Does complexity enhance stability in food webs?
Ecologists have pondered whether more complex food webs—those with more species and more links among them—are more stable than simpler food webs with lower diversity and fewer links.
Stability, in this context, is usually evaluated by the magnitude of changes in the population sizes of the organisms in the food web over time. Stability may also be expressed through ecosystem processes such as primary production. As we saw in Chapter 11, large oscillations in population size over time increase the susceptibility of species to local extinction. Thus, a less stable food web means a greater potential for extinction of its component species. The question of stability is taking on ever greater importance with increasing rates of biodiversity loss and non-native species invasions worldwide, which have significant implications for ecosystem function.Early proponents of the idea that food web complexity increases stability based their arguments on observations of real trophic interactions as well as on intuition. Ecologists such as Charles Elton and Eugene Odum argued that simpler, less diverse food webs should be more easily perturbed, experience larger changes in species population densities, and experience greater species losses as a result. More rigorous mathematical analyses of food webs, however, provided a contrary view. Robert May (1973) used food webs made up of random assemblages of organisms to demonstrate that food webs with higher diversity are less stable than those with lower diversity. The instability in May's models resulted from accentuation of population fluctuations by strong trophic interactions: the more interacting species there were, the more likely that their population fluctuations would reinforce one another, leading to the extinction of one or more of the species.
May's work overturned the notion that more complex systems are inherently more stable than simpler ones.
Yet anyone visiting a tropical rainforest or a coral reef can attest to the fact that highly diverse, productive, and complex communities do persist in nature. Therefore, much ecological research has been devoted to discovering the factors that allow naturally complex food webs to be stable. More recent models, for example, have incorporated distributions of interaction strengths more closely resembling those observed in nature. These models and experiments suggest that, while more complex systems are not necessarily more stable, some natural food webs may have a particular structure or organization that allows increased species diversity to have a stabilizing effect. Other studies suggest that the buffering influence of weak interactions (McCann et al. 1998; Neutel et al. 2002) and of behavioral or evolutionary changes in prey choice (Kondoh 2003) can help to reduce the population fluctuations associated with complex food webs. Additionally, the identity of the species in a food web is important to its behavior, with some species exerting a disproportionally greater influence on stability, and others being more likely to go extinct (Lawler 1993).How diversity at one trophic level affects the stability of populations at other trophic levels has also been of interest to ecologists, particularly in the context of biodiversity loss (as we will see in Concept 23.3). Elton (1958) proposed that plant diversity influences diversity at higher trophic levels, with greater plant diversity stabilizing animal populations. We saw in Concept 19.4 that plant production is often higher in more diverse communities and that more diverse plant communities are better able to recover from disturbances. Do these properties convey greater stability to higher trophic levels? Nick Haddad and colleagues set out to test this hypothesis, using experimental prairie plots established by David Tilman at the Cedar Creek Ecosystem Science Reserve in Minnesota (Haddad et al. 2011). They studied the abundance and species composition of arthropod (primarily insect and spider) communities in plots with 1, 2, 4, 8, or 16 plant species over an 11-year period.
A total of 733 different arthropod species were sampled during this period. These arthropods were divided into communities by their feeding preferences, which included detritivores, herbivores, predators, and parasitoids. Stability was evaluated by the amount of change in the abundances of individuals within populations and communities.Haddad et al. found that, in general, the arthropod communities were more stable in the plots with higher plant diversity. However, not all arthropod communities exhibited the same relationship between plant diversity and stability. Populations of specialist herbivores (those that eat one or a few species of plants) had lower stability with increases in plant diversity. In contrast, the community of all herbivores showed greater stability with increasing plant diversity. The researchers suggested that the underlying mechanisms by which plant diversity influences arthropod community stability include greater and more stable plant biomass and increased diversity in the arthropod communities (FIGURE 21.18). Higher plant diversity was linked to greater predator abundance and diversity through its influence on habitat diversity. These predators may exert top-down effects on herbivore and plant abundances (trophic cascades). Haddad et al. also suggested that community stability is enhanced by a portfolio effect, in which variation in the population of one species cancels out variation in another such that overall abundance in the community remains the same. Greater diversity among the arthropods would lead to a greater probability of the portfolio effect. The researchers concluded that plant diversity in the prairie ecosystem provides services to humans not only in the form of potential biofuels, but also by keeping arthropod communities more stable and preventing outbreaks of insects that can be problematic for crops and forests.
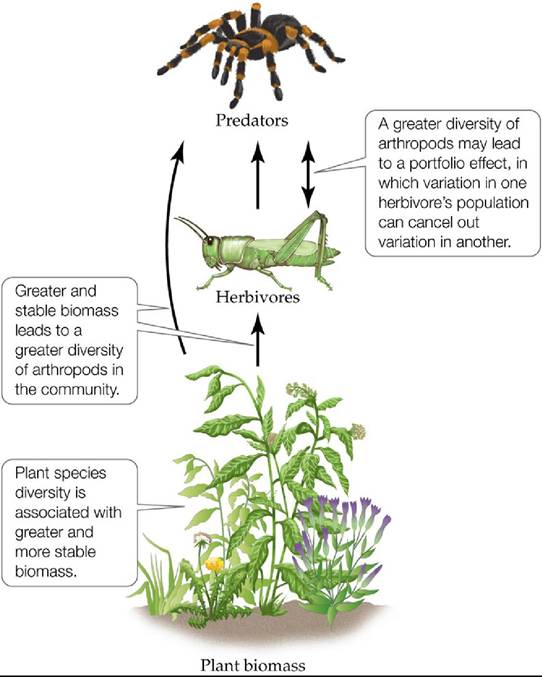
FIGURE 21.18 Plant Diversity and Stability in Food Webs Greaterplantdiversity enhanced the stability of arthropod communities in experimental plots.
The potential mechanisms of this effect include greater and more stable plant biomass. Plant diversity, which is associated with greater habitat complexity, may be associated with greater abundance and diversity of predators, which may lead to greater top-down influences on herbivores and plants (trophic cascades). In addition, plant diversity enhances the diversity of the arthropod community as a whole, enhancing portfolio effects, which keep overall abundance stable. (After N. M. Haddad et al. 2011. Ecoi Lett 14: 42-46.) View larger imageA Case Study Revisited
Toxins in Remote Places
Knowledge of how energy flows through the trophic levels of ecosystems is key to understanding the environmental effects of persistent organic pollutants like those described in this chapter's
Case Study. Some chemical compounds taken up by organisms, either directly from the environment or by consumption with their food, can become concentrated in their tissues. For a variety of reasons, these compounds are not metabolized or excreted, so they become progressively more concentrated in the body over the organism’s lifetime, a process known as bioaccumulation. Bioaccumulation can lead to increasing tissue concentrations of these compounds in animals at successively higher trophic levels as animals at each trophic level consume prey with higher concentrations of the compounds. This process is known as biomagnification (FIGURE 21.19). The POPs we discussed at the beginning of this chapter are particularly susceptible to these processes.
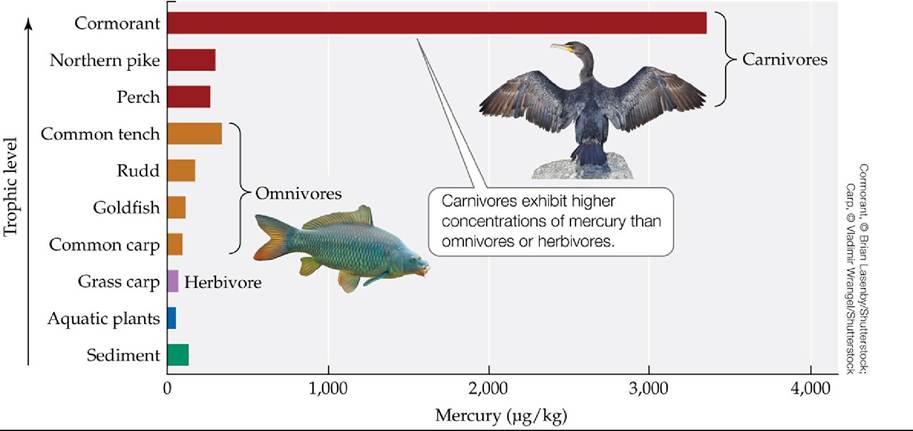
FIGURE 21.19 BioaccumulationandBiomagnification Levels of mercury (a toxic heavy metal) show bioaccumulation and biomagnification in a Czech pond ecosystem. (After P. Houserova et al. 2007. Environ Pollut 145: 185-194.) View larger image
The potential dangers associated with bioaccumulation and biomagnification of POPs were well publicized by Rachel Carson's book Silent Spring, published in 1962, in which she described the devastating effects that pesticides, particularly DDT, were having on nontarget bird and mammal populations.
DDT was thought of as a “miracle” insecticide during the 1940s and 1950s, when it was widely used to control a variety of crop and garden pests and disease vectors. However, DDT was also accumulating in higher- level predators as a result of biomagnification, and it contributed to the near extinction of some birds of prey, including the peregrine falcon and the bald eagle. In Silent Spring, Carson described the persistence of DDT in the environment, its accumulation in the tissues of consumers, including humans, and its health hazards. Because of Carson's careful documentation and her ability to convey her message in a well-crafted manner that could be appreciated by the general public, Silent Spring led to increased scrutiny of the use of chemical pesticides, which eventually resulted in a ban on the manufacture and use of DDT in the United States.The concept of biomagnification led researchers to suspect that the high concentrations of POPs found in the Inuit resulted from their position at the highest trophic levels of the Arctic ecosystem. This suspicion was reinforced by comparisons of the concentrations of toxins among different Inuit communities. The highest concentrations of toxins were found in communities that consumed marine mammals such as whales, seals, and walruses—animals that occupy the third, fourth, or fifth trophic levels. Inhabitants of communities where herbivorous caribou (at the second trophic level) were a more important part of the diet had lower concentrations of toxins. The Inuit preference for foods rich in fatty tissues, such as whale blubber (muktuk), poses a problem as well because many POPs are preferentially stored in the fatty tissues of
animals.
Although emissions of some POPs and other pollutants are declining globally as awareness of their effects increases and regulations are put in place, the potential for long-term persistence of these compounds in the Arctic environment means that their effects may not disappear any time soon (Pearce 1997).
While the cold temperatures and relatively low light levels in the Arctic limit the chemical breakdown of POPs, their concentrations have gradually decreased in lake sediments. There has also been a gradual decline in the concentrations of some POPs and heavy metals in the blood of Inuit individuals, but new emerging POPs and mercury continue to be a concern for public health. While switching to alternative food sources might seem to be a potential solution to the problem, the cultural identity of the Inuit is strongly associated with their hunting traditions and their diet, and they would be unlikely to make such a switch easily.Connections in Nature
Biological Transport of Pollutants
Pollutants have been reported in almost all environments on Earth— even Antarctic ice holds trace amounts of DDT and lead emitted from the burning of leaded gasoline. Animals in many remote areas have high concentrations of industrial and agricultural toxins in their tissues. Fish in isolated alpine lakes of the Canadian Rockies, for example, contain high concentrations of POPs, which have been associated with condensation of these compounds in snowfields and glaciers above the lakes (Blais et al. 1998). As suggested in the
Case Study Revisited, the concentrations of these pollutants are related to the trophic positions of the animals: consumers at the highest trophic levels, such as polar bears, seals, and birds of prey, contain the highest concentrations. The widespread nature of this problem underscores the notion that ecosystems are connected by the movements of energy and materials among them. Ecological processes in one ecosystem can have effects on other ecosystems through these movements (Polis et al. 2004).
The movement of POPs and other human-made toxins is usually associated with atmospheric transport from low to high latitudes. However, the behaviors of animals can also influence the movement of POPs. Salmon, for example, have been shown to transport nutrients from marine to freshwater and terrestrial ecosystems during their spawning runs. At reproductive maturity, salmon leave the ocean and move up rivers in large numbers, as described in the Case Study in Chapter 2. From the rivers, they move into freshwater lakes and streams, where they spawn and then die. The potential exists for salmon to move toxins, as well as nutrients, from the oceans to freshwater ecosystems via this spawning behavior.
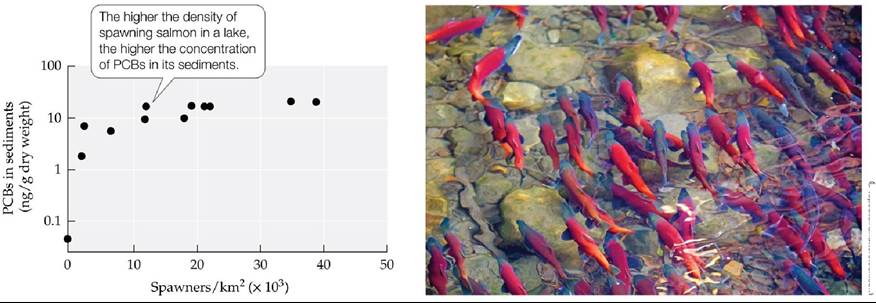
E. M. Krummel and colleagues studied the potential for spawning sockeye salmon (Oncorhynchus nerka) to act as a “fish pump” for pollutants by moving PCBs from the ocean to remote lakes in Alaska (Krummel et al. 2003). Salmon occupy the fourth trophic level, and thus, through bioaccumulation and biomagnification, they accumulate PCBs in their body fat at concentrations more than 2,500 times higher than those found in seawater. Krummel and colleagues collected sediment cores from eight lakes in southwest Alaska that had different densities of spawning salmon (ranging from 0 to 40,000 spawners/km2) and measured PCBs in the sediments.
They found that the concentrations of PCBs were strongly correlated with the density of spawners (FIGURE 21.20). Lakes that did not have visits from spawning fish had concentrations of PCBs similar to expectations based on atmospheric transport alone. The lake with the highest density of spawning fish (40,000 per km2) had PCB concentrations that were six times higher than the levels associated with atmospheric transport. A similar study found that DDT, other POPs, and mercury are transported by northern fulmars (Fulmarus glacialis, pelagic fish-eating seabirds) from the ocean to small ponds near their nesting colonies (Blais et al. 2005). These examples demonstrate how the behaviors of some species (spawning in fish, colonial nesting in birds) can exacerbate problems of pollution associated with biomagnification in ecosystems.
© topseller7Shutterstock.com
FIGURE 21.20 BiologicaipumpingofPoIIutants Spawning salmon act as biological pumps, concentrating toxins from the oceans in their bodies and transporting them en masse to freshwater ecosystems. (After E. M. Krummel et al. 2003. Nature 425: 255-256.) View larger image