Environmental context can change the outcome of species interactions
As we have seen in this section, interactions among species can vary in strength and direction, and their outcomes are highly dependent on the influence of each of the species in the community.
As we saw in Unit 4 and Ecological Toolkit 16.1, another important factor in the outcome of species interactions is the environmental context in which they occur. For example, under benign environmental conditions that are favorable for population growth, it makes sense that species will thrive and be limited by resources and will therefore engage in negative interactions such as competition or predation. Under harsh environmental conditions, species will naturally be more strongly limited by physical factors and will therefore interact either weakly or positively with other species.This view of species interactions as context dependent, or changeable under different environmental conditions, is relatively new to ecology, but a number of important examples of context dependence exist. Most of these examples involve keystone or foundation species that play important roles in their communities in one context, but not in another. Mary Power, a professor at the University of California, Berkeley, who works on stream communities in Northern California, has shown that the role of fish predators (roach [Hesperoleucas symmetricus] and steelhead [Oncorhynchus mykiss]) changes from year to year. The role of these predators shifts from that of keystone species following winters of scouring floods to that of weak interactors during years with winter droughts and in places where flood control is operating (Power et al.
2008).
In the Northern California rivers where Power works, there is a natural winter flood regime that produces dramatic population cycles of the green filamentous alga Cladophora glomerata. In most years, scouring winter floods remove most of the inhabitants—particularly armored herbivorous insects such as large caddisfly larvae—from the river bottom.
In the following spring, there are large blooms of Cladophora. Increased light and nutrients and the lack of invertebrate herbivores allow Cladophora mats to grow profusely over the rocks, producing filaments up to 8 m (26 feet) long. By midsummer, these mats detach from the rocks and cover large portions of the river, at which time midge larvae, which feed on the floating alga and use it to weave small homes, increase in number. The midges are fed on by small fish and damselfly larvae, which in turn are eaten by steelhead and roach (a trophic cascade with four levels; FIGURE 16.19A). The steelhead and roach are able to decrease the size of the algal mats by eating small fish and damselfly larvae, which feed on midge larvae, which feed on the mats. The roach also feed on the algae directly, but only to a small degree.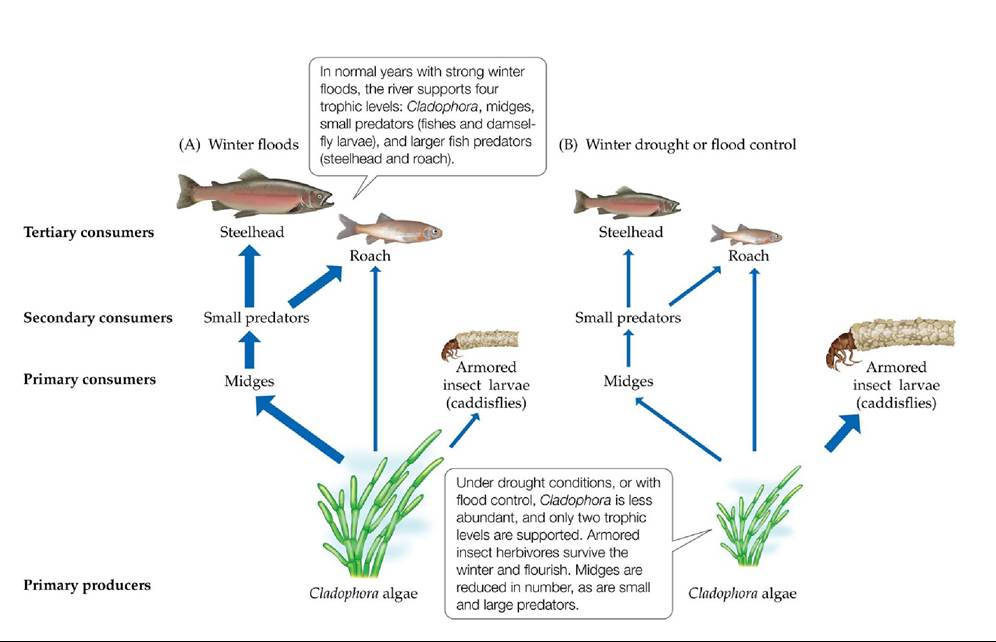
FIGURE 16.19 Context Dependence in River Food Webs Environmental changes alter the relative importance of different trophic levels in the Eel River of Northern California during winter floods (A) and winter droughts or flood control conditions (B). Wider arrows represent stronger interactions. (After M. E. Power et al. 1996a. In Food Webs: Integration of Patterns and Dynamics, G. A. Polis and K. O. Winemiller [Eds.], pp. 286-297. Chapman & Hall: New York.) View larger image
During drought years, however, and in rivers where flood control is operating, flooding and scouring of the river bottom do not occur. In those years, Cladophora persists but does not form large, lush mats. Power and colleagues showed that this change was due to the presence of more armored herbivorous insects, which were not removed by floods and which ate the Cladophora while it was still attached to the rocks. This interaction led to declines in Cladophora and the loss of the detachment phase of the alga. The armored insects are much less susceptible to predation than the midges and thus are not controlled by higher trophic levels.
In essence, the typical river food web with four trophic levels is converted into a two-level food web during drought years, and the steelhead and roach, which in flood years are keystone predators, become minor players in the food web (FIGURE 16.19B).Z
Climate Change Connection
Context Dependence of Ocean Acidification
One emerging environmental context that will affect communities is ocean acidification. Estimates show that oceans are absorbing about 48% of all atmospheric CO2. Marine primary producers use some of the human-created CO2 in photosynthesis, but the remainder reacts chemically with seawater, lowering its pH and causing oceans to become more acidic (see Concept 25.1). Ocean acidification can have negative effects on calcifying organisms such as corals, mollusks, and crustaceans, which rely on calcium carbonate for the accretion and maintenance of their external shells. But the negative effects of increasing CO2 and acidification are not universal. For example, primary producers such as seagrasses, algae, and phytoplankton are known to increase their productivity under elevated CO2. Although the effects of ocean acidification on the physiology of single species is a growing field of study, much less is known about how a world with a lower pH will affect the structure and function of communities. Moreover, along with the acidification of oceans, temperatures are increasing, creating multiple stressors. As a result, marine ecologists are asking, How will species interactions, both direct and indirect, be influenced by ocean acidification?
Christian Alsterberg and his colleagues (2013) considered how a food web in an estuarine community on the western coast of
Sweden was influenced by ocean acidification and warming. The researchers chose to focus on single-celled microalgae living in the sediments and their interactions with macroalgae and consumers. Under normal conditions in this estuarine system, omnivores (i.e., a guild of medium-sized crustaceans and snails that feed on two trophic levels) increase the productivity of benthic microalgae in two ways: (1) they increase the light available to microalgae by feeding on macroalgae, and (2) they prey on herbivores of microalgae (i.e., a guild of small crustaceans, snails, and worms) (FIGURE 16.20A).
To estimate the importance of ocean acidification and increased warming in a multispecies food web, the researchers conducted an experiment in which CO2 and temperature, as well as omnivores, were manipulated in mesocosms (in this case, large aquariums). After a 5-week period, the experiment showed that with elevated CO2 and temperature, macroalgae and herbivores increased in biomass. When omnivores were present, this greater biomass was consumed by omnivores, resulting in a stronger negative interaction between them and their food sources (FIGURE 16.20B). The stronger negative interaction lead to a stronger indirect positive effect for benthic microalgae. As a result, there was no change in benthic microalgae abundance despite the elevated CO2 and temperature conditions. However, when omnivores were absent, macroalgae and herbivores were not kept in check, and their negative interactions with benthic microalgae grew even stronger, resulting in the decline of benthic microalgae. Thus, this experiment showed that consumers were able to modulate the negative effects of elevated CO2 and temperature on primary producers in this estuarine system. This study highlights the importance of understanding how organisms, placed within the context of their food web communities, gain some resilience against the effects of climate change.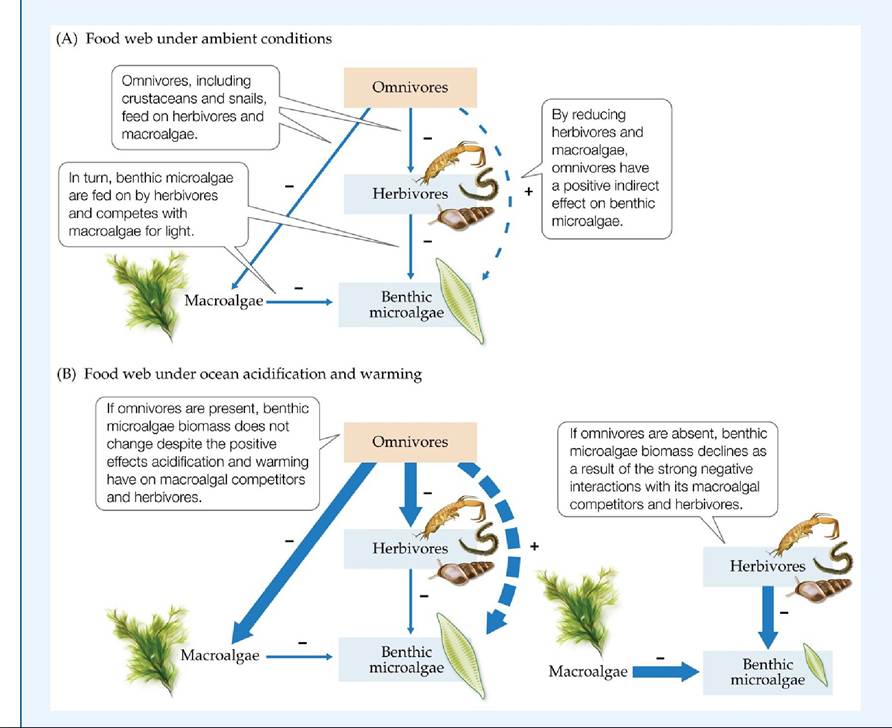
FIGURE 16.20 Food Webs in an Acidic and Warming World (A) The interaction web of species in an estuarine community off the western coast of Sweden. (B) The effects of ocean acidification and warming on the interaction web with (left) and without (right) omnivores. The biomass of benthic microalgae did not change with omnivores (left) but declined without them (right). Thicker arrows represent stronger interactions. (After C. Alsterberg et al. 2013. Proc NatlAcad Sci USA 110: 8603-8608.) View larger image
In upcoming chapters, we will consider in much more detail the effects of physical and biological factors on the outcome of species interactions and ultimately on the diversity of communities.
A Case Study Revisited
“Killer Algae!”
The introduction of Caulerpa taxifolia into the Mediterranean Sea in the early 1980s set in motion a series of unfortunate events that resulted in large carpets of fluorescent green algae dominating formerly species-rich nearshore marine communities. Caulerpa thrived because humans facilitated its dispersal and its physiological tolerance. Even in the early stages of the invasion, Meinesz documented the seaweed in at least three types of communities, with different species compositions, on rocky, sandy, and muddy substrates. Together, these communities are home to several hundred species of algae and three marine flowering plants, as well as a number of animal species. Once Caulerpa arrived, native competitor and herbivore species were unable to keep it from spreading.
The invasion of Caulerpa has changed the ways in which native species interact with one another, and thus the structure and function of the native communities. One obvious consequence of the presence of Caulerpa is the decline of seagrass meadows dominated by Posidonia oceanica (FIGURE 16.21). This seagrass has been likened to an “underwater tree” because of its long life span and slow growth (patches grow to 3 m in diameter in 100 years). Just like forests, seagrass meadows support a multitude of species that use the vegetation as habitat. Research showed that Posidonia and Caulerpa have different growth cycles: Posidonia loses blades in summer, when Caulerpa is most productive. Over time, these asynchronous growth patterns result in Caulerpa overgrowing the existing seagrasses and establishing itself as the foundation species. Additional research has shown that Caulerpa acts as an ecosystem engineer, accumulating sediments around its roots more readily than Posidonia, which can change the species composition of the small invertebrates that live on the seafloor. Some surveys have revealed a significant drop in the numbers and sizes of fish using the communities invaded by Caulerpa, suggesting that these habitats may be less suitable for some commercially important species.

FIGURE 16.21 AMediterraneanSeagrassMeadow Native communities like this one, dominated by the seagrass Posidonia oceanica, can be replaced by invasive Caulerpa taxifolia. Compare this photograph with Figure 16.1. View larger image
Future changes in Mediterranean seagrass meadows, and in the species dependent on them, will be difficult to predict, given the sheer number of species that are potentially affected by Caulerpa, the indirect effects that will be generated by changing interactions, and the relatively short time that has elapsed since the invasion began. A scientific approach, guided by a combination of theory and real-world observations, will be necessary if future predictions are to be made about the ultimate effect of Caulerpa on this potentially vanishing underwater community.
Connections in Nature
Stopping Invasions Requires Commitment
Even though it may be too late to stop the invasion of Caulerpa taxifolia in the Mediterranean, the lessons learned there have been important in other regions of the world. In 2000, just as Meinesz was making progress in banning international trade of the alga, he received an e-mail from an environmental consultant in San Diego, California. While surveying eelgrass in a lagoon, she had noticed a large patch of what was later identified as Caulerpa taxifolia. Acting on Meinesz's recommendation, a team of scientists and managers from county, state, and federal agencies immediately assembled to design an eradication plan. This plan involved treating the alga with chlorine gas injected under tarps placed on top of algal patches. More than $1 million was initially budgeted for the project in 2000, but it eventually took 6 years and $7 million to eradicate the alga. The invasion was widely publicized, resulting in the discovery of another patch of Caulerpa in another lagoon near Los Angeles, which was also eradicated. The California experience is a rare success story only because immediate action was taken by scientists, managers, and policymakers to deal with the invasion before eradication became an ecological and fiscal impossibility. To determine the origin of the Caulerpa that invaded California, molecular evidence was needed. This shift in the team's focus from communities to genes illustrates a point made in Chapter 1: ecologists must study interactions in nature across many levels of biological organization. The team sent specimens of the alga to geneticists at two universities, who analyzed the sequences of its ribosomal DNA and quickly determined that they were identical to those of Caulerpa from the Mediterranean, the Wilhelma Zoo (where the strain was first cultivated), and many other public aquariums around the world (Jousson et al. 2000). Unfortunately, it is still unknown how the species was introduced into the two California lagoons, but hypotheses range from amateur aquarists cleaning their tanks in the lagoons to an accidental release from aquariums on board a Saudi Arabian prince's yacht, which was being repainted in San Diego at about the time the alga probably arrived. Through the use of DNA analysis, it has been determined that the Caulerpa algae involved in subsequent invasions in Australia and Japan are genetically identical to the original German Caulerpa taxifolia strain. The molecular evidence makes it clear that the trade of this alga in aquarium circles poses a global threat to nearshore temperate marine environments. Legislation is now in place to ban the “killer alga” from a number of other countries where it has a good chance of invading successfully.