Ecological responses to climate change are occurring
As noted earlier, global warming of 0.97°C has occurred since 1880. Several physical environmental changes have occurred over the same period, including the retreat of glaciers, increased melting of sea ice, and a rise in sea level.
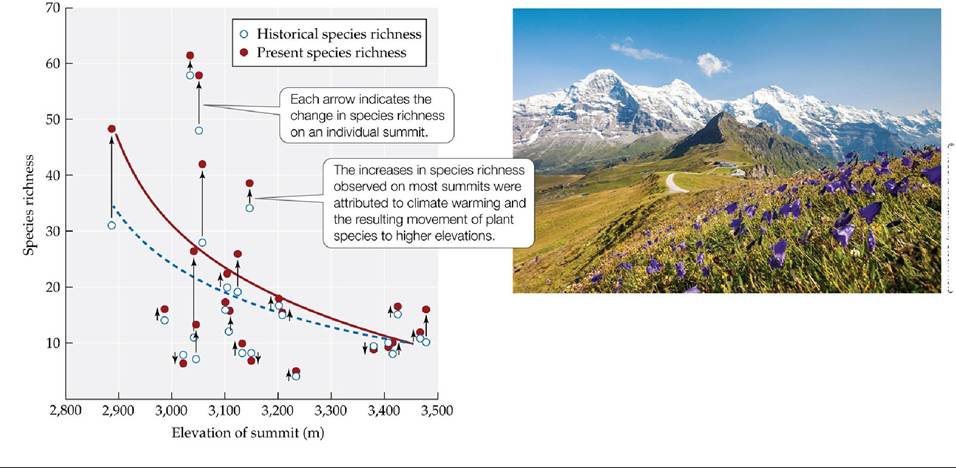
Have ecological systems also responded to this warming? Numerous reports of biological changes are consistent with recent global warming (Parmesan 2006; Walther 2010). These changes include earlier migration of birds, local extinction of amphibian and reptile populations, bleaching of coral reefs, fish die-offs in lakes, and earlier spring greening of vegetation.Although changes in species' range limits are more difficult to link directly to climate change, numerous changes in the geographic ranges of species have been noted that are consistent with warming. For example, Georg Grabherr and colleagues studied the vascular plant communities found on summits of mountains in the European Alps. They compared the current species richnesses of those communities with records dating back to the eighteenth and early nineteenth centuries (Grabherr et al. 1994). They found a consistent trend of increasing species richness resulting from the upward movement of species from lower elevations onto the summits (FIGURE 25.14). Bird species are shifting their ranges to higher elevations in the Andes, with lowland species expanding their ranges, but species at the highest elevations declining in abundance through time (Freeman et al. 2018). Similarly, Camille Parmesan and colleagues recorded a northward shift in the ranges of European nonmigratory butterfly species (Parmesan et al. 1999). Of the 35 species examined, 63% had shifted their ranges northward, while only 3% had shifted their ranges southward. More than half of the plant and animal species that have been investigated have shown geographic range shifts that are consistent with climate change (Parmesan and Yohe 2003).
© Roberto MoioIaZAIamy Stock Photo
FIGURE 25.14 Plants Are Moving Up the Alps Grabherrandcolleaguescompared historical records of vascular plant species richness on the summits of mountains in the European Alps with censuses taken in the early 1990s. The dashed blue curve indicates the relationship between species richness and summit elevation in the historical records, while the solid red curve indicates the present relationship. (After V. H. Grabherr et al. 1994. BioScience 53: 469-480.) View larger image
Climate change may be causing populations of some species to go extinct. Barry Sinervo and colleagues (2010) found that 12% of Mexico's Sceloporus lizard populations had gone extinct between 1975 and 2009. Recall from Concept 23.2 that population extinctions are potentially the initial steps toward the extinction of a species. The extinctions of the lizard populations corresponded more closely to increases in temperature than to losses of habitat. Surprisingly, warming in the spring was better correlated with the extinctions than extreme temperatures during the summer. Sinervo and colleagues concluded that the warmer spring temperatures limited the lizards' foraging time during the breeding season. Ectothermic lizards must move into the shade and remain there to avoid overheating when temperatures become too warm (see Concept 4.2), and during that time they cannot seek out food. The observation that the probability of extinction was greatest at low-elevation and low-latitude sites, where the animals
were most likely to be at the limits of their thermal tolerance, was consistent with this explanation.
Sinervo and colleagues also used a model of lizard thermal physiology to evaluate current and future worldwide effects of climate change on lizard populations. They estimated that climate change has already resulted in extinction of 4% of lizard populations worldwide.
Using projections of future climate change, they suggested that 39% of the world's lizard populations, and 20% of its lizard species, may go extinct by 2050.Migratory animals may also be adversely affected by climate change (Root et al. 2003). For example, migrating marine species, including whales and fish, may need to make longer journeys because of substantial changes in the distributions of their prey species as ocean temperatures warm. Some migratory bird species that breed in England and North America have been arriving at nest sites as much as 3 weeks earlier than they did 30 years ago because of warmer spring temperatures and faster snowmelt. However, plants and invertebrate prey species have responded faster to climate change than the migrating birds, resulting in a mismatch between bird arrival and prey availability. On the other hand, longer breeding seasons may increase the number of offspring produced by some bird species, particularly in high-latitude ecosystems.
Changes in community composition may also be indicators of climate change. These effects may be particularly apparent in some sessile marine species. Concepts 3.3 and 17.1 have described the effects of rising water temperatures on corals and the resulting changes in coral reef communities. Changes in the abundances of marine foraminiferans—a type of zooplankton—also reflect global climate trends during the past century (Field et al. 2006). Foraminiferan species have characteristic shells that allow them to be identified in marine sediments. Cores collected from benthic sediments can be examined to determine changes in the species composition of foraminiferans over time.
Because the environmental tolerances of different species are known, these changes provide a means of reconstructing marine environments of the past. Following the mid-1970s, an increase in tropical and subtropical foraminiferan species, and a decrease in temperate and polar species, occurred in the eastern North Pacific Ocean, indicating a warming of ocean waters there.
Climate change is impacting forest composition in western North America through changes in the frequency and intensity of bark beetle attacks (Anderegg et al. 2015) and forest fires (Dennison et al. 2014; Higuera et al. 2021). Longer frost-free seasons are allowing mountain pine beetles (Dendroctonus ponderosae) in some regions to transition from completing one life cycle per year to two, greatly enhancing their population growth and potential outbreaks (Mitton and Ferrenberg 2012). In addition, the beetles are found at higher altitudes and latitudes than in the past, where they are attacking trees lacking defenses to the beetles. As a result of climate change effects on weather and fuel moisture content, forest fires have doubled since 1984 (Abatzoglou and Williams 2016). Climate change will continue to enhance forest fires until fuels become the limiting factor for their occurrence.
Changes in global NPP also indicate biological responses to climate change. Ramakrishna Nemani and colleagues used remote sensing data to examine global patterns of NPP over an 18-year period (1982-1999) (Nemani et al. 2003). They found that global NPP increased 6% during the study period, or 0.3% per year (FIGURE 25.15). Tropical ecosystems exhibited the largest increase in NPP, which was associated with increases in solar radiation due to less cloud cover in the tropics during the study period. During the first decade of the twenty-first century, however, the trend toward increasing NPP was reversed. The decrease in global NPP during this decade was attributed to major droughts, particularly in the Southern Hemisphere (Zhao and Running 2010).
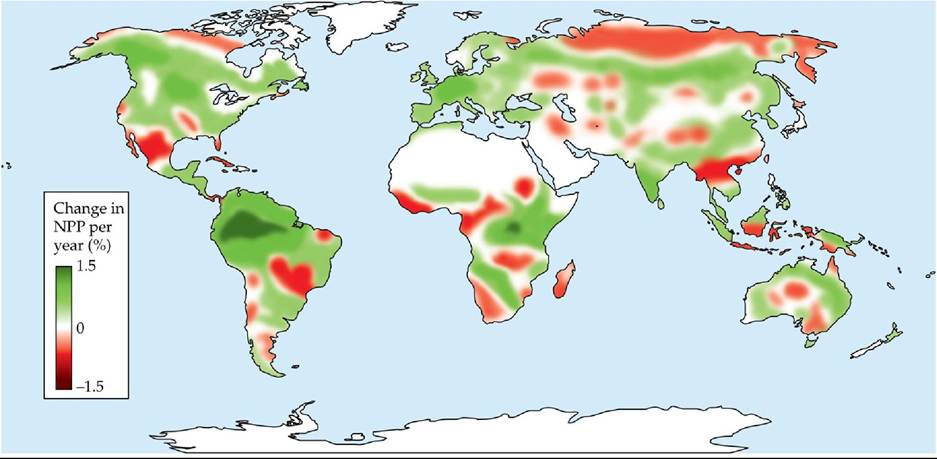
FIGURE 25.15 Changes in Terrestrial NPP Nemani and colleagues calculated changes in net primary production (NPP) between 1982 and 1999, expressed here as percentage change per year. The trend toward increased NPP in tropical regions of South America shown here was reversed in the first decade of the twenty-first century due to prolonged drought.
(From R. R.Nemani et al. 2003. Science 300: 1560-1563.) View larger image
There has been a notable decrease in net ecosystem exchange (see Concept 20.1) in terrestrial high northern latitudes during the past 30 years, which has coincided with some areas of the Arctic switching from a net uptake of CO2 from the atmosphere (acting as a sink) to a net export of CO2 (acting as a source) (Oechel et al. 1993; Zhang et al. 2017). Large amounts of C are stored in the soils of boreal and tundra ecosystems as a result of low-temperature constraints on decomposition and the long-term buildup of carbon since the last glacial maximum. Warming during the twentieth century, however, increased the rate of CO2 export from Arctic soils, such that losses now exceed gains from NPP. Warming of these high-latitude terrestrial ecosystems could provide a positive feedback to climate change by enhancing their emissions of CO2 and CH4. The rates of CO2 loss from Arctic ecosystems have decreased since the early 1990s, possibly due to changes in rates of nutrient cycling and physiological and compositional changes in the plants (Oechel et al. 2000). However, greater loss of CO2 due to heterotrophic consumption of soil organic matter during winter may offset the gains from greater summer productivity (Webb et al. 2016). During this same time period NPP of Arctic oceans has increased due to decreases in ice cover, warming, and greater reception of solar radiation and increased inputs of nutrients from rivers and coastal erosion (Terhaar et al. 2021).