Complex cycles may result from stagespecific selection pressures
Organisms with complex life cycles have multiple life stages, each adapted to its habitat and habits. This flexibility may be one of the reasons that complex life cycles are so common in so many groups of organisms.
Because separate life history stages can evolve independently in response to size- and habitat-specific selection pressures, complex life cycles can minimize the drawbacks of small, vulnerable early stages.Larval Function and Adaptation
Functional specialization of particular life stages is a defining feature of complex life cycles. Having multiple stages with largely independent morphological features can result in a pairing of particular functions with particular stages. Such a pairing can reduce some of the trade-offs that result from simultaneously optimizing multiple functions.
An example of this type of specialization occurs in many insects with complex life cycles. Such insects spend their entire larval stage in a very small area— sometimes on a single plant. Insect larvae such as caterpillars and grubs are specialized eating and growing machines. They spend almost all of their time taking in food and turning it into body mass, without forming many complex morphological structures other than mandibles (chewing mouthparts or “jaws”).
Once they have accumulated sufficient mass, these larvae metamorphose into adult butterflies, moths, and beetles, whose main function is often to disperse, find a mate, and reproduce. In extreme cases, such as mayflies, the adults are incapable of feeding and live only the few hours or days it takes them to reproduce.
Marine invertebrate larvae are also specialized for feeding, although they perform this function while dispersing on ocean currents. For example, the larvae of many mollusks (such as snails and clams) and echinoderms (such as sea urchins and sea stars) have intricate feeding structures that cover most of the larval body.
These structures, called ciliated bands, are ridges covered in cilia that beat in coordinated patterns to catch tiny food particles and move them, like a conveyer belt, toward the mouth. The ciliated bands wind and fold their way around the larval bodies, many of which have extra lobes or arms that support and elongate the ciliated bands. In sea urchins, the longer the larval arms, and the longer the ciliated band, the more efficiently the larvae are able to feed (Hart and Strathmann 1994).Other specialized larval structures can help to protect small life cycle stages from being eaten by other organisms. Examples include the toxin-bearing spines of some caterpillars, the head spines of crab larvae (FIGURE 7.19), and the setae or bristles of polychaete worm larvae, which deter some predators by making the larva a large and uncomfortable mouthful.
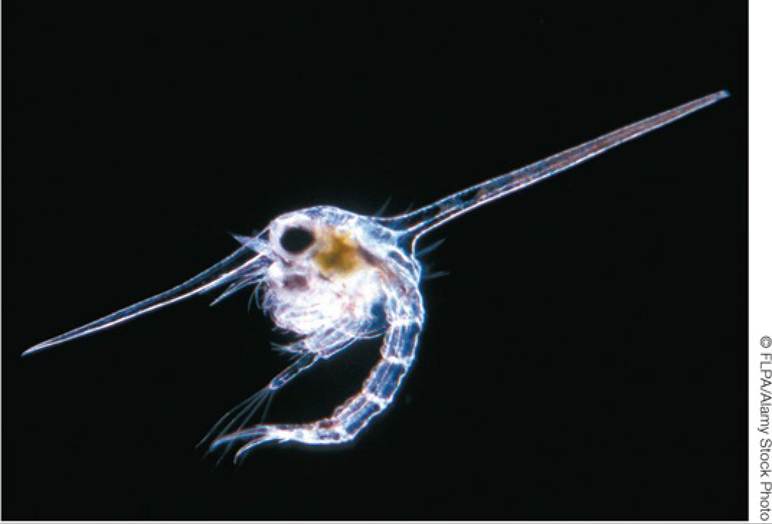
FIGURE 7.19 Specialized Defensive Structures in Marine Invertebrate Larvae The planktonic (floating) larvae of the sand crab Corystes Cassivelaunus have defensive head spines that can make them difficult for fish to eat. View larger image
Timing of Life Cycle Shifts
Most organisms with complex life cycles use different habitats and food resources at different life stages. Such shifts can occur abruptly, as in organisms that undergo metamorphosis, but they can also occur more gradually. Regardless of the speed with which changes in habitat and food preferences occur, differentsized and different-aged individuals of the same species may have very different ecological roles. We'll use the term niche shift to refer to such size- or agespecific changes in an organism's ecological function or habitat. (As we'll see in Concept 9.5, an organism's ecological niche consists of the habitat and resource requirements that the organism needs to grow, survive, and reproduce.)
In species in which an abrupt metamorphosis occurs at the transition between life cycle stages, the organism spends relatively little time in vulnerable stages that are intermediate between larva and adult.
In theory, there should be an optimal time to undergo metamorphosis, or any niche shift, that maximizes survival over the course of the life cycle. Thus, we might expect a niche shift to occur when the organism reaches a size at which conditions are more favorable for its survival or growth in the adult habitat than in the larval habitat.Dahlgren and Eggleston (2000) tested this idea for the Nassau grouper (Epinephelus striatus), an endangered coral reef fish that spends its juvenile stages in and around large clumps of algae. Smaller juveniles spend their time hiding within the algae, whereas larger ones spend their time in rocky habitats near algal clumps. By tethering and enclosing juvenile fish of different sizes in the two habitats, Dahlgren and Eggleston were able to measure mortality and growth rates in each habitat. They found that the smaller juveniles were very vulnerable to predation in the rocky habitats, while the larger juveniles were less vulnerable and were able to grow faster in the rocky habitats. Thus, the niche shift in this species appears to be timed to maximize growth and survival, as predicted.
In some cases, the larval habitat may be so favorable for growth and survival that metamorphosis is delayed—or even eliminated altogether. For example, most salamanders have aquatic larvae that metamorphose into terrestrial adults, but some salamanders, such as the mole salamander Ambystoma talpoideum, can become sexually mature while retaining gills and remaining in the aquatic habitat (FIGURE 7.20). These aquatic, gilled adults are referred to as paedomorphic, which means that they result from a delay of some developmental events (loss of gills, development of lungs) relative to sexual maturation. Both aquatic paedomorphic adults and terrestrial metamorphic adults can exist in the same population of mole salamanders. The frequency of paedomorphosis in these mixed populations seems to depend on factors such as predation, food availability, and competition—all of which influence survival and growth in the aquatic habitat.

FIGURE 7.20 Paedomorphosis in Salamanders The mole salamander Ambystoma talpoideum can produce both (A) paedomorphic aquatic adults and (B) terrestrial metamorphic adults. View larger image