Rodent Models
Small rodent models of paratuberculosis are usually utilized to reduce the cost associated with larger models of infection (e.g. cattle or goat models). In addition, the use of small animal models can successfully reproduce certain signs of paratuberculosis in a shorter period compared with ruminant models.
Mice represent the forefront small animal model for paratuberculosis. As for many infectious agents, mice provide a convenient model to study certain aspects related to the immunopathogenesis of infection with MAP. Given the impressive repertoire of immunological reagents and genetically distinct breeds (unlike ruminant models), mice were utilized efficiently to test the virulence of several MAP mutants, dissect the host-pathogen interactions and examine several vaccine candidates directed against paratuberculosis (Mullerad et al., 2002a; Shin et al., 2006; Bull et al., 2007; Chen et al., 2008; Settles et al., 2014; Ghosh et al., 2015). Another small animal, the white New Zealand rabbit (Beard et al., 2001a), is infrequently used. The leporid (rabbit) model is usually utilized to produce antibodies for researchers focused on paratuberculosis, but has gained more attention owing to its value in microbiota (Arrazuria et al., 2016a) and vaccination studies (Arrazuria et al., 2016b).16.2.1 Key features of the murine model of paratuberculosis
As often described among researchers in the field, mouse is the ‘poor man’s’ model for initial testing of vaccine candidates before investing more efforts and funds in more relevant models. Depending on the specific goals of the project, different protocols can be employed to examine MAP immune-pathogenesis or test novel vaccine candidates. In a typical vaccine study, candidate vaccines can be inoculated by the preferred route and then protection can be measured at later times (e.g. 6, 12 or 24 weeks) following challenge with a fully virulent strain of MAP.
These studies usually combine infection and immune-pathogenesis models based on the presence of control groups that receive only vaccines or only infection with a virulent strain of MAP. To evaluate the development of infection, sections from the liver, spleen and intestine are usually analysed for mycobacterial burden and histological lesions. In addition, immunological assessment can be done at the time of challenge with the virulent strain, as an attempt to uncover potential correlates of protection. Generally, a significant reduction of organ load with MAP (at least one log) and granuloma formation scores, compared with controls, are the major parameters for evaluation (Ghosh et al., 2014; Settles et al., 2014). These two outcomes depend on the genetic background of the mouse and the virulence of the mycobacterial challenge strain (Tanaka et al., 1994; Mullerad et al., 2002a; Shin et al., 2006). Some studies have used BALB/c or C57BL/6 mice, which are immunocompetent but susceptible to mycobacterial infection (Veazey et al., 1995). Others have used C3H mice, which are more resistant to MAP infection (Tanaka et al., 1994). Despite the presence of granuloma-like lesions in both susceptible and resistant breeds of mice, the number of lesions and bacterial colonization levels declined dramatically in the resistant breed. Alternatively, athymic nude mice inoculated via oral route with MAP resulted in bacterial shedding in the faecal materials of mice (Hamilton et al., 1991) - a rare clinical sign in mice.Another important parameter of the murine model is the dose of infection and the route of administration. Oral inoculation of 1011 colony-forming units (CFU)∕animal produces granulomatous lesions in 58% of the animals; however, lesions are limited to the mesenteric lymph node (Veazey et al., 1995). On the other hand, intra-peritoneal (IP) injection of a low dose of MAP (106 CFU) induces a small number of epithelioid granulomas, as compared with the multifocal granulomas composed of macrophages and epithelioid cells that are observed after a higher dose of infection (108 CFU) (Tanaka et al., 2000).
In our hands, an IP dose of 108 CFU/animal of virulent strains of MAP typically produces enough granulomatous responses to score by 3 weeks post-infection (WPI) and these lesions are well formed by 6 and 12 WPI (Shin et al., 2006; Wu et al., 2007b). In several reports, IP injection produced the highest reproducible infection rate (100%) compared with other routes of delivery (Mutwiri et al., 1992). In a high-throughput format, analysis is focused on the liver and intestine of inoculated animals. Both organs are analysed for histological lesions as well as mycobacterial burden over a period of 12 weeks (Shin et al., 2006). Recently, oral inoculation of immune-competent mice resulted in establishing MAP infection by 12 weeks postinoculation that required repeated inoculation of high doses of MAP (109 CFU/mouse, twice) (Cooney et al., 2014). More importantly, this oral infection elicited an immune profile in mice that is similar to that observed in cattle.Despite the several advantages of the murine model for MAP, several shortcomings limit the utility of this model. For example, there are known anatomical, physiological and
immunological differences between cattle and mice that could impact the outcome of infection. Also, some of the typical features of paratuberculosis in cattle (e.g. diarrhoea, severe intestinal lesions) cannot be reproduced in mice. None the less, the murine model for paratuberculosis may play an important role in vaccine development against paratuberculosis. Unlike the bovine, caprine or ovine models of paratuberculosis, many immunological reagents are well developed for mice, permitting a more mechanistic assessment of the histopathologic outcomes observed. Also, mice with variable genetic background are readily available allowing the dissection of host-pathogen interactions on a molecular level.
16.2.2 MAP infection models
Because MAP infection can require prolonged times from infection to clinical signs of paratuberculosis, mice offer the advantage that MAP-specific lesions can be observed within 3-6 weeks of infection (Tanaka et al., 1994; Shin et al., 2006).
Using immuno-compromised mice (SCID), intestinal lesions characteristic for paratuberculosis in cattle were obtained, including luminal infiltration with mononuclear cells containing acid-fast organisms (Mutwiri et al., 2001). Also, infected intestines show increased ion secretion and damage to the epithelial cell lining of intestinal villi, both potential reasons for the chronic wasting syndrome associated with paratuberculosis in infected cows. Factors associated with MAP invasion and attachment to intestinal cells, particularly M cells, were initially examined using the ileal loop of mice where mycobacterial FAP (fibronectin attachment protein) was analysed (Secott et al., 2002, 2004). Another set of virulence factors associated with how MAP regulates gene networks (sigma factors) was examined in mice (Ghosh et al., 2013, 2014) and showed that both sigL and sigH genes play a key role in MAP survival during infection. In these studies, at least 8 and 30 genes were under regulatory control of both sigL and sigH, respectively (Table 16.1).An attractive aspect of the murine model is the ability to utilize a large number of animals in an experiment with a relatively low budget. This economy of numbers is especially marked in comparison with ruminant models. In our experience, we have been able to assay the virulence of 11 MAP transposon mutants in BALB/c within a relatively short time (12 weeks of infection), enabling us to identify seven virulence genes in MAP, including pstA and papA2 (Shin et al., 2006). In a subsequent experiment, we assayed six additional mutants, again in a short time frame, permitting us to document the contribution of transcriptionally regulated genes (e.g. lipN, prrA) to MAP survival during infection. Further validation of the attenuation of some of these mutants was successfully performed for the ∆gcpE mutant (Wu et al., 2007a) and the ∆pstA mutant (Wu et al., 2009) in the cattle model, as well as the ∆lipN mutant in the caprine model (Shippy et al., 2017).
Results from the ruminant models validated the strategy for testing the virulence of MAP mutants first in the murine model before testing in any ruminant model of infection. It will be interesting to see whether mutants attenuated in ruminants are found for which no phenotype is detected in mice. Needless to say, mice were not able to reproduce exact organ lesions associated with paratuberculosis to those observed in ruminant models.With the advancement in murine genetics, several investigators took advantage of the available knockout strains of mice to investigate the role and type of immune responses directed against MAP infection. Studies of macrophages derived from mice with specific mutations on pathogen recognition receptors (e.g. TLR-2, TLR- 4, NOD2) have pointed to a role for innate immunity in the recognition and immune response directed against MAP (Ferwerda et al., 2007). For adaptive immunity, γδT cells were studied in γδT cell-knockout BALB/c mice to determine their role in granuloma formation (Tanaka et al., 2000). Another study, using C57BL/6 mice, demonstrated a major role for γ cells compared with γδ cells, by quantifying tissue lesions in mice deficient for the γ T-cell receptors (Stabel and Ackermann, 2002). Reconstitution of MAP- infected SCID mice with spleen cells from immunocompetent BALB/c mice significantly reduced lesions caused by MAP infection, indicating that protection against MAP is dependent on cell- mediated immunity (CMI) (Mutwiri etal., 2002).
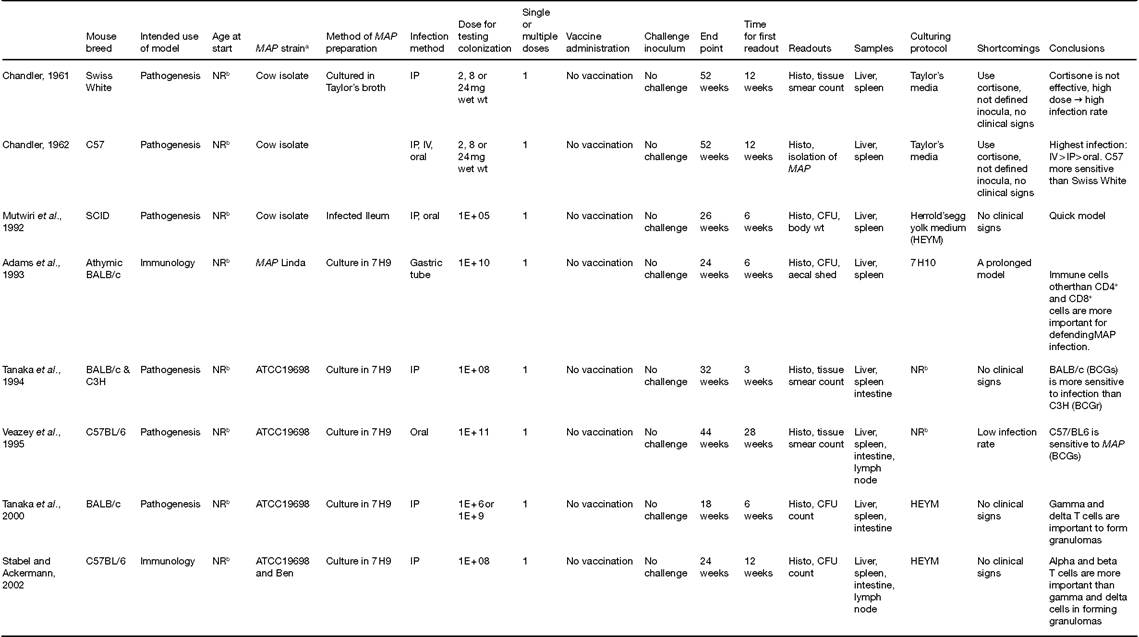
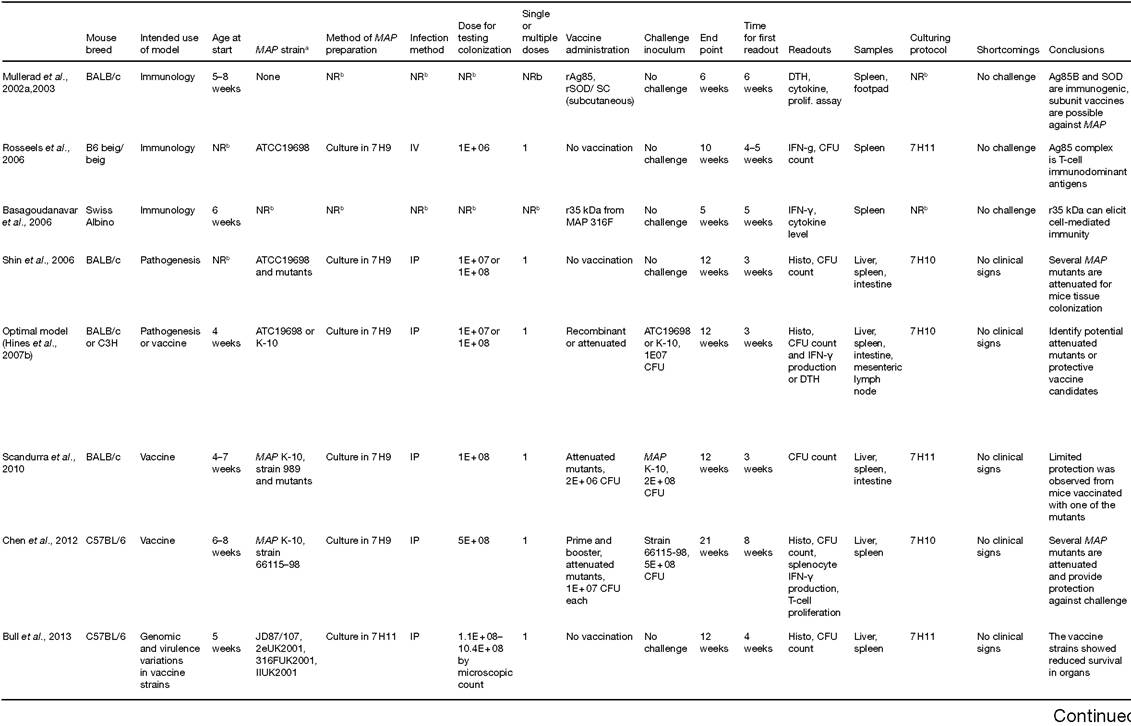
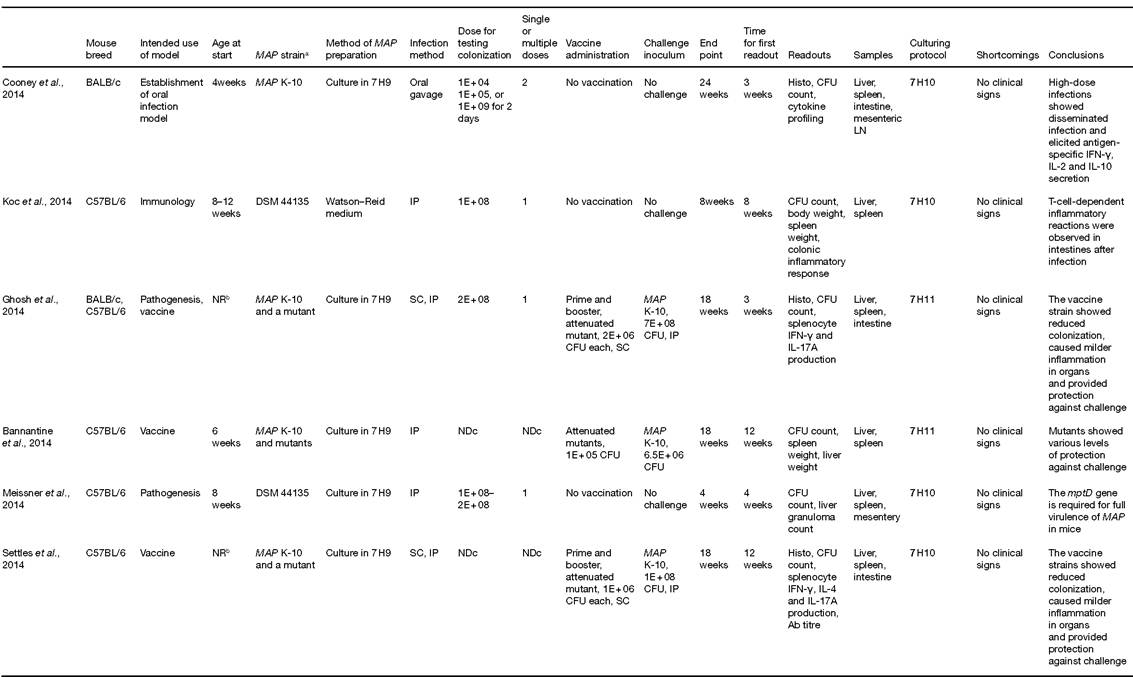
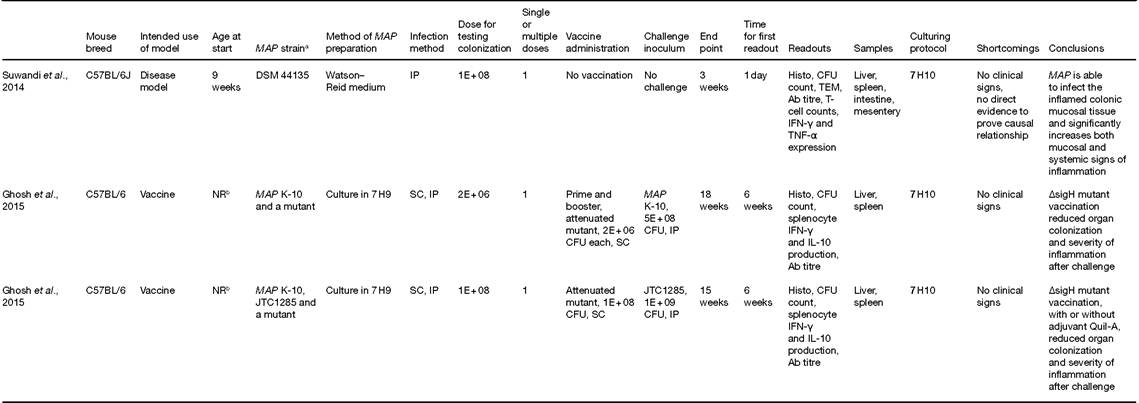
Table 16.1. Murine models for Mycobacterium avium subsp. paratuberculosis (MAP) infection.
A.M. Talaat et al.
Continued
A.M. Talaat eta∕.
Continued
3Where MAP strain was not provided, the provenance of the infecting strain is reported.
bNR = not reported.
cND = not determined.
CFU1 colony-forming units; SCID1 Severe combined immunodeficient; BCGs1 MycobaCterium bovis bacillus Calmette-Guerin (BCG) strains; LN1 Lymph node; SOD1 Superoxide dismutase; DTH1 Delayed-type hypersensitivity; Ab1 Antibody; IP1 Intraperitoneal; IV1 intravenous; SC1 subcutaneous.
By studying T-cell responses specific to the 35- kDa protein of MAP, the involvement of CMI in responding to infection with MAP was further documented (Basagoudanavar et al., 2006). Finally, murine models provided key information on the quality of adaptive immune responses. Both Th1 and Th2 responses were observed in mice inoculated with recombinant antigens (e.g. superoxide dismutase, SOD) (Mullerad et al., 2002a), opening the door for testing these antigens as vaccine candidates against MAP infection. In addition to paratuberculosis, the murine model is also used to investigate the potential link of MAP with human Crohn's disease. Although the casual relationship between the bacterium and the human disease remains debatable, MAP infection in mice was shown to worsen existing colitis that resembles Crohn's disease (Suwandi et al., 2014).
16.2.3 MAP vaccine models
Traditionally, the antigenicity of MAP-specific novel antigens was tested first in mice as a prelude to further analysis on naturally infected cattle (Paustian et al., 2004). This step-wise approach provides a paradigm for examining the functional significance of novel antigens and may also serve in the testing of novel vaccine candidates. Early on, several vaccine candidates (live-attenuated, subunit and vectored vaccines) developed against MAP were tested in the murine model of paratuberculosis (Table 16.1) before further testing in ruminant models. Recombinant proteins of Ag85B, SOD and thiol peroxidase (TP) of MAP elicited strong immune responses in mice (Mullerad et al., 2002a, b, 2003). In these models, subunit vaccine inoculation triggered strong levels of cytokines (e.g. IFN-γ, TNF-α, IL-4, IL-6) only in the presence of the Ribi adjuvant. Interestingly, a polypeptide encoding three open reading frames (ORFs) of MAP antigens provided protective responses against challenge with a virulent strain of MAP (Chen et al., 2008). Because DNA immunization does not require protein preparations (Huntley et al., 2005; Talaat and Stemke-Hale, 2005), a library of MAP encoding ORFs were analysed in mice (Huntley et al., 2005). A total of 26 protective antigens have been identified in such a screen. To take advantage of safety of the traditional vaccine vectors, an adenovirus-based vaccine (Ad5) encoding four fused antigens elicited strong immune responses against MAP using a strategy based on vaccination with DNA constructs followed by Ad5 inoculation of mice (Bull et al., 2007). This vaccine candidate was later further assessed and validated in a calf model of paratuberculosis, showing increased antigen-specific CD4+, CD8+ IFN-γ producing T- cell populations up to 7 months post-challenge (Bull et al., 2014).
Recently, more attention has been directed towards live-attenuated vaccine (LAV) candidates because of their ability to elicit a robust immunity against mycobacterial infections (Kaufmann, 2011). Some members among the group who established the AMSC standards also examined the utility of mice to screen eight LAV candidates in mice (Bannantine et al., 2014) to show that not all vaccine constructs need to be persistent to offer protection against paratuberculosis. Targeting specific gene deletions in extra-cytoplasmic sigma factors of MAP, sigH and sigL mutants were found to induce protective immunity in mice against challenge with virulent strain of MAP (Ghosh et al., 2013, 2014). When tested in goats, sigH mutant was also protective, similar to IipN mutant (Shippy et al., 2017). In all murine vaccine testing scenarios, the murine model was utilized as a preliminary screen of vaccine candidates to eliminate non-protective constructs. Whether the mouse model faithfully serves this purpose will require comparative experiments in different hosts.
16.2.4 Leporid (rabbit) models
Rabbits can be naturally infected with MAP (Judge et al., 2005) and provide an attractive model to study the immunopathogenesis of MAP. Historically, rabbits were used to generate polyclonal antibodies against MAP antigens. Both live and heat-killed MAP isolates were able to elicit strong immune responses in rabbits (Stabel et al., 1996). Several proteins expressed from clones of a genomic library of MAP were reactive to rabbit antibodies, including the 25-kDa protein (a.k.a Csp1) and Pks7 (Bannantine and Stabel, 2001). One of these antigens (Csp1) was shown to be expressed during macrophage survival. Recently, rabbits were also used to generate antibodies against a specific MAP antigen (34 kDa) that was shown to exist in naturally infected cows (Malamo et al., 2007). All of these reports suggested that rabbits can provide a valid model for generating antibodies necessary to investigate MAP pathogenesis. With the success of the leporid model in immune-diagnostics of MAP, this model was also investigated to study the pathogenesis of MAP. All models of MAP pathogenesis reported so far included oral administration of MAP with variable success in inducing infection in all of the animals (Mokresh and Butler, 1990; Mokresh et al., 1989; Vaughan et al., 2005). In one report, almost 50% of the inoculated animals did not show any sign of infection despite the high dose of infection (107 CFU/animal) (Mokresh et al., 1989). None the less, higher inocula (108 CFU) resulted in a higher infection rate (~70%) of inoculated rabbits (Mokresh and Butler, 1990). Interestingly, rabbits showing signs of the disease suffered from granulomatous enteritis and diarrhoea (Vaughan et al., 2005), which are hallmarks of paratuberculosis in cattle. Additionally, signs of body weight loss and clinical emaciation were also evident in a considerable number of inoculated rabbits (Vaughan et al., 2005). In all of these reports, MAP was isolated from variable tissues, especially the intestinal tract and mesenteric lymph nodes. However, faecal shedding of MAP was not always detectable in infected animals (Vaughan et al., 2005). To investigate factors that affect susceptibility to oral challenges of MAP in rabbits, dietary variations were applied 4 days prior to oral administration of MAP. The group given regular rabbit diet showed more signs, including low weight gain, faecal shedding, tissue colonization and histopathology, which are consistent with paratuberculosis infection, compared with groups switched to a high protein or high fibre diet (Arrazuria et al., 2015). These phenomena are thought to be partly attributed to shifted gut microbiota upon MAP infection, as evidenced by microbiome analysis of samples of caecum and sacculus ro- tundus from MAP-infected rabbits under different dietary regimens (Arrazuria et al., 2016a).
In a novel leporid model of vaccination study, heat-killed MAP vaccine was administrated 1 month before or after an oral MAP challenge (Arrazuria et al., 2016b). Both groups showed a elevated humoral response against MAP and reduced MAP colonization, but vaccination after infection provided a therapeutic effect as it showed slightly stronger protection compared with vaccination before infection. Overall, rabbits provide an important small animal model to study key aspects of paratuberculosis that cannot be investigated in mice. Aspects related to the induction of diarrhoea, weight loss or intestinal colonization could be better investigated in rabbits rather than mice. In most reports, rabbits developed signs of paratuberculosis within the first year post-infection, a time period that is much shorter than calf infections. Despite all of these advantages, the use of the leporid model in MAP research is limited because of the limited availability of immunological reagents and the expenses associated with housing rabbits compared with mice. See Table 16.2 for key aspects associated with both the murine and leporid models of paratuberculosis.
16.3