Sensory receptors
Sensations are the awareness of a stimulus, whereas perception involves the interpretation of the sensation and occurs in the CNS. All senses involve three steps: (1) a physical stimulus, (2) transformation of the stimulus into a nerve impulse, and (3) a response to the sensation in the form of a perception or conscious experience of sensation.
Furthermore, all sensory systems give four types of information about the stimuli, including modality, location, intensity, and timing, which collectively yield sensation (Table 10.1).The various modalities of sense include vision, hearing, touch, taste, smell, the vestibular sense of balance, and the somatic senses, including nociception (pain), temperature, itch, and proprioception (posture and movement of body parts). General senses include temperature, pain, touch, pressure, vibration, and proprioception; special senses include olfaction (smell), vision (sight), gustation (taste), equilibrium (balance), and audition (hearing). General sensory receptors are located throughout the body; receptors for special senses are located in specialized structures or organs. General senses are discussed later; special senses are covered in Chapter 11.
Classes of sensory receptors
There are five classes of sensory receptors: mechanical, chemical, nociceptors (nocere = to injure), thermal, and electromagnetic. Mechanoreceptors can detect touch, proprioceptive sensation (muscle stretch or contraction), joint position, hearing, and sense of balance. Chemoreceptors function in the sensing of itches, taste, and smell. Nociceptors detect pain. Thermoreceptors can sense either hot or cold; photoreceptors sense electromagnetic energy.
These receptors can be further classified by location as exteroceptors, interoceptors, or proprioceptors (from the Latin word proprius, meaning "belong to one's own self"):
1.
Exteroceptors. These are sensitive to stimuli outside (external) the body. Located near the surface of the body, they can detect touch, pressure, pain, and temperature, as well as special senses such as smell, taste, vision, and auditory.2. Interoceptors. These receptors monitor the visceral organs and their function. They monitor chemical and temperature changes, as well as stretching within the viscera. Although an animal is not normally consciously aware of their signals, they may produce pain signals, alerting the animal to a problem.
3. Proprioceptors. Although these receptors also respond to internal signals, they are restricted to those receptors in muscles and joints that provide information concerning the position of the bones and muscles.
General senses
Mechanoreceptors
Mechanoreceptors detect distortions in their cell membranes, such as bending and stretching. There are three classes of mechanoreceptors:
1. Tactile receptors. These are responsible for the sensations of touch, pressure, and vibration.
Table 10.1. Sensory receptors and modalities.
| Sensory System | Modality | Stimulus | Receptor Class | Receptor Cell Type |
| Auditory | Hearing | Sound | Mechanoreceptors | Hair cells (cochlea) |
| Visual | Vision | Light | Photoreceptors | Rods and cones |
| Vestibular | Balance | Gravity | Mechanoreceptors | Hair cells (vestibular labyrinth) |
| Somatosensory | Somatic Senses: Touch Proprioception Temperature Pain | Pressure Displacement Thermal Chemical, thermal, or mechanical | Mechanoceptors Mechanoceptors Thermoreceptors Chemoreceptors, thermoreceptors, or mechanoreceptors | III, IV, VI, VII, X and cervical nerves |
| Gustatory | Taste | Chemical | Chemoreceptors | Taste buds |
| Olfactory | Smell | Chemical | Chemoreceptors | Olfactory sensory neurons |
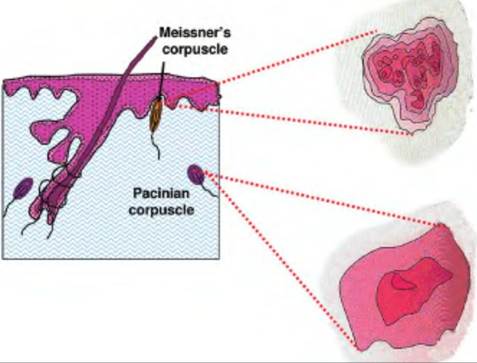
Fig.
10.4. Receptive field. Meissner's corpuscles have a smaller receptive field compared with the more deeply located Pacinian corpuscles.2. Baroreceptors (baro = pressure). These detect changes in pressure in the walls of blood vessels, as well as the digestive, reproductive, and urinary tracts.
3. Proprioceptors. These detect changes in the position of joints and muscles.
Tactile receptors
Named after the German and Italian histologists who discovered them, the two principal mechanoreceptors located in the superficial skin layers are MerkeTs discs and Meissner's corpuscles. The Merkel disc receptor is a slowly adapting receptor consisting of a small epithelial cell surrounding a nerve terminal. They are involved in the sense of touch and pressure. Meissner's corpuscles are rapidly adapting and consist of a globular, fluid-filled structure enclosing a stack of flattened epithelial cells around which the sensory nerve is entwined.
The two mechanoreceptors found in the deep subcutaneous layers are the Pacinian corpuscle (Fig. 10.4) and the Ruffini ending. Although larger than the receptors found in the superficial layers, these receptors are less numerous. The Pacinian corpuscle, physiologically similar to the Meissner's corpuscle, is a large receptor measuring as long as 2 mm and nearly 1 mm in diameter.
The Pacinian corpuscle, also called a large Iamel- Iated corpuscle, is fast-adapting and responds to rapid indentation of the skin but not steady pressure. Since the capsule surrounding this receptor is attached to the skin, this receptor can sense low-frequency vibration occurring several centimeters away. These receptors are activatedby touching a tuning fork (200-300 Hz) to the skin or bony structure.
Ruffini endings, slightly smaller than Pacinian corpuscles, are slow-adapting. They consist of a capsule surrounding a core of collagen fibers that are continuous with fibers in the surrounding dermis. Dendrites from the sensory neuron are intertwined with the collagen fibers in the capsule.
They link the subcutaneous tissue with folds in the skin at the joints and nails. Thus, they sense stretch of the skin bending in these regions.Vibration is the detection of sinusoidal oscillations of objects in contact with the skin. The various tactile receptors differ in their sensitivity to vibration. Merkel's discs are most responsive to low-frequency oscillations (5-15Hz). Meissner's corpuscles are sensitive to midrange oscillations (20-50 Hz), and Pacinian corpuscles to high frequencies (60-400 Hz). The lowest stimulus intensity to which a receptor produces an action potential is the receptor's tuning threshold. The intensity of the vibration is coded by the number of sensory nerve fibers that are firing rather than the frequency of action potentials within a fiber.
The size of the receptive fields for the various touch receptors differs. Meissner's corpuscles and Merkel's discs, located in the superficial skin layers, have small receptive fields. A single dorsal root ganglion cell innervates 10-25 of each of these receptors and produces a receptive field of 2-10 mm in diameter. Therefore, these receptors are responsible for a fine discriminating touch that detects small spatial differences. These receptors are important in a two- point discrimination test. When performed on people, the skin is simultaneously touched by two pointed objects, and the person is asked whether they can detect two objects or a single object. As the two points are moved closer together, the person will eventually be unable to discriminate between the two objects.
In contrast, each Pacinian corpuscle and Ruffini ending, located in deeper skin layers, is innervated by a single nerve fiber, but its receptive field is large since these receptors can detect changes from mechanical displacement over a large distance. Because of their large receptive fields, these receptors are involved in coarse resolution of touch. The combination of the adaptation rate and size of the receptive field leads to variations in how the tactile receptors respond to stimuli (Fig.
10.5).In addition to differences in receptive fields of the various tactile receptors, there are also differences in the number of these receptors located throughout the body. The smallest receptive fields, that is, those with the most receptors, are located in the tips of the paws and whiskers.
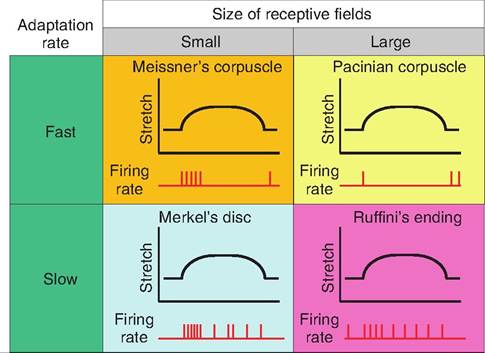
Fig. 10.5. Response of tactile receptors to stretch. The various tactile receptors change their firing rate qualitatively and quantitatively in response to stretch. Whereas Meissner's and Pacinian corpuscles respond quickly but also adapt quickly, Merkel's discs and Ruffini's endings adapt more slowly.
Baroreceptors
Consisting of free nerve endings, baroreceptors sense change in the walls of distensible organs, including blood vessels, and a portion of the respiratory, digestive, and urinary tracts. When the pressure in the walls of these organs increases, the walls are stretched, causing a deformation in the sensory nerves. As the pressure in these organs decreases, the elastic fibers cause the walls to return to their original structure.
Baroreceptors monitor blood pressure in the major vessels, particularly in the carotid artery at the carotid sinus, and the aorta at the aortic arch. Increases in blood pressure at these sites initiate the baroreceptor reflex, in which increased firing rates from these baroreceptors is relayed to the CNS and appropriate adjustments in heart rate and blood pressure are initiated. Baroreceptors in the lungs send information regarding lung inflation to the respiratory rhythmicity centers in the brain stem to regulate breathing. Similarly, baroreceptors in the colon and urinary bladder function in defecation and micturition, respectively. There are also baroreceptors along the gastrointestinal tract (GI tract) that are involved in peristalsis.
Proprioceptors
Proprioceptors monitor the position of joints, and the tension in tendons, ligaments, and muscle.
These receptors do not adapt, and they continuously send information to the CNS. There are three groups of proprioceptors:1. Muscle spindles. Discussed in Chapter 8, these receptors detect the length of skeletal muscles.
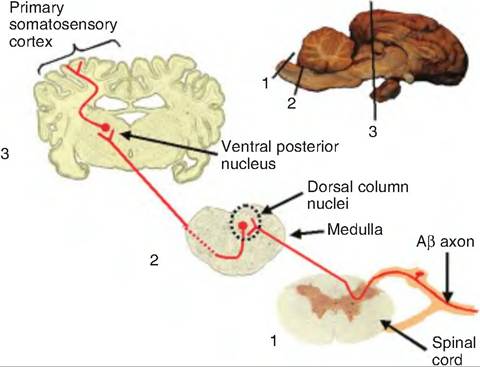
Fig. 10.6. Dorsal column-medial lemniscal pathway. Mechanoreceptor signals enter the spinal cord through the dorsal root where they either synapse on secondary fibers in the gray area or ascend in the dorsal column. Fibers ascending in the dorsal column synapse in the dorsal column nuclei (nucleus gracilus and cuneatus). Fibers from these nuclei immediately decussate and ascend to the ventral posterior nucleus of the thalamus via the medial lemniscus. From there, fibers ascend to the primary somatosensory cortex.
2. Golgi tendon organs. Located at the junction between skeletal muscle and its tendon, the Golgi tendon organs detect stretch of the tendons. The dendrites of the receptor neurons branch extensively, wrapping around the collagen fibers of the tendon.
3. Receptors in joint capsules. In order to monitor the position of the body, joint capsules are innervated with free nerve endings that detect pressure, tension, and movement of the joint.
Dorsal column-medial lemniscal pathway (mechanoreceptor pathway)
Axons of skin sensory receptors are designated, in order of decreasing size, as Act, Aβ, Aδ, and C, which correspond to axons innervating muscles and tendons called groups I, II, III, and IV, respectively. Act, Aβ, and Aδ are myelinated; C fibers are unmyelinated. Sensory nerves from the skin do not have Act fibers. Mechanoreceptors send their messages via Aβ. These fibers enter the dorsal horn of the central gray area of the spinal cord and branch. One branch synapses on second-order sensory neurons deep in the dorsal horn and are involved in reflexes. The other branch ascends to the brain in the dorsal column-medial lemniscal pathway (Fig. 10.6). This branch enters the ipsilateral dorsal column of the spinal cord. Composed of primary sensory axons, as well as second-order axons from neurons in the central gray area of the spinal cord, these fibers ascend to the dorsal column nuclei at the junction of the spinal cord and medulla where they synapse. The fibers leaving the dorsal column nuclei decussate (cross to the other side) and ascend in the medial lemniscus that courses through the medulla, pons, and midbrain, and synapses in the ventral posterior nucleus of the thalamus.
Nociceptors
Pain is mediated by nociceptors. These receptors respond to stimuli that can damage tissue. Some nociceptors respond directly to stimuli, and others respond indirectly since they respond to chemicals released by damaged tissue. The list of chemical intermediates includes histamine, K+, and proteases released from injured cells, bradykinin, substance P, acidity, adenosine triphosphate (ATP), prostaglandins, serotonin, and acetylcholine (ACh).
Chemical mediators are released in response to various stimuli. Bradykinin is produced when peptidases released from injured cells cleave the extracellular protein kininogen. Bradykinin acts directly on nociceptors, and it also increases the synthesis and release of prostaglandins from nearby tissue. Tissue acidity can increase when, for example, a galloping horse begins anaerobic metabolism producing lactic acid that leads to an increase in extracellular H+ ions. Histamine is released when mast cells found in the connective tissue are stimulated, such as during a bee sting. Prostaglandin E2 is a metabolite of arachidonic acid and is generated by the enzyme cyclooxygenase released from damaged cells. Aspirin and other nonsteroidal anti-inflammatory analgesics work by blocking cyclooxygenase and inhibiting the synthesis of prostaglandins.
There are three classes of nociceptor. Mechanical and thermal nociceptors are sensitive to mechanical and thermal stimuli, respectively, whereas polymodal nociceptors respond to traumatized tissue rather than physical properties. Mechanical nociceptors respond to a strong tactile or sharp penetrating stimulus that results in pain. Their firing rate increases with the destructiveness of the mechanical stimuli. Thermal nociceptors respond to extremes in temperature. One group responds to noxious heat above 45oC; the second group responds to noxious cold below 5oC. The polymodal nociceptors respond not only to painful mechanical stimuli such as a strong pinch or puncture, but also to noxious heat and cold and irritant chemicals. Stimulation of these receptors evokes slow, burning pain. These are the primary receptors in tooth pulp.
Pain signals are carried by lightly myelinated Aδ and unmyelinated C fibers. These fibers have different conduction velocities; therefore, pain information can
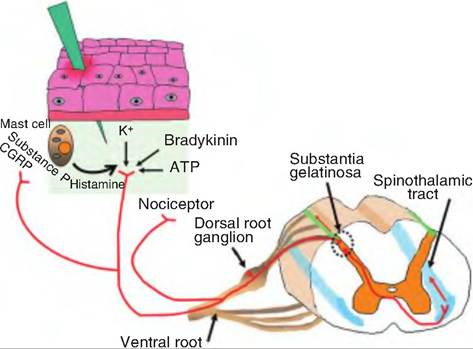
Fig. 10.7. Nociception. Damage to the skin can cause release of certain substances, including ATP, prostaglandins, and bradykinin. These substances can stimulate the nociceptors. Collaterals of the receptors can release substance P and calcitonin gene-related peptide (CGRP) that can stimulate mast cells to release histamine. Histamine can stimulate nociceptors as well as cause vasodilation. Hence, the redness associated with inflammation. The pain signals are carried by Aδ and C fibers through the dorsal root to the substantia gelatinosa of the central gray area of the spinal cord. Here the fibers release glutamate and/or substance P, which signals secondary fibers. The secondary fibers cross to the contralateral side and synapse on neurons that carry the impulse to the brain via the spinothalamic tract.
produce two different kinds of pain perceptions (Fig. 10.7). The first pain is a fast, sharp pain mediated by Aδfibers; it is followed by secondary pain that is duller, but longer lasting and mediated by C fibers. Once stimulated, branches of the nociceptor neurons can secrete substance P and calcitonin gene-related peptide, peptide neurotransmitters, from peripheral terminals of collaterals of the primary nociceptive neurons. These two peptides can cause vasodilation as well as release of histamine from mast cells.
Heat, redness, swelling, and pain are the cardinal signs of inflammation. Substance P can cause all of these symptoms. The heat and redness are caused by the vasodilation, swelling is caused by the leakage of proteins and cells from these blood vessels into the interstitial space, and the pain can result from the induced release of histamine that stimulates nociceptors.
Pain pathway
The Aδ and C fibers enter the spinal cord through the dorsal root and synapse in the substantia gelatinosa of the central greay area. The neurotransmitters released at this site are thought to be glutamate and substance P The glutamate, released from Aδ and C fibers, acts at AMPA-type glutamate receptors
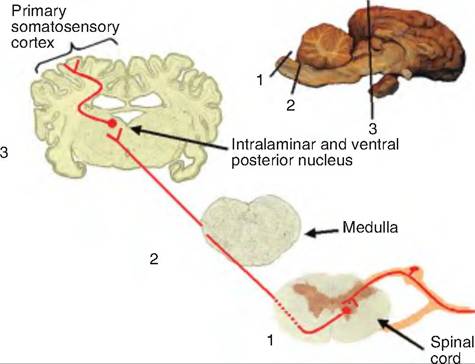
Fig. 10.8. The spinothalamic pathway. Pain and temperature information is carried to the brain via the spinothalamic pathway. Sensory fibers enter the spinal cord by way of the dorsal root and synapse in the central gray area of the spinal cord. They synapse on fibers that decussate and then ascend through the spinothalamic tract to the intralaminar and ventral posterior nuclei of the thalamus, where they synapse on fibers that then course to the primary somatosensory cortex.
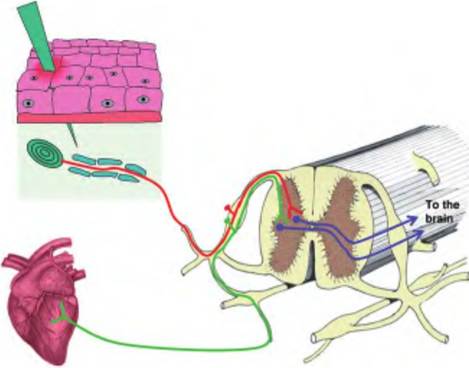
Fig. 10.9. Referred pain. Referred pain occurs when visceral nociception is perceived as a cutaneous sensation. The classic example of this is angina, in which the heart receives insufficient oxygen resulting in pain from this region. However, the body perceives the pain as coming from the upper chest or left arm since cutaneous sensations from this region synapse in the spinal cord near the same region, and the brain is unable to distinguish between the two.
and evokes fast synaptic potentials in dorsal horn neurons. Substance P is released from C fibers and evokes slow excitatory postsynaptic potentials (EPSPs). Glutamate and substance P act together to transmit pain signals and, with substance P, enhance and prolong the actions of glutamate. The impulse is then carried by secondary fibers that immediately decussate and then ascend to the brain via the spinothalamic tract (Fig. 10.8).
Many times, shortly after an injury, the site becomes extremely painful and especially sensitive to touch. This is called hyperalgesia and is the body's way of protecting this site from further injury. Primary hyperalgesia is associated with the damaged tissue; however, the surrounding area can also become supersensitive, a process called secondary hyperalgesia. Hyperalgesia is due to the action of various compounds released during injury that make the nociceptors more sensitive.
Nociceptors from the viscera also enter the spinal cord by the same route as those from cutaneous nociceptors. These signals can get mixed within the spinal cord since the afferent fibers from the viscera and somatic area converge on the same projection neurons in the dorsal horn of the spinal cord (Fig. 10.9). This leads to the phenomenon of referred pain in which visceral pain is perceived as a cutaneous sensation. Such is the case with angina, in which ischemia in the heart leads to pain in the upper chest and down the left arm.
Thermoreceptors
Thermoreceptors alter their firing rate as a result of changes in temperature. Unlike mechanoreceptors that are silent in the absence of stimuli, thermoreceptors maintain a low, tonic firing rate (2-5 spikes ∕s) at normal body temperature. There are separate cold and warm receptors, which can be shown by differential mapping on the skin. These free nerve endings are located in the dermis of the skin, skeletal muscles, liver, and hypothalamus.
Warm receptors begin firing around 30oC, increasing their firing rate up to 45oC, after which their firing rate decreases (Fig. 10.10). Above 50oC, warm receptors stop firing. Instead, the animal senses heat pain rather than warmth due to the firing of thermal nociceptors. Cold receptors actively fire at temperatures ranging from 35oC down to 10oC. Below this temperature, cold becomes an anesthetic. For unknown reasons, some cold receptors increase their firing rate above 45oC.
Thermoreceptors are very sensitive to differences in temperature between an object being touched and skin temperature. They respond vigorously to these abrupt changes in temperature and then adapt their firing rate. This can be demonstrated by placing your hand in a beaker of cold water. Notice that with time, the sensations of cold decrease. Then, take your hand and plunge it into a beaker of warm water. Notice that the water will actually feel hot due to the sudden change in temperature.
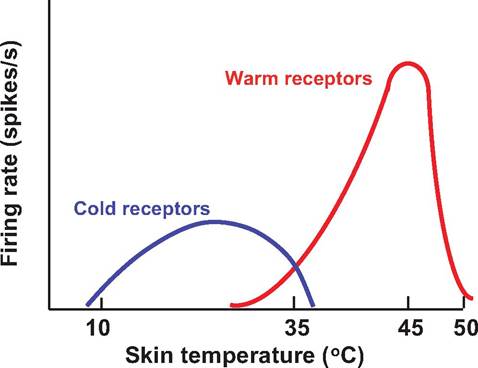
Fig. 10.10. Thermoreceptors. Thermoreceptors change their firing rate in response to changes in skin or organ temperature. At normal body temperatures (36-38oC), both cold and warm receptors are discharging. As skin temperature decreases below 30oC, warm receptors discontinue firing, whereas cold receptors increase their firing rate, which is maximal at 25oC. Warm receptors fire maximally at 45oC.
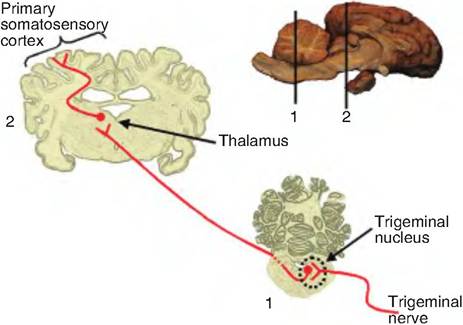
Fig. 10.11. Trigeminal pathway. Pain and temperature information from the face send information via the trigeminal nerve (cranial nerve V). This information is carried to the trigeminal nucleus and synapses on second-order neurons that decussate and ascend to the thalamus where they synapse on neurons and then ascend to the primary somatosensory cortex.
Thermoreceptor pathway
Cold receptors connect to Aδ and C fibers, whereas warm receptors connect only with C fibers. These fibers synapse in the substantia gelatinosa of the dorsal horn in the spinal cord. The secondary fibers then decussate and ascend in the contralateral spinothalamic tract along with the pain signals.
Pain and temperature information from the face and head reaches the thalamus via the trigeminal pathway (Fig. 10.11). Fibers in the trigeminal nerve synapse on
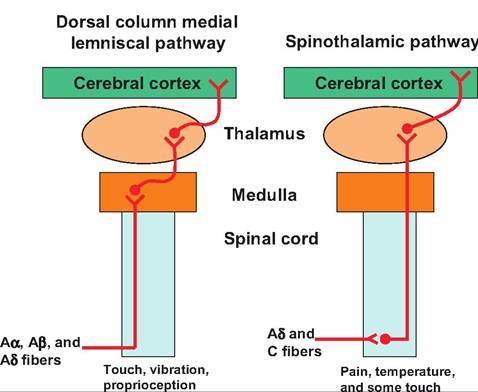
Fig. 10.12. Pain and temperature pathways. The two major ascending pathways that carry pain and temperature information include the dorsal column-medial lemniscal pathway and the spinothalamic pathway. Note that the dorsal column medial lemniscal pathway enters the spinal cord through the dorsal root and ascends to the medulla where it synapses on second-order neurons that cross over to the contralateral side and then ascend to the cerebral cortex. In the spinothalamic tract, fibers enter the spinal cord via the dorsal root and synapse on second-order fibers in the central gray area. These second-order neurons cross over to the contralateral side before ascending to the thalamus where they synapse on neurons that then project to the cerebral cortex.
second-order neurons in the spinal trigeminal nucleus in the brain stem. These fibers decussate and ascend to the thalamus in the trigeminal lemniscus. The pain and temperature pathways are summarized in Figure 10.12.
Chemoreceptors
Chemoreceptors are responsible for detecting changes in concentrations of specific chemicals or compounds. These receptors are also responsible for the special senses of taste (gustation) and smell (olfaction). Taste and olfaction, which are considered special senses whose signals are relayed to the primary sensory cortex, will be discussed in a separate chapter. The chemoreceptors whose signals do not travel to the primary sensory cortex will be discussed here. They are responsible for sensing irritating substances on the skin, or nutrients within the GI tract or brain, and carbon dioxide or oxygen levels in our blood.
There are chemoreceptors in the respiratory centers of the brain that sense changes in H+ and CO2 concentrations. There are also chemoreceptors in the carotid bodies located near the origin of the internal carotid arteries and in the aortic bodies found between the major branches of the aortic arch. These receptors respond to changes in blood pH, CO2, and oxygen concentrations. Signals for the carotid and aortic bodies travel to the respiratory centers through the glossopharyngeal (cranial nerve IX) and vagus (cranial nerve X).
Detection of sensory signals
Sensory receptors are morphologically specialized structures that respond to specific stimuli. When stimulated, these receptors transform the stimulus into an electrical signal called a receptor potential, which is a graded potential causing either depolarization or hyperpolarization of the cell. The amplitude and duration of the receptor potential are related to the magnitude and length of time of the stimulus. If the receptor potential is large enough to reach threshold, it is called a generator potential, and causes an action potential to form in the sensory neuron. The process of converting the stimulus into a receptor potential is called stimulus transduction.
The various types of sensory receptors have different mechanisms for transducing the stimulus into a receptor potential. In mechanoreceptors, when there is a conformational change in the tissue in which the receptor resides, it causes a change in the plasma membrane of the mechanoreceptor, thus causing a physical change in the cation channels located in the sensory neuron membrane (Fig. 10.13). This physical change results in the opening of stretch-sensitive channels that increase ion conductance. This leads to depolarization of the neuron and generation of a receptor potential. This mechanism is very similar to the production of an EPSP. The amplitude of the receptor potential is proportional to the intensity of the stimulus. Greater conformational change in the tissue will result in a greater number of channel openings on the mechanoreceptor. When the stimulus is removed, the ion channels close.
Receptors involved in general senses are either free dendritic endings or encapsulated dendritic endings. Free nerve endings are located throughout the body, but are especially abundant in epithelia and connective tissue. They are nonmyelinated and end in a knob-like swelling, sensitive to touch and pressure. There appears to be no structural differences between those that detect touch and pressure and those that detect temperature and pain. While these are the only sensory receptors on the surface of the eye, there are specialized tactile receptors located throughout the body surface that are probably more important. A summary of the various sensory receptors is shown in Table 10.2.
Surrounding all hairs is a root hair plexus that monitors distortions and movements. Movement of the hair causes a distortion in the sensory dendrites, resulting in the production of a receptor potential. These are rapidly adapting receptors. Therefore, they are most important for detecting initial movements or changes. For example, when a saddle is first placed on a horse, these receptors send signals to the brain notifying the presence of the saddle, but rather quickly these receptors adapt and stop sending signals.
Reflexes
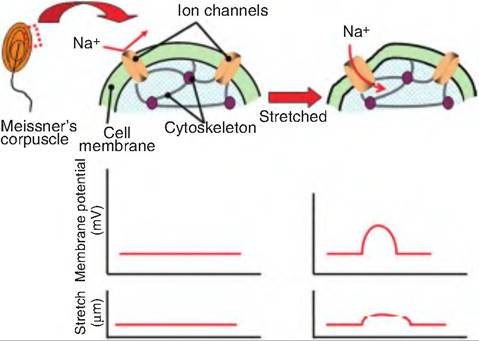
Fig. 10.13. Mechanoreceptor depolarization. Mechanoreceptors respond to physical deformation of the plasma membrane. In this example, a Meissner's corpuscle, located in the skin, is deformed when the skin is pressed. This produces a physical change in the corpuscle membrane, which causes an opening of an ion channel on the plasma membrane. As cations move inward, a receptor potential is generated in the corpuscle.
Reflexes are automatic, neural responses to specific stimuli. Reflexes work to preserve homeostasis by making rapid adjustments that do not require conscious activity. Therefore, all reflexes involve a sensory receptor and a motor response.
The neural path controlling a reflex is called a reflex arch (Fig. 10.14). The reflex arch begins with a sensory receptor and ends with the effector. There are four steps to a reflex arch:
1. Stimulus activates receptor. Sensory receptors, such as those described earlier, receive a stimulus that generates a receptor potential, resulting in the production of an action potential in the receptor. If the sensory cell and sensory neuron are separate, the action potential is produced in the sensory neuron.
2. Information processing. The sensory information is transmitted to the CNS where it is processed. In a monosynaptic reflex, the sensory neuron synapses directly on a motor neuron, releasing an excitatory neurotransmitter and causing the
Table 10.2. Sensory receptors: structure and function.
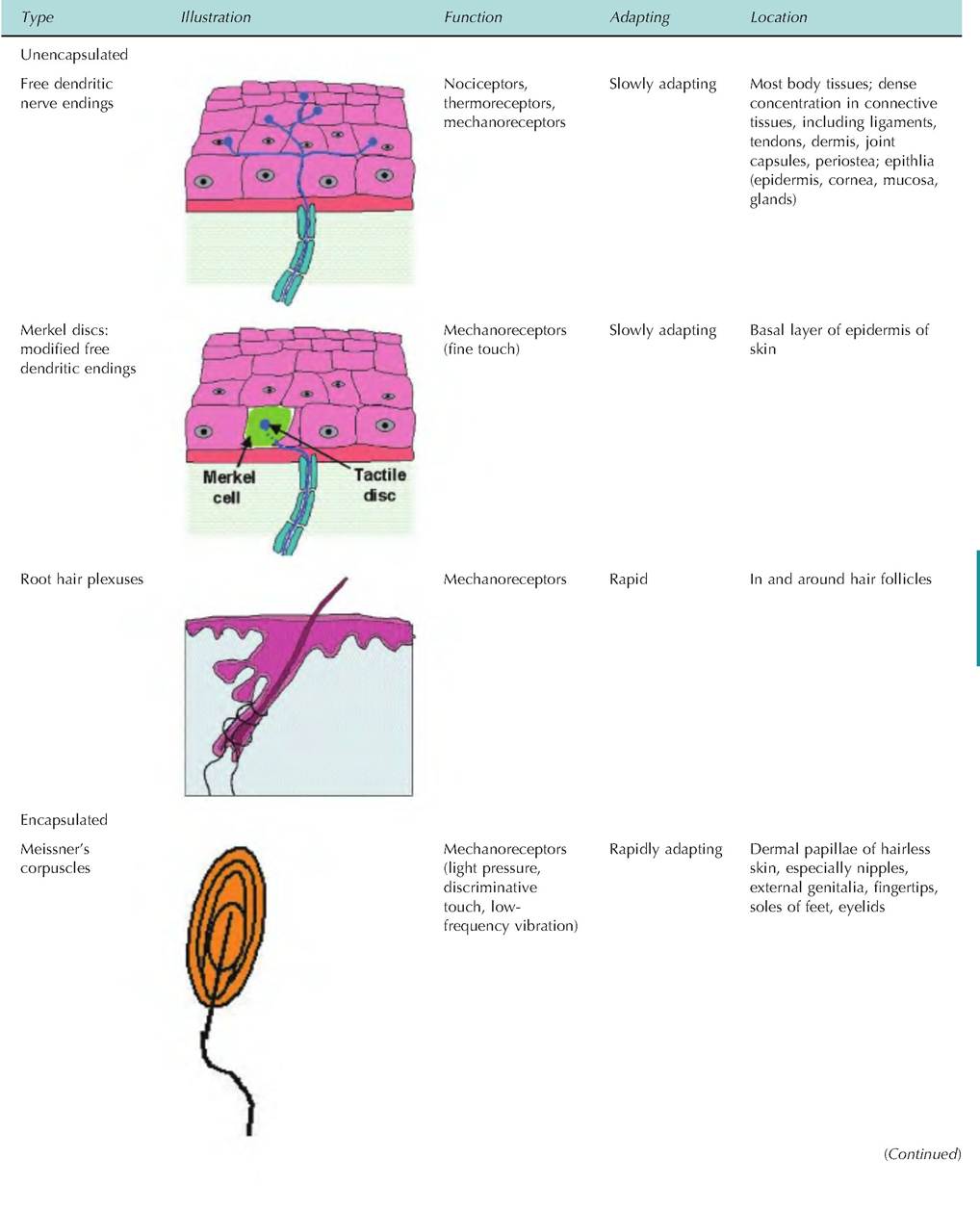
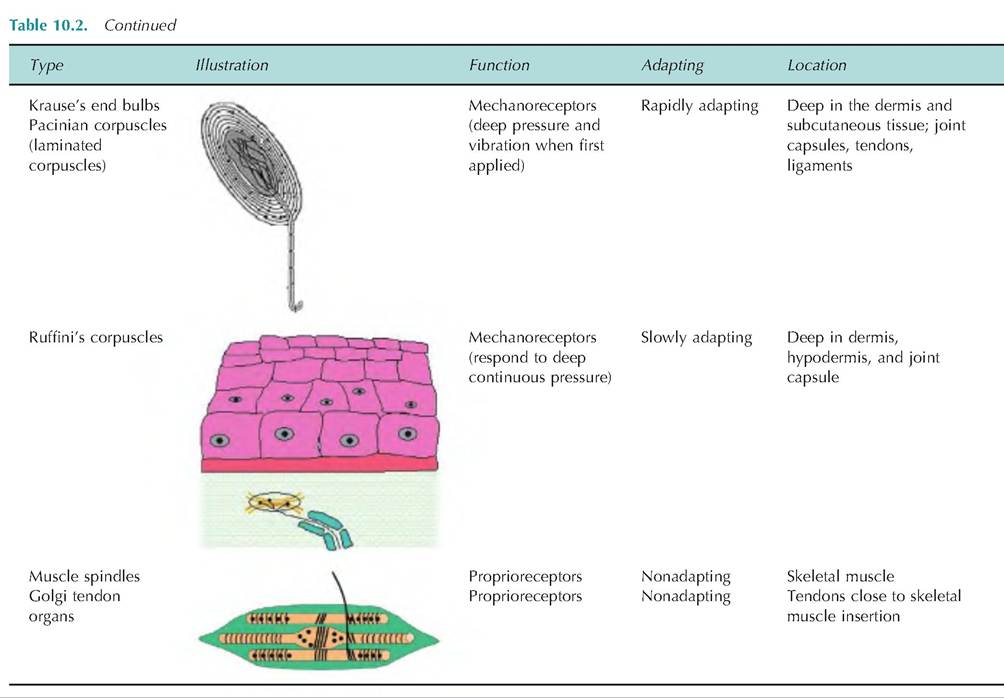
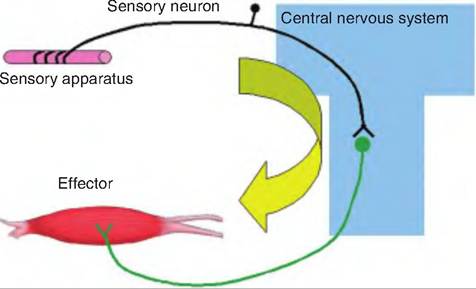
Fig. 10.14. Reflex arch. A reflex arch includes a sensory apparatus, a sensory neuron, a site of information processing or integration, and an effector. The sensory apparatus detects a change that is transmitted via the sensory neuron to the central nervous system. There, the information is processed and a motor response is sent via a motor neuron to the periphery in order to maintain homeostasis.
production of an EPSP in the motor neuron. Under normal conditions, EPSPs in motor neurons always result in an action potential in the motor neuron. In a polysynaptic reflex, there are pools of interneurons generally carrying signals to multiple sites (Fig. 10.15). These reflexes generally involve both excitatory and inhibitory neurotransmitters being released from various interneurons (i.e., excitatory interneurons and inhibitory interneurons).
3. Activation of the motor neuron. As a result of the stimulus, a motor neuron is activated to cause contraction in order to establish homeostasis. In the case of polysynaptic reflexes, there are other synergistic muscles that may be stimulated and antagonistic muscles that are inhibited.
4. Response of the peripheral effector. Stimulation of the motor neurons results in release of a neurotransmitter at the synapse between the motor neuron and its effector. This results in contraction of a single muscle in the case of a monosynaptic reflex, or multiple muscles in the case of a polysynaptic synapse.
Classification of reflexes
Reflexes can be classified based on their development, site of information processing, or resulting motor response.
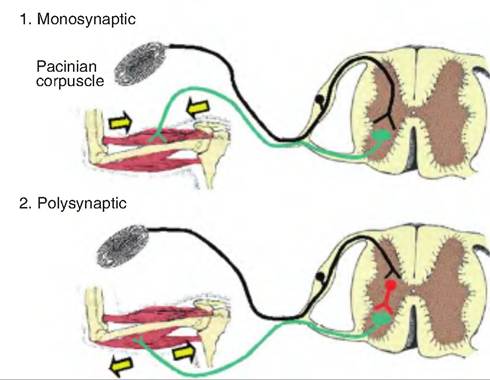
Fig. 10.15. Monosynaptic and polysynaptic reflexes. 1. In monosynaptic reflexes, the sensory neuron is activated and carries a signal to the central nervous system where it synapses on an effector neuron. The effector neuron is activated causing an effect to occur in the periphery. 2. In a polysynaptic reflex, the sensory signal synapses on an interneuron in the central nervous system. The interneurons process the information and then synapse on an effector neuron. The interneurons can either be excitatory, causing activation of the effector neuron, or they can be inhibitory, thus decreasing the firing rate in the effector neuron.
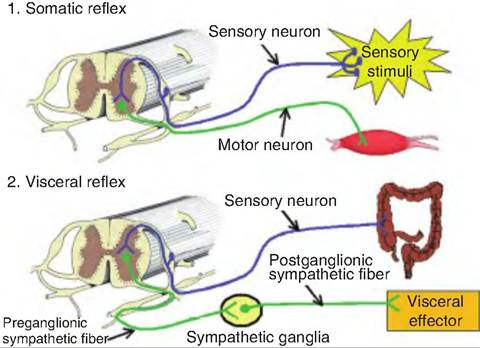
Fig. 10.16. Somatic and visceral reflexes. 1. Somatic reflexes involve a somatic sensory signal that is carried to the central nervous system where it stimulates a somatic motor neuron whose fiber travels to a skeletal muscle and causes contraction. 2. A visceral reflex is polysynaptic. There is either a somatic sensory signal or a visceral sensory signal that is relayed to the central nervous system where it stimulates a presynaptic autonomic fiber, which then exits the central nervous system and synapses on a postsynaptic fiber that then stimulates a visceral effector (i.e., smooth muscle, cardiac muscle, or gland).
Development of reflex
Animals are born with some reflexes called innate reflexes. Such reflexes involve the development of a genetically programmed response to specific stimuli. Such reflexes generally entail some process vital for life. Examples of such reflexes include suckling, chewing, a withdrawal reflex from painful stimuli, and tracking objects with the eyes. A newborn calf will generally begin suckling almost immediately after birth.
Other reflexes may develop later in life as a result of experience, and are called acquired reflexes. An example of an acquired reflex might be salivation in response to the sound of a bell. Like innate reflexes, these responses are quick, automatic, and stereotypic, but they must be learned.
Site of information processing
If the sensory information is processed in the spinal cord, the reflex is called a spinal reflex (e.g., withdrawal reflex). Such reflexes do not require input or processing from the brain; therefore, they are functional in decerebrate animals. The processing of information in the brain results in cranial reflexes.
Resulting motor response
Reflexes that involve the contraction of skeletal muscle are termed somatic reflexes; those that involve smooth muscle, cardiac muscle, or glands are called visceral reflexes (Fig. 10.16). Although the contraction of skeletal muscle is generally under conscious, voluntary control, somatic reflexes involve the involuntary contraction or relaxation of skeletal muscle.
Visceral reflexes are essential for maintaining homeostasis. Some input to the autonomic nervous system is somatosensory. For example, if an animal suddenly cuts itself, this noxious stimulus can activate the sympathetic nervous system, causing local vasoconstriction as well as increased blood pressure and heart rate. Visceral sensory information can also stimulate such reflexes. Sensory information from the thoracic and abdominal region is carried to the brain via the vagus nerve, information from the head and neck via the glossopharyngeal nerves, and visceral chemo- sensory information (i.e., taste) by the facial nerves. This sensory information is all carried to the nucleus of the solitary tract.
Spinal reflexes
Monosynaptic reflexes
Skeletal muscle is essential for the control of posture and voluntary movement. The brain sends messages to the muscles in order to initiate movement. For example, assume the brain sends a message to the leg of a horse in order to raise the leg and hold it above the ground. After the leg is raised, proprioceptive messages are constantly sent back to the brain from the leg in order to inform the brain as to the location of the leg.
Stretch reflex
The stretch reflex, also called the myotatic reflex from its Greek roots (myo = muscle + tatic = stretch), is the simplest reflex in the body, and it is an example of a proprioceptive message involved in maintaining posture and muscle tone. It is a monosynaptic reflex providing autonomic control of skeletal muscle. The sensory mechanism involves the muscle spindles, which are small, encapsulated sensory receptors located within skeletal muscle that provide information about the changes in length of the muscle.
Anatomy of the muscle spindle
There are two types of muscle fibers within the skeletal muscle: extrafusal and intrafusal. Extrafusal muscle fibers are those fibers found outside of the muscle spindle making up the bulk of skeletal muscle, and which are responsible for muscle contraction. In contrast, intrafusal (intra = within; fusal = spindle) muscle fibers are about one-quarter the size of the extrafusal fibers, are located within the connective tissue capsule that surrounds the muscle spindle, and run parallel to the extrafusal fibers (Fig. 10.17). The middle third of the capsule is swollen, giving it a spindle shape. The central region of each intrafusal fiber lacks myofilaments and is thus noncon tractile.
There are two types of intrafusal fibers: nuclear bag fibers and nuclear chain fibers. A typical muscle spindle has two to three nuclear bag fibers and a varying number of nuclear chain fibers. Each intrafusal fiber is innervated by both motor and sensory
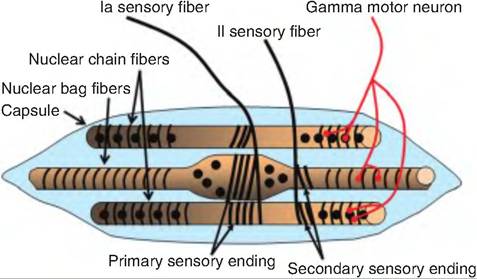
Fig. 10.17. Muscle spindle. A muscle spindle consists of three components: intrafusal fibers, sensory endings, and a motor neuron. There are two types of intrafusal fibers: nuclear chain and nuclear bag fibers. There are two types of sensory endings: primary endings and secondary endings. The intrafusal fibers are innervated by a gamma motor neuron.
neurons. There are two types of sensory fibers. The primary sensory endings are large type Ia fibers that sense both rate and amount of stretch of the muscle spindle. The secondary sensory endings are small type II fibers that surround the endings of the intrafusal fibers and sense only stretch of the muscle spindle. There are gamma motor neurons that innervate each intrafusal fiber, whereas α-motor neurons innervate the extrafusal fibers.
When a muscle is stretched, the intrafusal fibers in the muscle spindle are also stretched. This causes increased activity in the sensory endings. When the muscle shortens, the activity in the sensory endings decreases. Therefore, the components of the stretch reflex include (1) stretch of the muscle spindle, (2) activation of the sensory neurons in the muscle spindle, (3) transmission of the sensory signal to the α-motor neurons located in the dorsal horn in the spinal cord, and (4) stimulation of muscle contraction induced by the α-motor neurons (Fig. 10.18).
Stimulation of the gamma motor neuron causes the intrafusal fibers to contract, while having no effect on the extrafusal fibers. Contraction of the intrafusal fibers does not contribute to the force of muscle contraction; instead, that is generated by the extrafusal fibers. Stimulation of the gamma motor neurons causes the intrafusal fibers to contract at either end. Since they are attached to the ends of the capsule, this causes the sensory endings to increase their firing rate because they sense stretch.
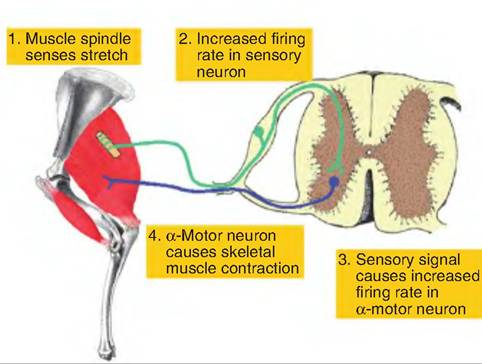
Fig. 10.18. Components of stretch reflex. 1. A muscle spindle senses stretch in the skeletal muscle caused either by contraction of the muscle or tension on the muscle. 2. The sensory endings in the muscle spindle are activated, resulting in an increased firing rate in the sensory fibers from the muscle spindle. 3. The sensory fibers synapse within the spinal cord on the α-motor neurons going to the same muscle. 4. The α-motor neuron is activated and causes the same muscle to contract. (Leg was adapted from Riegel and Hakola, 1996.)
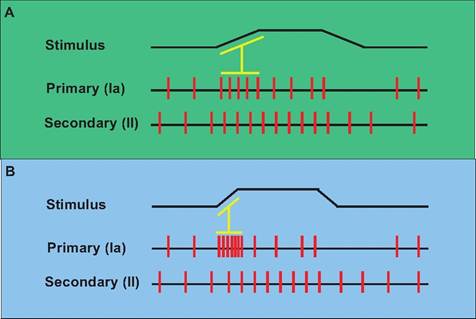
Fig. 10.19. Response of muscle spindles to stretch. (A) During stretch of the muscle, both primary and secondary endings in the muscle spindle respond and increase their firing rate. The primary endings give additional information regarding the velocity of stretch as seen here as an increase in firing rate associated with the initiation of stretch. (B) Since stretch occurred more rapidly, the primary endings showed increased frequency of firing at the beginning of the muscle stretch compared to that seen in panel A.
Muscle spindle and muscle contraction
Since contraction of intrafusal fibers does not contribute to the force of muscle contraction, what is the function of the muscle spindle? When a muscle contracts, there are two phases to contraction. The dynamic phase is the period during which the muscle length is changing; the static phase is when the muscle has stabilized its length. The two sensory endings provide information regarding these phases (Fig. 10.19). When the muscle is first stretched, both the primary and secondary endings increase their firing rate. Since the primary endings are also sensitive to the velocity of stretch, their firing rate also provides information regarding the rate of stretch (or contraction) of the muscle.
Function of the gamma motor neurons
When the α-motor neurons are stimulated, they cause contraction of extrafusal fibers within skeletal muscle. Since the muscle spindle capsule is anchored within the extrafusal fibers, the muscle spindle becomes flaccid as the two ends of the spindle move closer together (Fig. 10.20). As a result, the muscle spindle
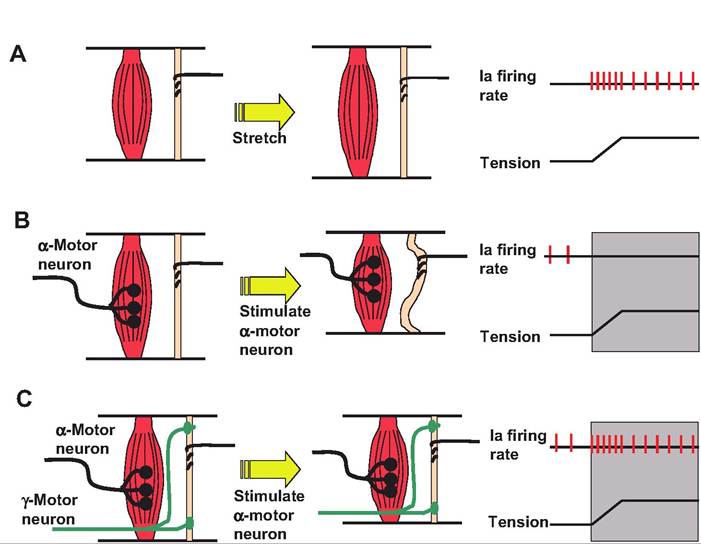
Fig. 10.20. Role of gamma motor neuron in muscle spindle. Although the muscle spindle is actually found deep in the muscle, for clarity, the extrafusal and intrafusal fibers have been displayed separately in this figure. (A) When the muscle is stretched, there is an increase in firing rate in the sensory neurons. Only the primary endings are displayed in this figure. (B) When the α-motor neuron is stimulated, it causes contraction of the extrafusal fibers. Since the muscle spindle is anchored within these fibers, the muscle spindle become flaccid as the extrafusal fibers shorten. (C) If the gamma motor neuron is stimulated simultaneously with the α-motor neuron, the intrafusal fibers contract along with the extrafusal fibers, thus preventing the muscle spindle from becoming unloaded. Therefore, the muscle spindle is able to maintain its ability to sense stretch within the muscle. (Figure modified from Kandel et al., 2000.)
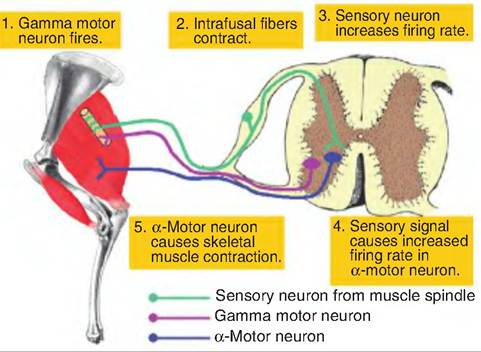
Fig. 10.21. Gamma motor neuron loop. Stimulation of the gamma motor neuron causes shortening of the intrafusal fibers within the muscle spindle. This causes an increased frequency of firing in the sensory neuron that synapses on the α-motor neuron. Stimulation of the α-motor neuron results in contraction of the skeletal muscle. (Leg was adapted from Riegel and Hakola, 1996.)
becomes insensitive to stretch. To correct this situation, the intrafusal fibers in the muscle spindle are innervated with γ-motor neurons. Stimulation of γ- motor neurons results in contraction of intrafusal fibers, thus causing the muscle spindle to shorten so that it remains responsive to changes in the length of the extrafusal fibers.
Gamma motor neuron loop
Although conscious motor movement is usually initiated by signals carried by the α-motor neuron, there is a second method to initiate the contraction of skeletal muscle called the gamma motor neuron loop (Fig. 10.21). Stimulation of a gamma motor neuron causes contraction of intrafusal fibers, which causes the muscle spindle to detect stretch. The sensory endings in the muscle spindle detect the stretch, resulting in an increased firing rate in the sensory neuron. This information is transmitted back to the CNS where the sensory neurons synapse directly on the α-motor neurons going to the same muscle. Increased firing rate in the α-motor neuron results in contraction of the muscle in order to reduce the stretch in the muscle spindle.