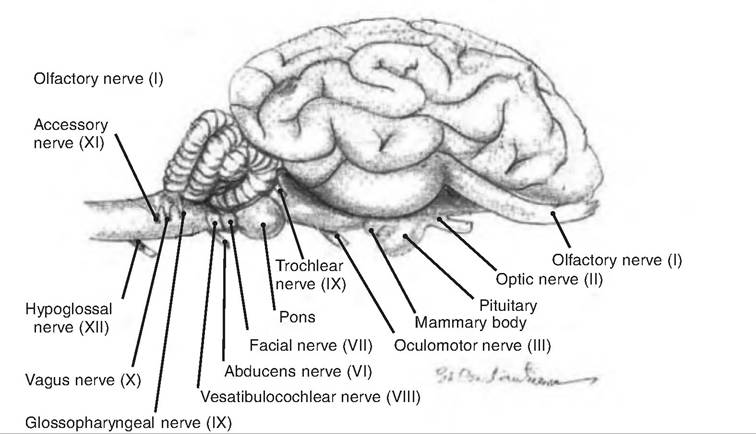
Fig. 9.22. Cranial nerves. Cranial nerves shown exiting the goat brain. (Reprinted from Constantinescu, 2001. Used by permission of the publisher.)
Table 9.1. Cranialnerves.
| Number | Name | Types of Axons | Function |
| Olfactory | Special sensory | Smell | |
| Il | Optic | Special sensory | Vision |
| III | Oculomotor | Somatic motor | Movements of eye and eyelid |
| Visceral motor | Parasympathetic control of pupil size | ||
| IV | Trochear | Somatic motor | Movement of the eye |
| V | Trigenimal | Somatic sensory | Sensation of touch to the face |
| Somatic motor | Motor control of mastication | ||
| Vl | Abducens | Somatic motor | Movement of the eye |
| Vll | Facial | Somatic motor | Facial expressions |
| Special sensory | Taste in anterior two-thirds of the tongue | ||
| Vlll | Vestibulocochlear | Special sensory | Hearing and balance |
| IX | Glossopharyngeal | Somatic motor | Movements in the throat |
| Special sensory | Taste in posterior two-thirds of the tongue | ||
| Visceral sensory | Detection of blood pressure (BP) changes in aorta | ||
| Visceral motor | Parasympathetic control of salivary glands | ||
| X | Vagus | Visceral sensory | Sense of pain in viscera |
| Visceral motor | Movement in the throat | ||
| Somatic motor | Parasympathetic control of heart, lungs and abdominal organs | ||
| Xl | Spinal accessory | Somatic motor | Movement in throat and neck |
| Xll | Hypoglossal | Somatic motor | Movement of the tongue |
Cranial nerves III, VII, IX, and X are associated with the parasympathetic nervous system. They carry autonomic signals to preganglionic fibers located in the periphery.
Cranial nerves are involved in cranial reflexes. These are reflexes involving sensory and motor fibers of the cranial nerves. Examples of these reflexes are shown in Table 9.2.
More on the topic Organization of the brain:
- Cognitive Architecture and Massive Modularity
- Albrecht Karl. Brain building: Easy games to develop your problem-solving skills. Prentice Hall Trade,1984. — 92 p., 1984
- Relations between Disciplines
- References
- Footnotes
- ABOUT THE CONTRIBUTORS
- REFERENCES
- Aggression as Instinct
- Three Classes of Models
- Introduction