The Nervous System
OBJECTIVES
• describe the anatomy of a neuron and a nerve using diagrams or a prepared slide
• understand the reflex arc from the sensory afferent nerve fibers, through the spinal cord, to the efferent nerve fibers
• understand the mechanism of a nerve impulse
• locate and name the major anatomical structures of the sheep’s brain and meninges
• explain the function of each part of the brain
• locate and name the major nerves of the peripheral nervous system
• understand the transmission of an impulse across a synapse
• understand and describe the flow of cerebrospinal fluid from its origin
• name the cranial nerves and describe their functions
MATERIALS
• compound microscope
• prepared slide of a giant multipolar neuron and cross section of mammal spinal cord
• cat cadaver, triple injected (order without skin attached)
• sheep brains
• Mayo dissecting scissors
• probe
• 1 ? 2 thumb forceps or Adson tissue forceps
• #4 scalpel handle with blade
• bone cutting forceps
• rubber gloves
• percussion hammer
• live dog or cat
Introduction
The nervous system is the master integrating and coordinating unit of the body.
It is continuously monitoring sensory input from internal systems and from the external environment, then processing this information. All thoughts, actions, and perceived sensations are a reflection of the nervous system's activity.The nervous system can be divided into two parts: the central nervous system (CNS) and the peripheral nervous system (PNS). The CNS consists of the brain and spinal cord, whereas the PNS consists of the spinal nerves from the spinal cord and the cranial nerves from the brain. The PNS can also be divided functionally into the autonomic nervous system, which acts automatically, and the somatic nervous system, which involves controlled muscular action.
Nervous tissue has just two different classes of cells: neurons and their supporting cells. The supporting cells of the CNS are usually referred to as neuroglia, or glial cells. These cells include astrocytes, oligodendrocytes, microglia, and ependymal cells (Figure 15.1). They hold the neurons and their processes in place and have been described as nerve glue, which is the meaning of the word neuroglia. Supporting cells of the PNS are Schwann cells and satellite
298
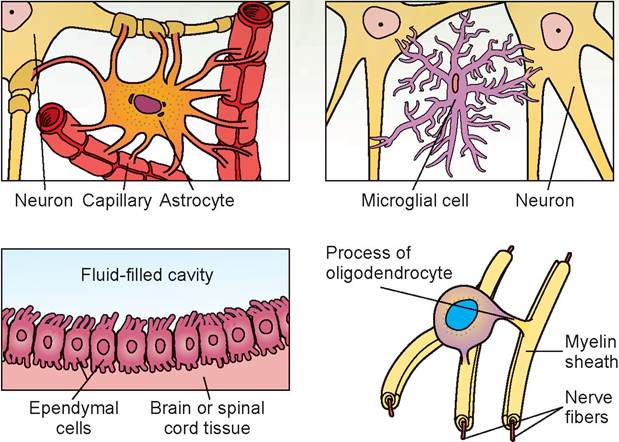
FIGURE 15.1: Supporting cells of the nervous system (all species).
cells. They serve neurons by acting as phagocytes and by bracing, protecting, and myelinating the neurons' tiny, delicate fibers.
In addition, these support cells play a role in the exchange between local capillaries and neurons to control the surrounding chemical environment. Although the neuroglia resemble the neurons (because of their fibrous cellular extensions), they cannot generate or transmit nerve impulses. Their pathological importance in veterinary medicine is that, on rare occasions, they may be the source of tumor development.
In the exercises in this chapter the important structures are listed in colored bold print. If a structure is mentioned prior to its dissection, it is italicized. Structures discussed before dissection may also be in bold print for special emphasis.
EXERCISE 15.1 THE NEURON
The neuron is the basic structural unit of nervous tissue (Figure 15.2). Neurons transmit messages as nerve impulses from one part of the body to another. Although neurons in different parts of the body differ structurally, they have a number of commonly identifiable features, which are discussed in the following text.
The cell body is where the nucleus is located, and the slender processes, or fibers, extend from it. The cell bodies can be found within the CNS or outside of it. Ganglia (singular: ganglion) are small masses of nervous tissue, containing primarily the cell bodies of neurons, that are located outside the brain and spinal cord.
Collections of nerve cell bodies within the CNS constitute the gray matter.Within the cell body of a neuron, neurofibrils can be found in the cytoplasm. These are the cytoskel- etal elements of the neuron that help with support and intracellular transport. Also visible are Nissl (chromatoρhilic) bodies, which are elaborate types of endoplasmic reticulum involved in the metabolic activities of the cell.
Extending from the cell body are the neuron’s processes, or fibers, which can take a variety of forms. Dendrites are the receptive region of the neuron, as they bear receptors for neurotransmitter substances released from adjoining axons and conduct the nerve impulse toward the cell body. Neurons have many dendrites.
Axons are another type of process. Generally axons carry impulses in one direction only: away from the cell body. However, we now know that some axons transmit impulses both to and from the cell body. Therefore, axons are now defined as nerve impulse generators and transmitters.
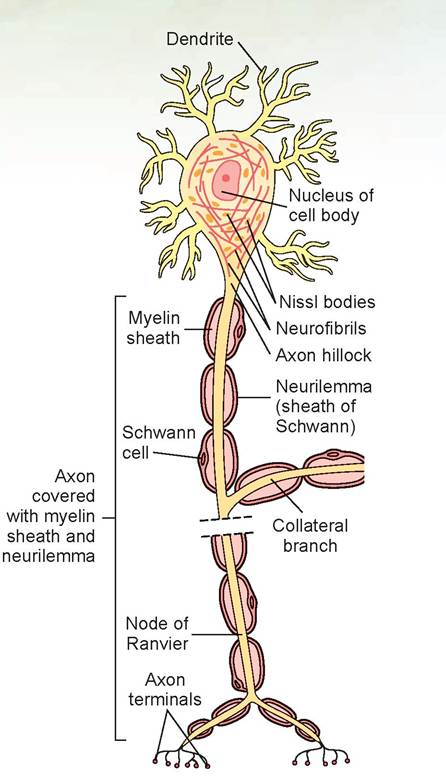
FIGURE 15.2: The structure of a neuron (all species).
Neurons are classified as unipolar, bipolar, and multipolar (Figure 15.3). Unipolar neurons are sensory neurons that originate in the embryo as bipolar neurons. During development, the axon and dendrite fuse to form a single process that divides into two branches a short distance from the cell body. Both branches function together as an axon. The branch that extends to the periphery at its distal tip has unmyelinated dendrites attached. The other branch extends into the CNS and synapses with other neurons.
Bipolar neurons have one main dendrite and one axon. They are found in the retina of the eye, in the inner ear, and in the olfactory area of the brain. Multipolar neurons usually have several dendrites and one axon. Most neurons of the brain and spinal cord are of this type. Even though multipolar neurons have only one axon, the axon may branch into collaterals.
Note that the term nerve fiber is a synonym for axon and is thus quite specific.Neurons communicate with one another or with cells of other tissues (such as muscle) by transmitting impulses from the terminal end of the axon (axon terminal) to another neuron’s dendrite (Figure 15.4) or to another type of cell. The terminal end of an axon meets a dendrite at a synapse. These axon terminals have synaptic vesicles containing the neurotransmitter substance, and numerous mitochondria for energy. The membrane of the axon terminal is called the presynaptic membrane. When the nerve impulse reaches the axon terminals, some of the vesicles release the neurotransmitter substance to diffuse across the synaptic cleft (the tiny gap between the presynaptic membrane of an axon and the postsynaptic membrane of the dendrite). This substance then stimulates the dendrite, and thus the impulse is picked up and carried to the receiving nerve’s cell body. It was long thought that each type of axon produced only one neurotransmitter, but we now know that axons may produce two or three different types of neurotransmitter substances, each with its own specific synaptic vesicle.
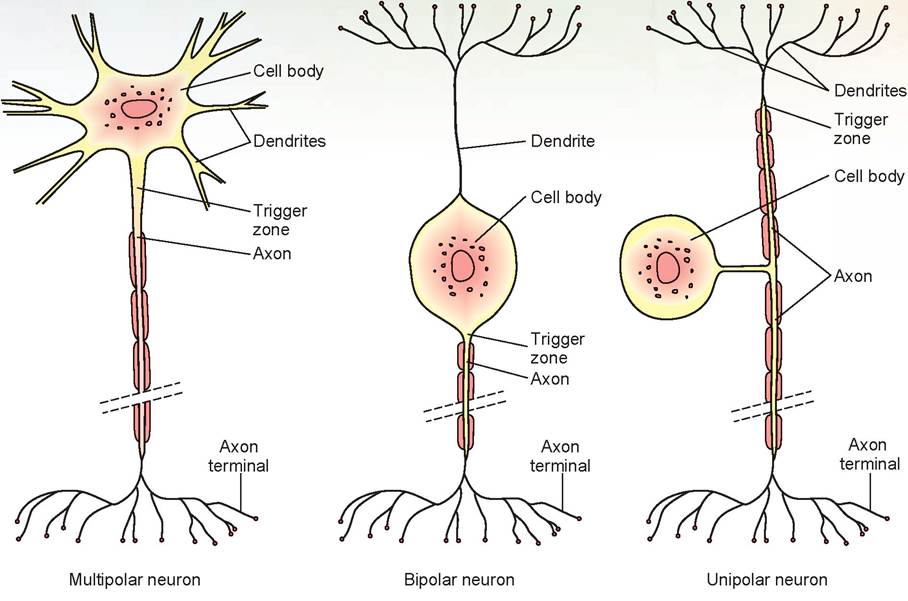
FIGURE 15.3: Structural classification of neurons.
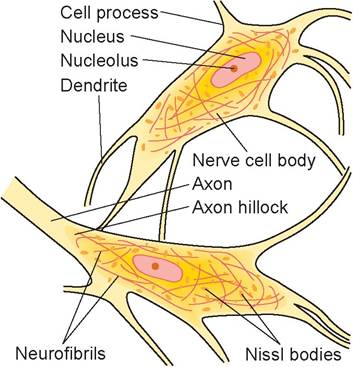
FIGURE 15.4: Neuron cell bodies and their processes.
Procedure
1. First, examine the giant multipolar neuron smear slide using the 10X objective lens. Locate a large, darkblue motor neuron. Center the neuron in your field of vision and change to the 40X objective lens.
2. Note the distinct separation between the blue and the pink background at the cell boundary. This is the plasma membrane. The details of this membrane are not visible using the light microscope. These cells have numerous cytoplasmic extensions called cell processes. Some of these processes were cut off during the sectioning of the tissue.
Each cell, if sliced perfectly down the middle, would have a long process called an axon, and numerous shorter ones called dendrites.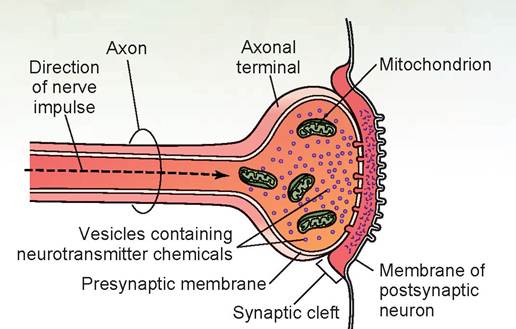
FIGURE 15.5: A synapse.
If you are using a slide stained specifically for RNA, the nucleus (with mostly DNA) has a halo appearance around the RNA-rich nucleolus (the dark dot in the center). The nuclear membrane will not be visible on an RNA slide (Figure 15.5) unless it is stained with hematoxylin and eosin stain. Observe the granular structures within the cytoplasm; these are the Nissl bodies. Now look for neurofibrils, which are linear, darkly stained structures often found at the widened area where the axon leaves the cell body (the axon hillock).
The Nerve Impulse
Neurons have two major physiological properties: They are excitable (can convert stimuli into a nerve impulse), and they are conductive (can transmit the impulse). The following characteristics differentiate a nerve impulse from an electric impulse that runs through a wire:
1. Nerve impulses are based on ion movement rather than electron movement.
2. Nerve impulses are slower than electric impulses.
3. Nerve impulses are active and selfpropagating.
4. Nerve impulses require energy in the form of ATP.
5. Nerve impulses move at a constant amplitude and velocity.
A nerve impulse is described as an action potential (Figure 15.6). The difference in charge on two sides of a cell membrane results in a voltage known as the resting membrane potential, and a neuron in this state is polarized. This resting potential, measured across the axonal cell membrane, is approximately -70 millivolts (mV). The value is negative because the inside of the nerve cell membrane is negatively charged relative to the exterior
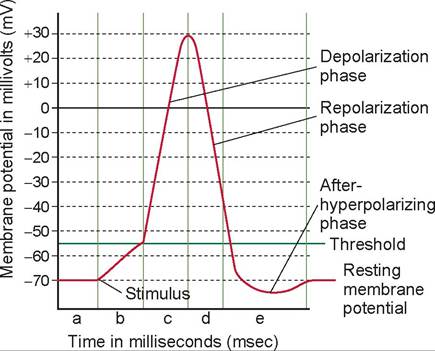
FIGURE 15.6 The action potential, or nerve impulse, and its phases.
A. During resting membrane potential the voltage-gated Na+ channels are resting and voltage-gated K+ channels are closed. B. Rise to threshold potential: Sufficient stimulus causes depolarization to the threshold potential. C. Depolarization: The voltagegated Na+ channels' activation gates are open. Part C represents the absolute refractory period. D. Repolarization: The Voltage-gated K+ channels are open; Na+ channels are inactivating. E. After hyper-polarization: voltage-gated K+ channels are still open; Na+ channels are in a resting state. Parts D and E represent the relative refractory period.because of an excess of cations in the extracellular fluid (ECF), especially sodium (Na+).
At rest, the plasma membrane is virtually impermeable to sodium. A balance is maintained (by an active sodium-potassium pump) in which there are more potassium ions (K+) inside the cell and more sodium ions (Na+) outside the cell. When an axon receives a stimulus, the plasma membrane briefly becomes more permeable, and sodium rushes into the cell, resulting in more positive ions inside the cell (the chloride ions ∣ Cl ∣ in the extracellular fluid stay outside the cell). Thus, the interior of the cell starts to become less negative, and the outside less positive. This process is called depolarization.
If the stimulus is strong enough to depolarize the membrane to a critical level, called a threshold (approximately -55 mV), voltage-gated Na+ channels (special gates that open and close to allow ions to pass) rapidly start to open. Both the electrical and chemical gradients favor inward diffusion of Na+; this initiates the depolarizing phase of the action potential. The inflow of Na+ becomes so large that the membrane potential passes 0 mV and rises to + 30 mV. Action potentials arise according to the all-or-none principle: Once depolarization reaches the threshold an action potential occurs that is always the same size in terms of amplitude. Different neurons may have different thresholds for the generation of action potentials, but the threshold in any one neuron is usually constant.
When the voltage-gated Na+ channels close, the voltage-gated K+ channels are opening. This produces the repolarizing phase of the action potential and a process called repolarization. With the slowing of Na+ influx and the acceleration of K+ outflow, the membrane changes from +30 mV to -70 mV. However, the voltage-gated K+ channels continue to allow K+ to flow out; this overshoot is called afterhyperpolarization of the action potential. As the voltage-gated K+ channels close, the membrane potential returns to -70 mV. In a typical neuron, the entire action potential lasts about 1 msec, or 0.001 seconds.
When the Na+ is rushing in, the neuron is totally insensitive to additional stimuli and is said to be in an absolute refractory period. During the period of repolarization, it is nearly insensitive to further stimuli; however, a very strong stimulus may reactivate it. This period is called the relative refractory period. The sodium-potassium pump reestablishes the ionic balance soon after the action potential is completed, and because only minute amounts of sodium and potassium ions change places, once repolarization is completed, the neuron can quickly respond again to a stimulus.
Once generated, the action potential is selfpropagating: It spreads along the entire length of the nerve fiber. It is never partially transmitted but is an all-or-none response (see previous discussion of this concept). The nerve impulse is the propagation of the action potential in which the disruption of the membrane permeability in one area of the axon causes disruption of the membrane permeability of the distal adjacent axonal area. This initiates another action potential in that axonal area, which stimulates the next axonal area, and so on down the axon. Because these axonal areas are small, propagation (and thus nerve impulse conduction) is slow under these circumstances.
As an animal develops and grows, either in utero or early in life, the nerves become myelinated. This results in faster nerve transmission and increased coordination. Animals that need to run immediately after birth to survive are born with fully myelinated nerves; others, such as kittens, do not complete myelination of motor neurons until 4 to 6 weeks of age. Figure 15.7 shows a myelin sheath around an axon. Myelin is a fatty substance within Schwann cells that segmentally envelopes the axon for its
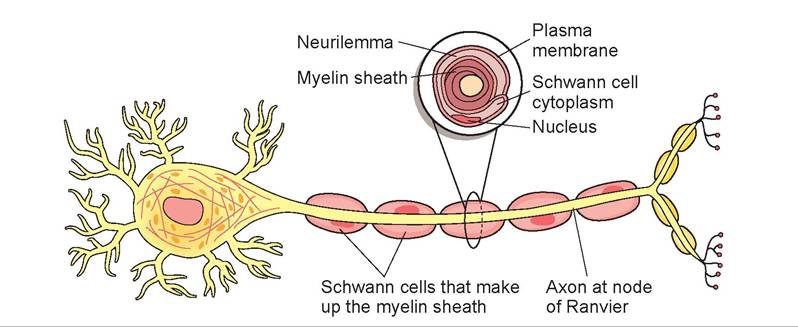
FIGURE 15.7: Axon with Schwann cells.
entire length. The areas between the Schwann cells are called nodes of Ranvier (or neurofibril nodes). In myelinated nerves, these nodes act as the adjacent axonal area in which propagation occurs.
In other words, the impulse jumps from node to node, progressing down the axon at a very fast rate, especially when compared to an unmyelinated nerve fiber.
EXERCISE 15.2 DISSECTION OF THE PERIPHERAL NERVES
There are approximately 38 pairs of spinal nerves in the cat. There are usually eight cervical, 13 thoracic, seven lumbar, three sacral, and seven coccygeal nerves. The first pair of cervical spinal nerves exits the atlas through the lateral vertebral foramina; the second pair exits between the atlas (C1) and the axis (C2). Each spinal cord segment produces a pair of spinal nerves; for example, cord segment C6 produces spinal nerve pair C6. Within the seven cervical vertebrae are eight cord segments producing eight pairs of spinal nerves. From T1 to L2 in the dog, and to L3 in the cat, the cord segment is located entirely within its associated vertebrae; in other words, inside the seventh thoracic vertebra is cord segment T7 that produces spinal nerve pair T7.
From C1 caudally, each spinal nerve emerges from the intervertebral foramen between its associated vertebra and the next one or, in other words, just caudal to its associated vertebra. Continuing with the previous example, T7 exits the vertebral column just caudal to the seventh thoracic vertebra (between the seventh and eighth thoracic vertebra). Caudal to L2, more than one cord segment can be found within the vertebral bodies (Table 15.1).
Even though the cord segments lie within a more cranial vertebra, their spinal nerves still exit caudal to the associated vertebral body. This means the nerves travel down the vertebral canal to reach the proper intervertebral foramen. Collectively, these spinal nerves come off the end of the spinal cord to form the cauda equina, so named for its resemblance to a horse’s tail (Figure 15.8).
Procedure
1. As the spinal nerves emerge from the intervertebral foramina, they divide to form a dorsal ramus and a ventral ramus. The ventral rami form the major plexuses, the brachial and lumbosacral plexuses, which supply nerves to the front legs and hind legs as well as to the skin and musculature of the ventral trunk. The dorsal rami are smaller and supply only the skin and musculature of the back.
2. The first four cervical nerves supply the lateral neck musculature. It is not necessary to locate these nerves.
TABLE 15.1: Relationship between Vertebral Bodies and Cord Segments
| Vertebral Body | Cord Segment | |
| Dog | L1 | L1 |
| L2 | L2 | |
| L3 | L3, L4* | |
| L4 | L5*, L6*+, L7+ | |
| L5 | S1++, S2+, S3+ | |
| L6 | Ca++ (Cy++), all caudal (all coccygeal) | |
| Cat | Cord segments S1, S2, and S3 are located either within L6 or at the L5, L6 junction. | |
*Patellar reflexes (includes L4, L5, L6 cord segments) + Pedal reflexes (includes L6, L7, S1 cord segments) + Perineal reflexes (includes S1, S2, S3 cord segments)
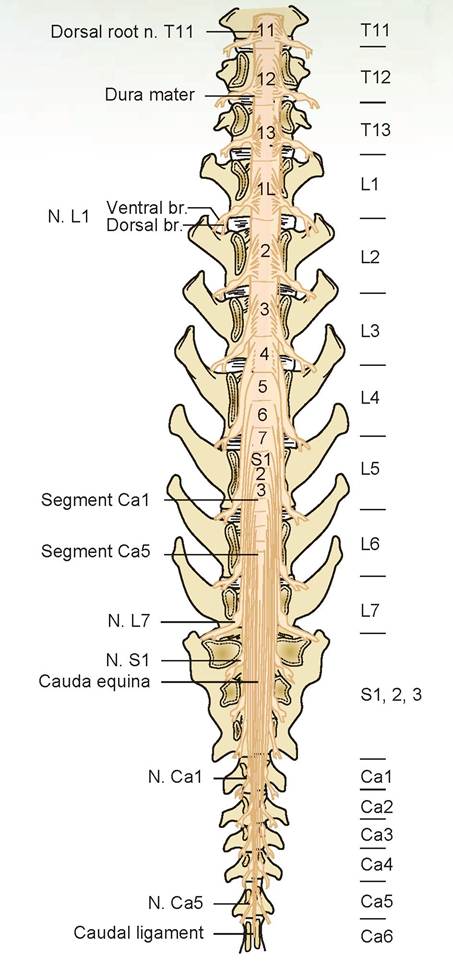
FIGURE 15.8: Spinal cord segment's placement within the spinal vertebrae and the cauda equina of the dog.
3. The phrenic nerve was identified during the dissection of the respiratory system. The two phrenic nerves are formed by the fifth and sixth cervical nerves; they pass lateral to the heart on their way to the diaphragm.
4. The sixth through eighth cervical nerves and first thoracic nerve form the brachial plexus, which supplies nerves to the muscles of the front leg. This plexus was exposed during the dissection of the chest and arm muscles. In your previous dissection, we tried to preserve the nerves on the right front leg by only dissecting the vessels on the left. Therefore, carefully cut through the pectoralis muscle group on the right side of the cat’s chest until these nerves are exposed.
5. The most cranial of the nerves of the brachial plexus that we will dissect is the musculocutaneous nerve (Figures 15.9 and 15.10). This small nerve may be identified as it passes along the medial edge of the biceps brachii muscle. This nerve supplies the biceps, coracobrachialis, and brachialis muscles.
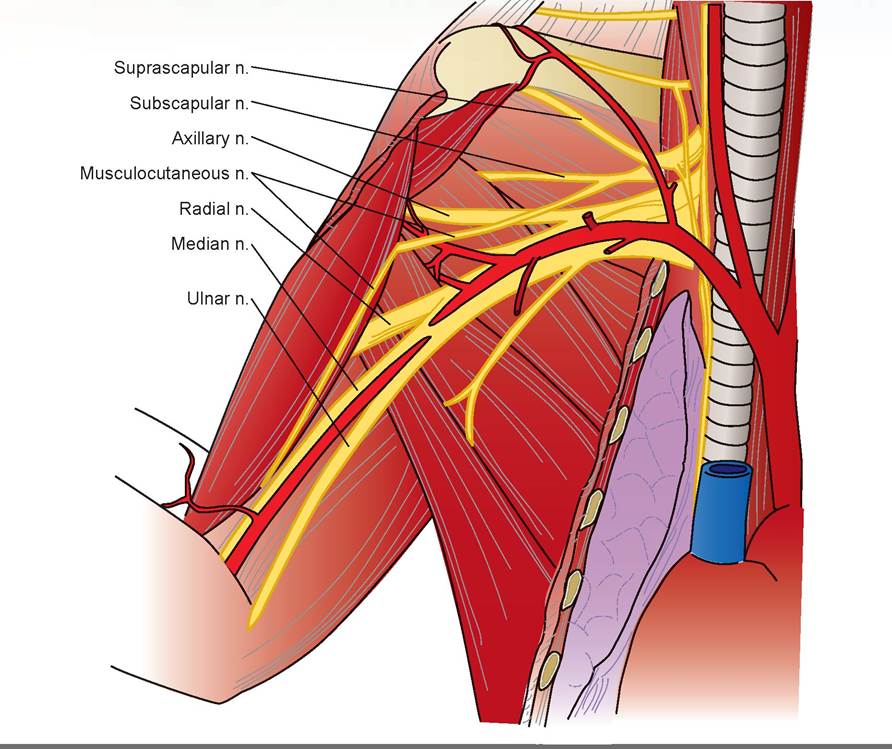
FIGURE 15.9: Diagram of the nerves of the brachial plexus of a cat.
3. The deepest of the nerves is dorsal and cranial to the brachial artery and passes between the triceps and the humerus: the radial nerve. It is the largest nerve of the plexus. Follow this nerve as it passes to the lateral surface of the upper leg and trace it distally. Near the elbow it divides into a superficial branch and a deep-muscular branch. This nerve supplies the triceps, the supinator muscles, and the extensor muscles of the lower forelimb. The superficial branch innervates the skin of the cranial foreleg.
4. Coursing adjacent and cranial to the brachial artery is the median nerve. Follow it to the elbow region, through the supracondyloid foramen in the humerus, and to the forearm. It supplies the pronators and flexors of the lower forelimb, except for the flexor carpi ulnaris.
5. The ulnar nerve is the most caudal nerve of the brachial plexus that we will dissect. It emerges from its course adjacent to the median nerve and runs caudally from the brachial artery to the carpus. It passes between the medial epicondyle of the humerus and the olecranon process of the ulna at the elbow. This nerve supples the flexor carpi ulnaris and the ulnar head of the flexor digitorum profundus.
6. The next 12 thoracic nerves pass between the ribs, each with an artery and vein. They are known as the intercostal nerves, and they supply the intercostal muscles.
7. The sciatic nerve is the major nerve emerging from the lumbosacral plexus (Figures 15.11 and 15.12). This plexus is made up of the last four lumbar nerves and the three sacral nerves. Locate the sciatic nerve beneath the biceps femoris muscle. Trace it to the popliteal fossa, where it divides into two
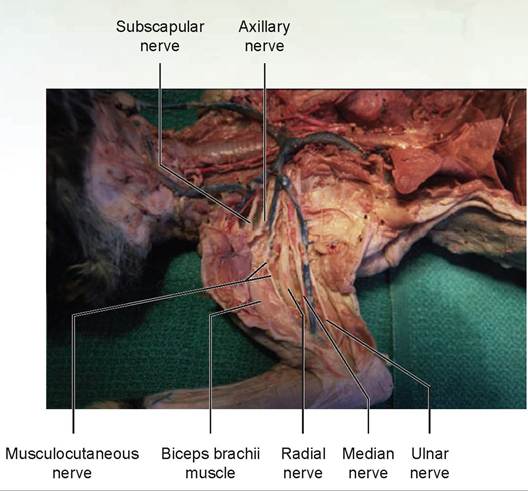
FIGURE 15.10: The nerves of the brachial plexus of a cat.
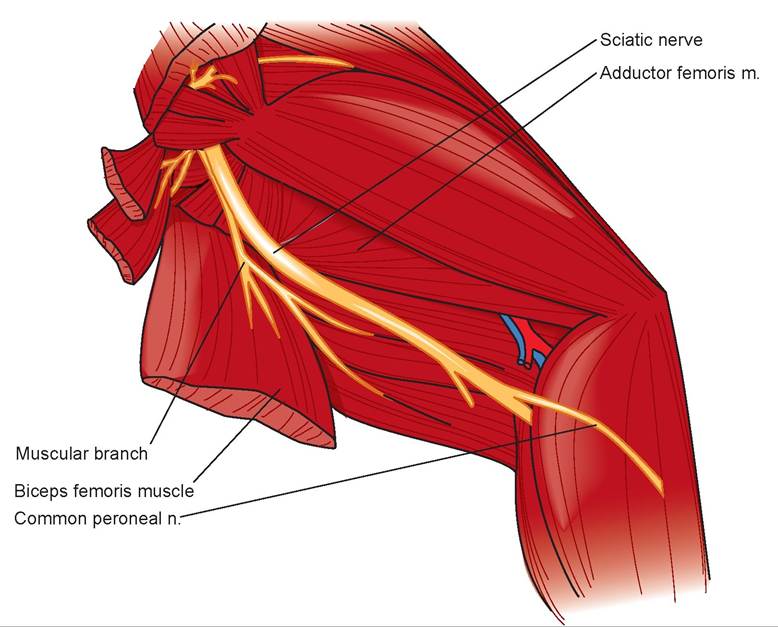
FIGURE 15.11: Diagram of the sciatic nerve of the lumbosacral plexus of a cat.
branches, which supply the leg. This nerve supplies the flexor muscles of the stifle, extensors of the hock, and the digital flexors.
11. The femoral nerve also emerges from the lumbosacral plexus. This nerve can be located on the ventral surface of the thigh, coursing with the femoral artery between the gracilis and sartorius muscles (Figures 15.13 and 15.14). It supplies the sartorius muscles and the quadriceps femoris group
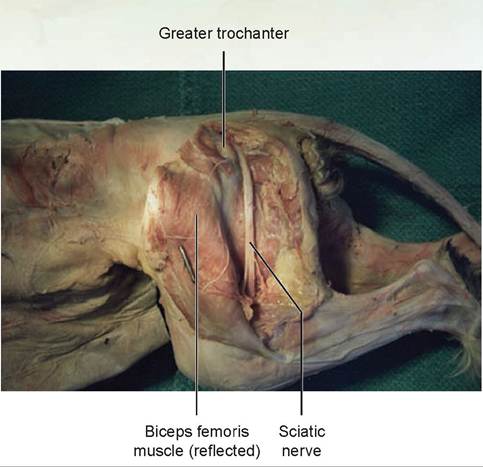
FIGURE 15.12: The sciatic nerve of a cat.
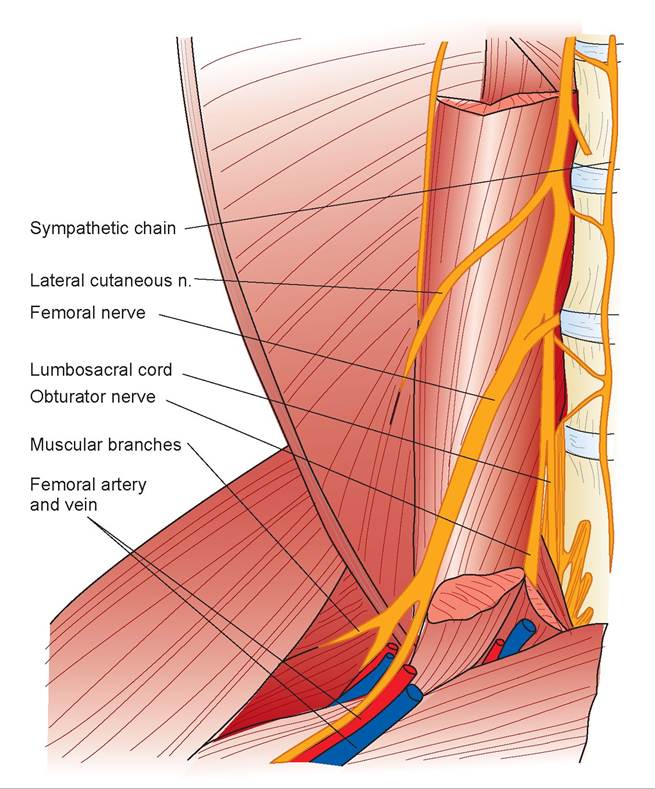
FIGURE 15.13: Diagram of the femoral and obturator nerves of a cat.
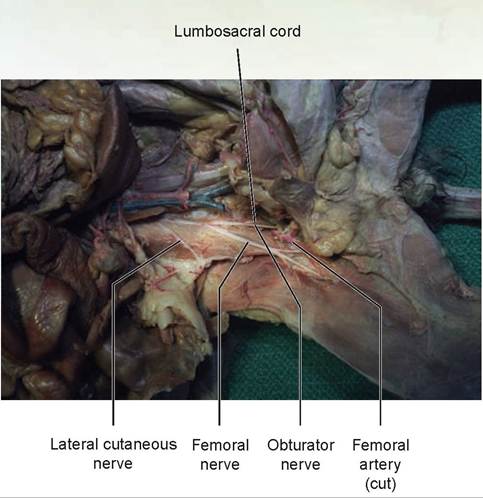
FIGURE 15.14: The femoral and obturator nerves of a cat.
(the extensors of the stifle). It continues as the saphenous nerve to supply the skin on the medial aspect of the leg. Trace the femoral nerve cranially to where it enters the ventral back muscles. Separate these muscles and continue the dissection to the femoral nerve’s origin with other lumbar nerves. Follow these nerves caudally, finding the separation between the lumbosacral cord and the obturator nerve (see Figures 15.13 and 15.14). The obturator nerve supplies the adductors, pectineus, and gracilis.
12. The autonomic nervous system of the cat will not be dissected. Without dissection, you can observe (in the thoracic cavity) the two sympathetic trunks, or chains, located on each side of the vertebral column. Push the left lung to the right and locate the sympathetic trunks on the dorsal wall of the thoracic cavity. The sympathetic trunks are beneath the parietal pleura, parallel to the vertebral column. They resemble white threads.
13. The cranial nerves also will not be dissected. The vagus nerve, or cranial nerve X (CN10), has been noted previously, as it courses adjacent to and is attached to the common carotid arteries on both sides.
The Cranial Nerves
The cranial nerves are the nerves of the peripheral nervous system that come from the brain and the brain stem. Each of these nerves is either sensory, motor, or both (if they have multiple functions). They may have autonomic function or may be voluntary. Table 15.2 lists the numbers and corresponding names of each of these nerves, its type, and its key function(s). Table 15.3 has two mnemonic devices for remembering the name and type of each nerve.
The Autonomic Nervous System
The autonomic nervous system is divided into the sympathetic and parasysmpathetic nervous systems. Figure 15.15A illustrates the neuronal arrangement of the somatic, sympathetic, and parasympathetic nerves. The parasysmpathetic nerves arise from the cranial nerves and sacral nerves. The sympathetic nerves arise from the thoracic and lumbar spinal nerves (Figure 15.15B).
The sympathetic nerves’ presynaptic fibers synapse at the sympathetic chain or other ganglia,
TABLE 15.2: Names and Functions of the 12 Cranial Nerves
| Number | Name | Type | Key Function |
| I | Olfactory | Sensory | Smell |
| II | Optic | Sensory | Vision |
| III | Oculomotor | Motor | Eye movement, pupil size, focusing lens |
| IV | Trochlear | Motor | Eye movement |
| V | Trigeminal | Both | Sensations from the head and teeth, chewing |
| VI | Abducens | Motor | Eye movement |
| VII | Facial | Both | Face and scalp movement, salivation, tears, taste |
| VIII | Vestibulocochlear | Sensory | Balance and equilibrium, hearing |
| IX | Glossopharyngeal | Both | Tongue movement, swallowing, salivation, taste |
| X | Vagus | Both | Sensory from GI tract and respiratory tree; motor to larynx, pharynx; parasympathetic motor to the abdominal and thoracic organs |
| XI | Accessory | Motor | Head movement |
| XII | Hypoglossal | Motor | Tongue movement |
TABLE 15.3: Mnemonics for Remembering Cranial Nerve Names and Types
| Number | Name | Name-Mnemonic Word | Type-Mnemonic Word |
| I | Olfactory | on | six |
| II | Optic | old | sailors |
| III | Oculomotor | Oregon's (Oklahoma's, Olympus's, etc.) | made |
| IV | Trochlear | towering | merry |
| V | Trigeminal | tops | but |
| VI | Abducens | a | my |
| VII | Facial | fine | brother |
| VIII | Vestibulocochlear | veterinary | said |
| IX | Glossopharyngeal | gastroenterologist | bad |
| X | Vagus | viewed | business |
| XI | Accessory | a | my |
| XII | Hypoglossal | horse | man |
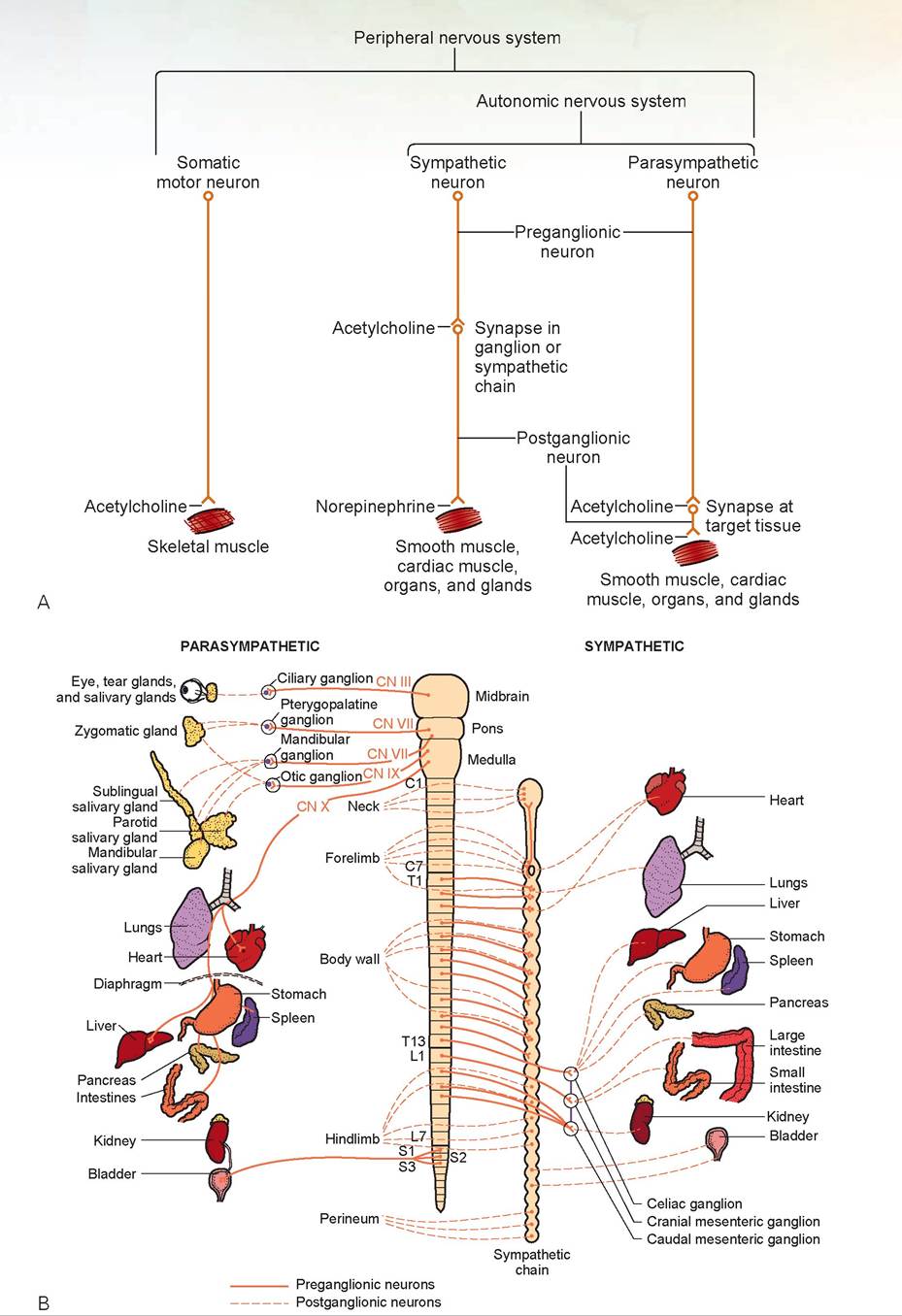
FIGURE 15.15: A. The peripheral nervous system. B. Sympathetic and parasympathetic preganglionic and postganglionic innervation.
and use acetylcholine as the neurotransmitter at this junction (see Figure 15.15A). The postsynaptic fibers innervate organs of the chest and abdomen and use norepinephrine as the neurotransmitter. The parasympathetic nerves’ presynaptic fibers are long and synapse at or just below the surface of the organ they innervate. A short postsynaptic fiber innervates the organ. Both use acetylcholine as the neurotransmitter.
The easiest way to remember the effects of the sympathetic and parasympathetic nervous systems is to think about which system is active in a given set of circumstances. Consider a cow being chased by wolves: The cow’s sympathetic nervous system is
fully turned on, and the parasympathetic turned off. The cow’s heart rate increases; the force of its heart contractions increases; the bronchioles dilate; the pupils dilate; its gastrointestinal motility shuts down; the diameter of blood vessels in the skin decreases; the diameter of vessels in muscles increases; and the diameter of vessels in the kidneys decreases.
Conversely, a cow lying down in the field, relaxed and chewing its cud, is in the exact opposite situation: The cow’s parasympathetic nervous system is active. The cow’s heart rate is decreased; the bronchioles constrict slightly; the pupils constrict slightly; and gastrointestinal blood flow, motility, and secretions
(including saliva) increase. There is no significant effect on the force of heart contractions or on the diameter of blood vessels in the skin, muscle, or kidneys. If a drug such as atropine (called a parasympatholytic), which turns off the parasympathetic nervous system, is given to an animal, the sympathetic nervous system will take over. Heart rate will increase and mucous membranes will become dry, among other effects.
Anatomy and Physiology of the Brain
The regions of the brain originate in the embryo. They are discussed in the following text.
The telencephalon is composed of the cerebrum, or cerebral hemispheres. These are divided into functional lobes, which roughly correspond to the area of the cerebrum underlying the bone of the same name.
1. The frontal lobe contains the part of the motor cortex associated with voluntary movement, as well as areas associated with psychomotor skills.
2. The parietal lobe also contains part of the motor cortex, as well as the somesthetic interpretation center, which controls conscious perception and localization of pain, touch, and temperature.
3. The occipital lobe is associated with visual interpretation.
4. The temporal lobe is associated with auditory function, behavior, and memory.
5. The piriform lobe is found on the ventral surface of the cerebrum and is associated with olfaction (Figure 15.16).
The major connection between the right and left cerebral hemispheres is the corpus callosum. Ventral to the corpus callosum and septum pellucidum is the body of the fornix. The fornix is the white matter connection between the hippocampal gyrus and nearby areas, such as the mammillary body and the brain stem.
Other areas of the brain include the diencephalon, which consists mainly of the thalamus but also
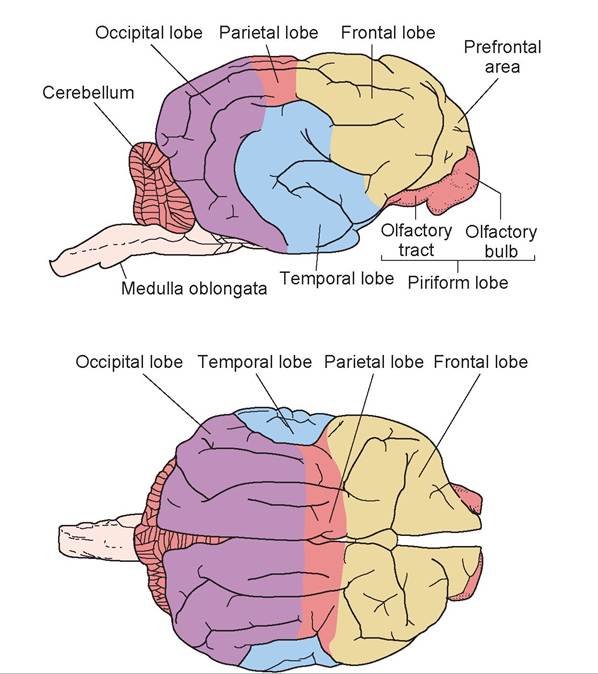
FIGURE 15.16: The lobes of the cerebrum of a sheep.
includes smaller areas of the epithalamus, subthalamus, metathalamus, and hypothalamus. The thalamus functions as a sensory relay center; it receives general sensory impulses and transmits them to the telencephalon. The other parts of the diencephalon also act as relay centers. The hypothalamus consists of the optic chiasm, tuber cinereum, mammillary body, infundibulum, and hypophysa. The functions of the hypothalamus were discussed in Chapter 13 on the endocrine system. The diencephalon and telencephalon together may be referred to as the forebrain.
The mesencephalon, or midbrain, is located immediately caudal to the diencephalon and consists of the corpora quadrigemina (made up of the rostral and caudal colliculi) and the paired cerebral peduncles. The rostral colliculi are associated with vision, and the caudal colliculi with hearing. The cerebral peduncles are essentially the continuation of the right and left halves of the spinal cord and brain stem into the respective cerebral hemispheres. They contain nerve fiber tracts and nuclei.
The embryonic rhombencephalon (hindbrain) develops into the metencephalon and myelencephalon. The metencephalon consists of the cerebellum dorsally and the pons ventrally. The cerebellum has a cortex and a medulla made up of white matter, which, on a cut surface, has a branching appearance and is called the arbor vitae. The cerebellum is connected by the rostral, middle, and caudal cerebellar peduncles to the cerebral peduncle, pons, and brain stem, respectively. The cerebellum coordinates motor activity. The pons is visible on the ventral surface, rostral to the brain stem. It is a mixture of white matter and gray matter and contains the ascending reticular activating system (ARAS). The ARAS maintains alertness or awareness by way of the cerebrum, and it controls the apneustic center and pneumot- axic center (for respiration), the nuclei of the vestibular apparatus, and the motor nucleus of cranial nerve 5 (CN V). The myelencephalon is the medulla oblongata, or brain stem. It contains the center for heart functions, as well as centers for respiration, swallowing, and vomiting.
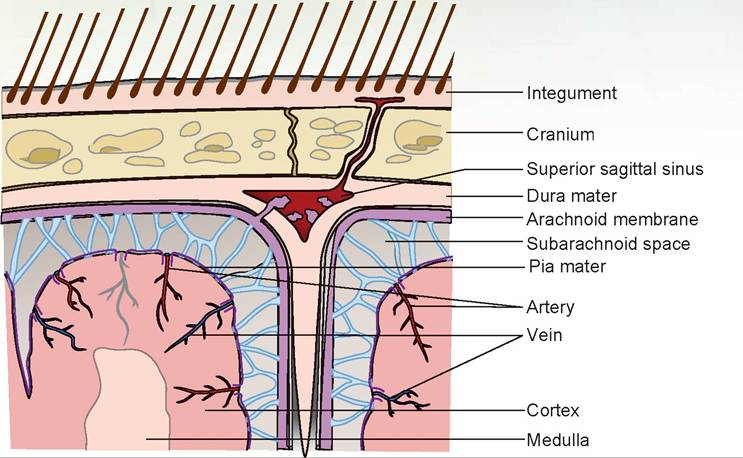
FIGURE 15.17: Diagram of the meninges of the brain (all species).
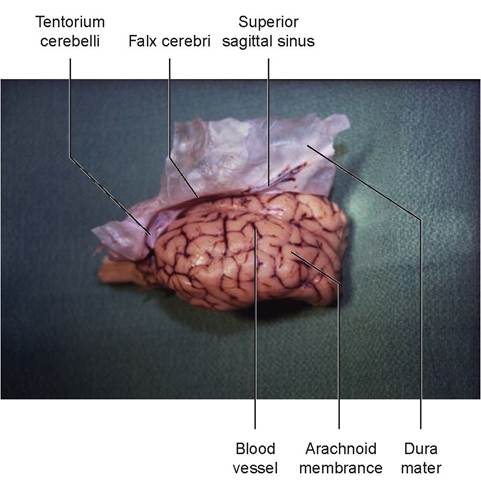
FIGURE 15.18: The meninges of the sheep's brain.
The trochlear nerve (cranial nerve [CN] IV) appears as a thin, white strand, directed ventrally and slightly caudal to the caudal colliculi.
6. Caudal to the cerebral hemispheres is the cerebellum. The cerebellum is connected to the brain stem by three prominent fiber tracts, or peduncles. Lift the cranial edge of the cerebellum and locate the rostral cerebellar peduncle, which connects the cerebellum to the cerebral peduncle of the midbrain. The middle cerebellar peduncle connects the cerebellum with the pons. Slightly caudal to this is the caudal cerebellar peduncle, connecting the cerebellum to the medulla.
7. Turn the brain over so that the ventral surface is visible. A pair of olfactory bulbs can be seen beneath the cerebral hemispheres. These bulbs lie over the cribriform plate of the ethmoid bone and receive the olfactory neurons from the nose (Figures 15.22 and 15.23).
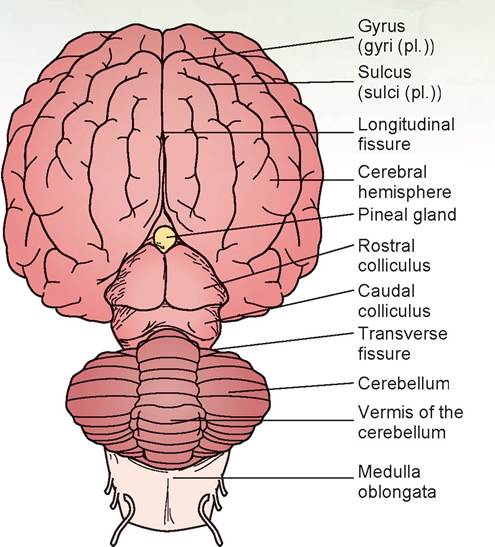
FIGURE 15.19: Diagram of the dorsal view of the sheep's brain.
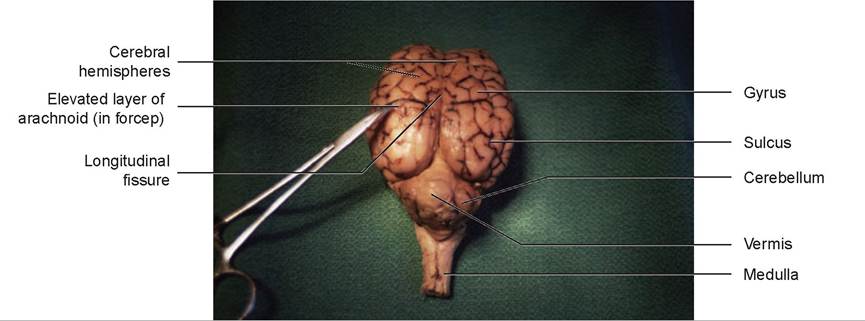
FIGURE 15.20: Dorsal view of the sheep's brain.
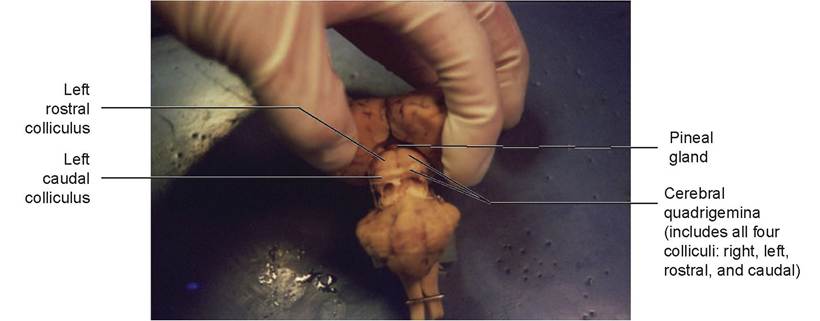
FIGURE 15.21: Corpora quadrigemina of the sheep's brain.
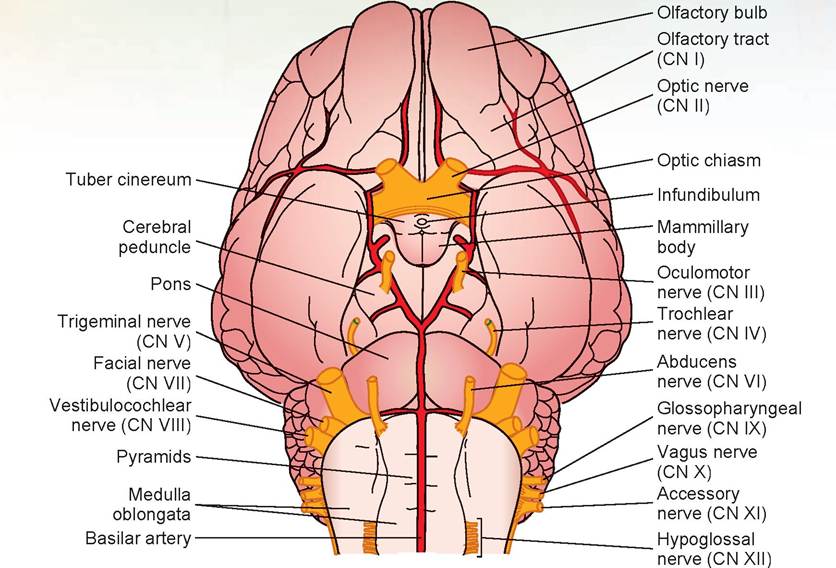
FIGURE 15.22: Diagram of the ventral view of the sheep's brain.
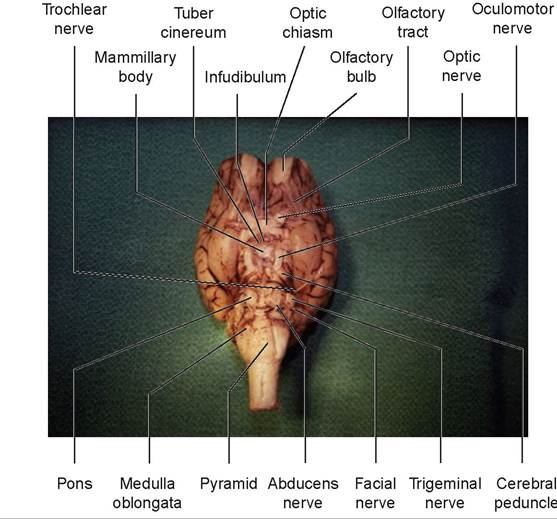
FIGURE 15.23: Ventral view of the sheep's brain.
8. A white band, the olfactory tract, extends from each olfactory bulb along the ventral surface of the cerebral hemispheres.
9. On the ventral surface of the diencephalon is the hypothalamus, located posterior to the olfactory tracts. The optic nerves (CN II) undergo a partial crossing over (decussation) to form the rostral border of the hypothalamus, known as the optic chiasm. This results in the left eye connecting to the right side of the brain, and the right eye to the left side of the brain.
10. The rest of the hypothalamus is the oval area lying caudal to the optic chiasm. The infundibulum can be seen connecting the pituitary to the hypothalamus. Caudal to the infundibulum are the paired, rounded mammillary bodies. Around the infundibulum is the tuber cinereum (see Figures 15.22 and 15.23).
11. Observe the cerebral peduncles on the ventral surface of the midbrain. The large oculomotor nerves (CN III) arise from the cerebral peduncle caudal to the mammillary body.
12. Caudal to the midbrain is the pons. This is composed primarily of white fibers, many of which run transversely across the pons and out to the cerebellum. Just caudolateral to the pons on both sides is the large trigeminal nerve (CN V), and caudolateral to this nerve are two nerves, the facial nerve (CN VII) and the vestibulocochlear nerve (CN VIII). The abducens nerve (CN VI) is located at the border between the pons and the medulla just off the midline on both sides. The rest of the nerves will be difficult to locate because they are often broken off as the dura mater is removed.
13. The medulla oblongata is caudal to the pons. The longitudinal bands of tissue on either side of the ventral median fissure on the ventral surface of the medulla are known as the pyramids (see Figures 15.22 and 15.23).
14. Cut the brain in half along the longitudinal sulcus to create a sagittal section of the sheep’s brain (Figures 15.24 and 15.25). Locate the corpus callosum, the white fibers connecting the two cerebral hemispheres.
15. The thin covering of tissue separating the left and right lateral ventricles is the septum pellucidum, which is ventral to the corpus callosum. The septum should be intact on one side of the brain, and an opening to the lateral ventricle should be visible on the opposite side (as it is virtually impossible to split the septum pellucidum down the middle). Inside the lateral ventricles is the choroid plexus, a network of capillaries protruding into each ventricle and covered by a layer of ependymal cells (derived from the lining membrane of the ventricles). This produces cerebrospinal fluid (CSF), which flows out of each lateral ventricle through the foramen of Monro, to the third ventricle through the mesencephalic aqueduct, and into the fourth ventricle. Then the CSF either flows down the central canal of the spinal cord or passes out into the subarachnoid space via the foramen of Luschka. The CSF forms a cushion to protect the brain and spinal cord.
16. The fornix, a band of white fibers, lies ventral to the septum.
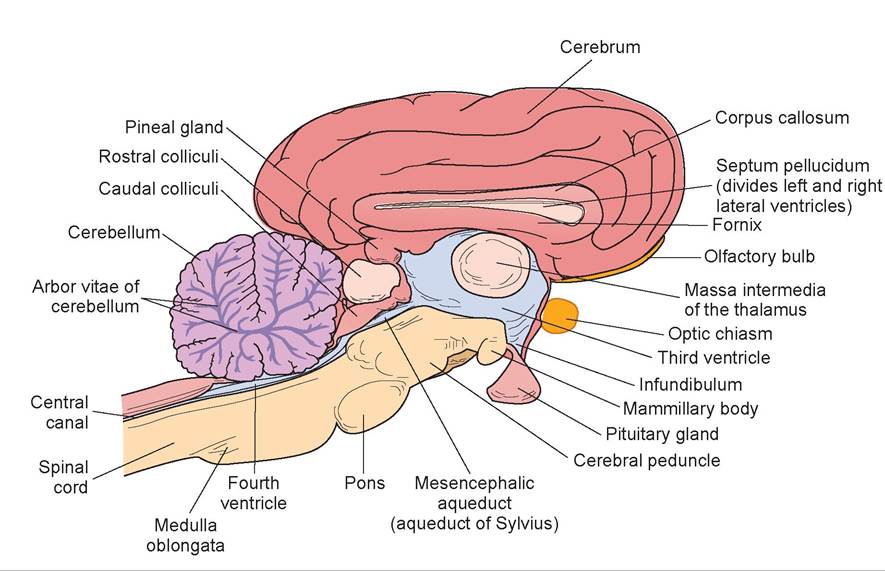
FIGURE 15.24: Diagram of the sagittal view of the sheep's brain.
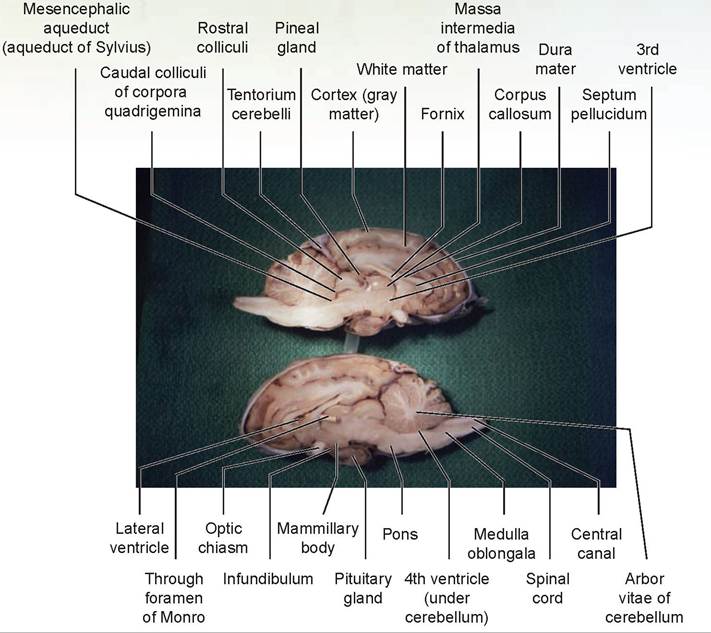
FIGURE 15.25: Sagittal view of the sheep's brain.
17. The third ventricle and thalamus lie ventral to the fornix. The narrow third ventricle, the walls of which are covered by a shiny layer of epithelium, is on the midline. The thalamus forms the lateral walls of the third ventricle.
18. The massa intermedia connects the two sides of the thalamus. The third ventricle forms a partial circle around the massa intermedia. This structure appears as a dull, circular area not covered by epithelium.
19. The foramen of Monro, the opening through which each lateral ventricle communicates with the third ventricle, lies in the depression rostral to the massa intermedia. Find this connection by passing a dull probe through it.
20. Relocate the hypothalamus ventral to the third ventricle. Also note the pineal body dorsal to the midbrain, near the rostral colliculi. Observe the narrow mesencephalic aqueduct (aqueduct of Sylvius) leading through the midbrain and connecting the third and fourth ventricles.
21. The fourth ventricle lies above the pons and medulla and below the cerebellum.
22. The beginning of the spinal cord may be seen connected to the medulla. A canal known as the central canal, which is connected to the fourth ventricle, is present in the center of the cord.
23. Note the tree-like arrangement of gray and white matter in the cerebellum. This arrangement is known as the arbor vitae (“tree of life”). The gray matter of the cerebellum is the cortex, and the white is the medulla.
24. Make a cut into the cerebral hemisphere. The outer layer of each cerebral hemisphere is its cortex, composed of gray matter. The white area beneath the cortex is the white matter.
EXERCISE 15.4 THE CROSS SECTION OF THE SPINAL CORD
The spinal cord is part of the central nervous system and starts at the foramen magnum of the skull and lies within the vertebral canal of the vertebrae.
1. Obtain a slide of a cross-sectional view of mammal spinal cord (Figure 15.26).
2. The spinal cord is divided into two mirror halves, right and left. The two halves are divided dorsally by a sulcus, the dorsal median sulcus, and ventrally by a fissue, the ventral median fissure. The gray matter is located centrally on both sides, surrounded by white matter peripherally. The gray matter contains nerve cell bodies, both in the brain and in the spinal cord.
3. The white matter contains the white tracts, or columns, that communicate with the brain stem, cerebellum, and cerebrum. It is white because these nerve fibers are myelinated. The dorsal white column is called the dorsal funiculus; the lateral column, the lateral funiculus; and ventral column, the ventral funiculus.
4. In the center is the central canal, which communicates with the fourth ventricle.
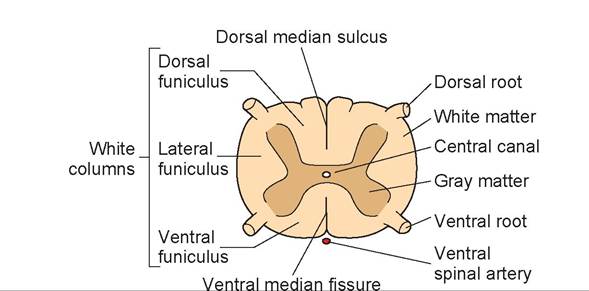
FIGURE 15.26: The cross section of the spinal cord (any species).
EXERCISE 15.5 SPINAL REFLEXES
A spinal reflex arc is a pathway of a nerve impulse. A sensory afferent neuron extends to the spinal cord, synapses there with an interneuron (also called an internuncial neuron or Renshaw cell), and synapses again with a motor efferent neuron (Figure 15.27).
The neurotransmitter at the efferent motor neuron is excitatory. It establishes an excitatory postsynaptic potential in the efferent motor neuron that goes to the agonist muscle. This neurotransmitter is acetylcholine. Simultaneously, another interneuron establishes an inhibitory postsynaptic potential to the efferent motor neuron of the antagonist muscle by releasing an inhibitory neurotransmitter, glycine, at the spinal cord level. Gamma-amino butyric acid (GABA) is the inhibitory neurotransmitter in the brain. As a result, the agonist muscle is contracting while its opposing muscle, the antagonist, is relaxing, thus allowing easy movement of a limb or other body part. The excitatory neurotransmitter is removed from the synaptic cleft by the release of an enzyme—in this case, acetylcholinesterase.
Procedure
Using a live dog or cat, observe the following reflexes. They are used to gauge the depth of anesthesia.
1. palpebral (blink) reflex: Touch the hairs on the lateral side of the eyelid or the medial canthus of the eye; the response should be a blink.
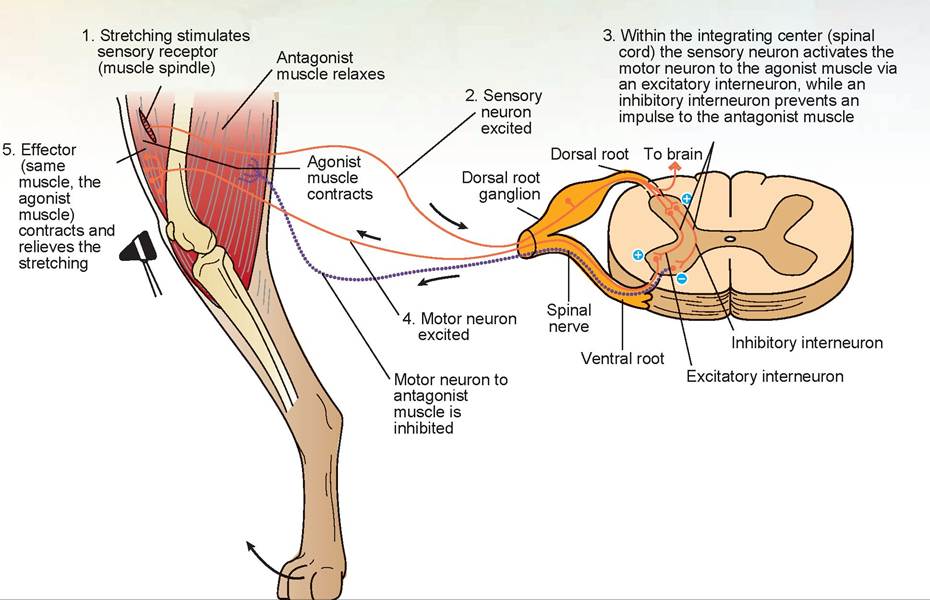
FIGURE 15.27: The spinal reflex arc in a dog.
2. corneal reflex: Touch the cornea with a wisp of cotton from a cotton ball; the response should be a blink.
3. oral-pharyngeal reflex: Try to open the animal’s mouth or place an object in the mouth while touching the pharyngeal region. The normal response should be for the animal to close its mouth. This reflex is often stimulated when pilling an animal.
4. laryngeal reflex: Touch a part of the larynx, such as the epiglottis, with a wooden tongue depressor. The normal response should be for the animal to cough or swallow; the epiglottis should close and so should the vocal folds.
5. ear pinna reflex: Lightly touch the lateral aspect of the upper part of the ear; the normal response is for the ear to twitch.
6. pedal reflex (toe pinch): Extend the rear limb while simultaneously pinching the middle toe; the normal response is a reflexive withdrawing of the limb.
7. patellar reflex: Strike the straight patellar tendon lightly with a percussion hammer; the normal response is reflexive jerking of the lower leg.
8. flexors of the tarsus and extensors of the digit: Place your index finger on the cranial tibial muscles and strike your finger semi-softly with the percussion hammer. The response should be flexion of the tarsus and extension of the digits.
9. extension of the tarsus: Grasp behind the stifle with your thumb, and strike it with the percussion hammer. The response should be for the animal to extend the tarsus.
10. extension of the elbow: Place your finger on the tendon of the long head of the triceps and strike your finger with the percussion hammer. The animal should extend the elbow joint.
Questions
1. Why is it necessary to strike your finger or thumb placed over the muscle instead of the muscle directly?
2. Why is the patellar reflex not abolished during general anesthesia?
Discussion
With the patellar reflex, the percussion hammer strikes a relatively tough tendon, whereas at the other sites it would strike the muscle directly and could damage it. Plus, by striking your finger or thumb you spread the stimulus across the entire muscle belly.
The patellar reflex, being an entirely spinal reflex, is not abolished during general anesthesia because the anesthetic is depressing the brain but not the spinal cord.
Clinical Significance
The nervous system is so intricate and delicately balanced that even minor problems can have catastrophic effects. Take, for instance, the disease hydrocephalus. The condition may be congenital or acquired. The congenital form occurs in small and brachycephalic breeds, such as the bulldog, Chihuahua, Maltese, Pomeranian, toy poodle, Yorkshire terrier, Llasa apso, Cairn terrier, Boston terrier, pug, Pekingese, and the Siamese cat. It usually appears a few weeks after birth but may present at up to one year of age. Acquired hydrocephalus can occur in any breed at any age.
There are two types of hydrocephalus. The noncommunicating type is caused by an obstruction of CSF flow within the ventricular system of the brain. The communicating type is due to a lack of resorption of CSF into the subarachnoid space. Congenital hydrocephalus is usually obstructive, caused by a congenitally narrow cerebral aqueduct. Acquired hydrocephalus usually results from trauma, Inflammatory conditions, or neoplasia. The result of both types is an increased pressure within the lateral ventricles, which hydraulically compresses the cortex of the brain. This pressure may result in seizures, altered mental status, visual deficits, and an uncoordinated gait.
A set of diseases classified as peripheral neuropathies results in demyelination of the spinal cord and peripheral nerves. This condition may occur in many species and may be a primary condition or secondary to axonal degeneration. It most often affects the cervical spine and spinal nerves, causing progressive ataxia and spastic incoordination in the rear legs (paraparesis), which may progress to include the front legs (tetraparesis). The cause is unknown.
A host of viral and bacterial infections can cause inflammation of the brain and spinal cord. Canine distemper and rabies are two examples. The way the nervous system is structured tends to keep unwanted molecules out. The pia mater follows the vessels into the brain, enveloping them and allowing only certain-sized molecules to pass through. This is known as the pial barrier, or blood-brain barrier. It is also the reason why certain antibiotics will not enter nervous tissue. In inflammatory conditions, however, the pores in the pia mater may become bigger, allowing unwanted molecules in, but also allowing us to use certain antibiotics we could not use under normal circumstances.

λ remember being told in veterinary school that boxers as a breed are considered walking [J tumor factories. In my practice, however, this was not the case. My boxers were not getting tumors! It was magic, and I was the magician.
It was a nice delusion while it lasted. Then Tobey arrived at my practice's door. I wish I could have said “Never more!” but this dog just kept coming back. He made up for all the tumors other boxers didn't get. Fortunately, his tumors were not malignant. Most were skin tumors, but he seemed to have one of every type. Every veterinarian keeps samples of unusual tumors and growths, but Tobey had his own shelf in my cabinet. If he lived to be 15, I figured he would have his own wing in Ripley's Believe It or Not!
It was on a fine, sunny day in June that Anthony Tuscalini carried Tobey in and set him on my exam table. Tobey's hind legs were paralyzed, and he was without a deep pain response. This was not a good sign. Testing for deep pain involves pinching the toe so hard that it is painfully stimulated. If the dog withdraws its leg without evidence of feeling pain, the peripheral nerves and spinal centers are normal, but the spinal cord is not. If the dog shows evidence of feeling pain by turning its head and crying, the cord and thalamus are normal as well.
As I walked to the front office area to get another page for Tobey's medical record (I was already on page 23), I took a quick look outside. The sky had suddenly grayed, and I heard distant thunder. It was an ominous sign. I took Tobey to radiology and took a lumbar spine series of radiographs. The spinous process of the L3 vertebra had a moth-eaten appearance, which corresponded to the exact location where Tobey's panniculus reflex had stopped. This reflex comes from the dorsal rami of the spinal nerves to move the muscles of the skin. This was definitely not a good sign. After viewing the radiographs, I reexamined Tobey and was able to palpate two hard lumps under the skin on either side of the L3 vertebra.
“Tony, I think Tobey has a tumor around his backbone and spinal cord,” I told him.
“Can you do surgery to repair it?” he asked.
“You say he was moving his legs last night?” I asked skeptically.
“He wasn't walking very well, but he was moving,” he replied.
This may have been the best news yet, because that meant the spinal cord had been functional a short time ago and was still viable. After a discussion about referral to a neurosurgeon, Mr. Tuscalini asked if I could do the surgery on Tobey. “OK, I can try to do surgery, but it's a long shot, and I can't make any promises.”
“Do it!” Tony said. “We've got to give him a chance.”
About an hour later, after clipping the hair off of Tobey's back, I could see the tops of two golf ball-sized lumps buried deep under the skin on either side of the vertebral column. I made one long incision above the spinous processes of vertebral bodies T13 through L5. There were two semi-fibrous masses, one on either side of the vertebra, with a connecting stalk that passed directly through the spinous process of the L3 vertebra. I excised the two masses, which left massive holes in the subcutaneous fat and muscle tissue. The ventral part of each mass was necrotic and gelatinous rather than fibrous, and getting it out was difficult.
At the time, I felt my effort to remove every neoplastic cell had not been successful. With my 3-mm bone rongeurs, I removed bits and pieces of the dorsal lamina and spinous process. This surgery is called a dorsal laminectomy. Normally, in a decompressive surgery on the spinal cord I would do a hemilaminectory, opening only one side of the lamina to view the spinal cord. In this case, I had no choice but to remove the entire lamina and much of the dorsal spinous process. I tried to leave as much of the pedicle of the vertebra on either side of the spinal cord as possible. It was necessary to remove both articular processes of the L3-L4 junction and the left cranial articular process of the L2-L3 junction. This exposed the spinal cord, which had a significant dent in it where the tumor's stalk had passed. I knew I couldn't leave the vertebra without support; otherwise, it would tear apart at the L3-L4 intervertebral disc.
I had read about this technique in a veterinary journal and decided to use a thin, intramedullary pin and bend it on both ends around the spinous processes of L1 and L5.Then, I wired it to each vertebra, L1 to L5; there was just enough of the L3 spinous process left to attach the pin to it Had I had a spinal plate I might have used it, but attaching it to what remained of L3 would have been difficult. As it was, this worked fine.
I put Tobey on post-operative steroids and antibiotics, and five days later he walked out of the hospital. The pathology report said the tumor was a very hot fibrosarcoma (a malignant tumor of fibrous origin). I suggested that Tony consult a veterinary oncologist, but he decided against it. I thought for sure the tumor would return, but it never did. And the amazing thing was, Tobey never developed any more tumors. My only explanation was that we turned his immune system on. I know I didn't get every tumor cell, so his body must have destroyed them. Either that, or... maybe I am a magician!
Summary
Much of this chapter was devoted to explaining the differences between the central nervous system and the peripheral nervous system. You learned about the neurons and the supporting cells of nervous tissue. The structure of neurons was discussed, and you learned to classify neurons based on number of axons. You once again looked at a giant multipolar neuron, but this time with greater knowledge. You performed detailed dissections of the cat's periphral nervous system and the sheep's brain. You learned the names, types, and functions of the cranial nerves. Finally, using a live dog you learned how to test the various reflexes used to assess neurological function.
REVIEW QUESTIONS
1. Name the two parts of the nervous system and the structures of each part.
2. The peripheral nervous system can also be divided into what two parts?
3. Name the two classes of cells that exist in the nervous system.
4. What is a ganglion?
5. What are the cytoskeletal elements of the neuron cell body?
6. What is the name for the special endoplasmic reticulum found in the neuron cell body?
7. Differentiate between an axon and a dendrite.
8. Name the three classifications of neurons.
9. What is a synapse, and where may synapses occur?
10. What causes a postsynaptic potential to occur during impulse transmission?
11. What is the energy source at the axon terminal?
12. In what ways is a nerve impulse different from an electrical impulse?
13. What ion moves into a neuron’s axon to create depolarization?
14. What ion moves out of a neuron’s axon during repolarization?
15. How do the ions in Questions 14 and 15 reestablish the ionic balance that existed prior to the action potential?
16. Differentiate between the absolute refractory and relative refractory periods.
17. What does “self-propagating” mean?
18. What is the fatty substance of a Schwann cell?
19. What is the area between adjacent Schwann cells called?
20. Indicate the number of pairs of spinal nerves present in each part of the vertebral column.
21. What does the phrenic nerve innervate?
22. Name the two parts of the autonomic nervous system.
23. Name the 12 cranial nerves, and indicate the key function(s) of each.
24. What is the postganglionic excitatory neurotransmitter in the sympathetic nervous system?
25. What is the excitatory neurotransmitter for all other synapses and at the target organs?
26. Name the lobes of the brain.
27. What structure connects the right cerebral hemisphere to the left cerebral hemisphere?
28. In what part of the brain is the hypothalamus located?
29. What is the anatomical name of the brain stem?
30. Name the layers of the meninges.
31. Which of the meningeal layers is responsible for the blood-brain barrier?
32. Define sulci and gyri.
33. Which eye connects to the left side of the brain?
34. Describe the flow of cerebrospinal fluid from the lateral ventricles to the central canal of the spinal cord.
35. Describe the difference between white matter and gray matter.
36. At what location within the gray matter of the spinal cord does the impulse from a sensory afferent neuron establish either an excitatory or an inhibitory postsynaptic potential?
37. Name the two inhibitory neurotransmitters, and indicate where each would be found.
38. How is the excitatory neurotransmitter removed from the synaptic cleft?
39. Which spinal cord segments affect the patellar reflex? (Hint: See Table 15.1.)
40. What is the name for the collection of spinal nerves that extend from the end of the spinal cord as they travel to their associated intervertebral foramina?