Group size may reflect a balance between the costs and benefits of group living
If we apply the cost/benefit principles discussed in this chapter to group size, we might predict that groups should be of a size at which the benefits of belonging to a group exceed the costs.
For example, using an approach similar to that introduced in Concept 8.2 for optimal foraging, we could predict that groups will have an “optimal” size—the size at which the net benefits received by its members are maximized. However, as shown in FIGURE 8.22, unless group members can prevent other individuals from joining the group once an optimal size is reached, the observed group size may be larger than the optimal size. In addition, it can be very difficult to measure all the benefits and costs of group living; it is particularly challenging to quantify both costs and benefits with a single “currency,” such as energy use or offspring production.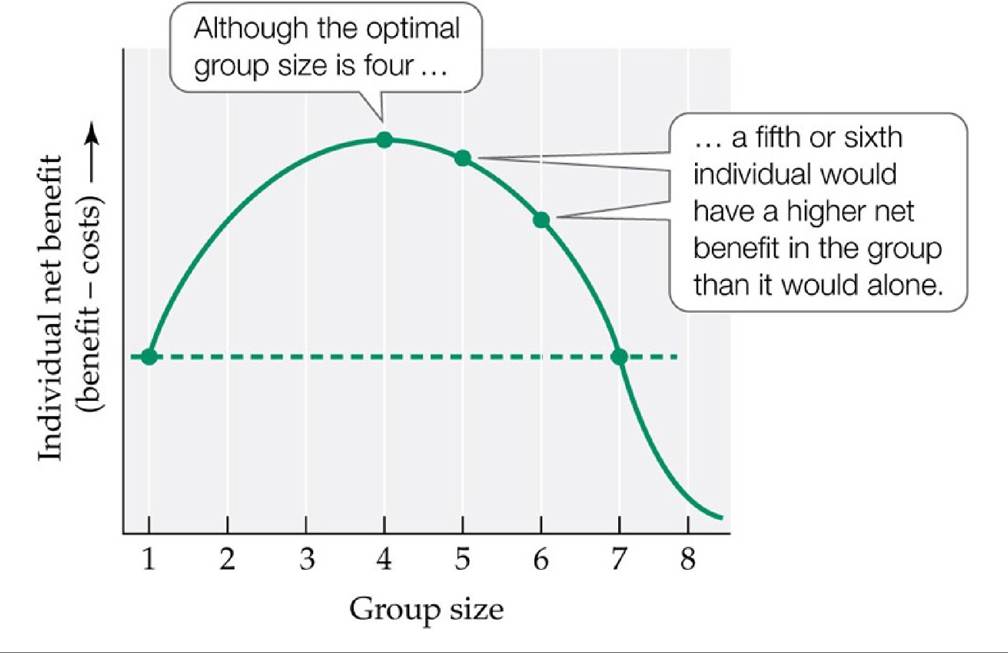
FIGURE 8.22 Should a New Arrival Join the Group? In this hypothetical example, the net benefit to an individual group member is maximized at the optimal group size of four. However, a new arrival would have a greater net benefit if it joined a group of size four than if it remained alone. Unless existing group members can prevent new arrivals from joining the group, new arrivals should continue to join until the group has reached a size of seven—at that point, the individual would do better on its own than in the group. View larger image
In general, an argument like that in Figure 8.22 suggests that it may be advantageous for individuals to belong to groups that are larger than the optimal size, but not so large that a new arrival would do better on its own. Such an intermediate-sized group might be large enough to reduce the risk of predation, but small enough to avoid running out of food. Using an overall measure of individual condition (level of stress as measured by fecal concentrations of the hormone cortisol), Pride (2005) found that ring-tailed lemurs in groups of intermediate sizes were less stressed than lemurs that belonged to smaller or larger groups.
Similarly, Creel and Creel (1995) found that the per capita intake of food for Tanzanian wild dogs chasing prey was greatest for packs of intermediate sizes.A Case Study Revisited
Infanticide in Lion Packs
Can an evolutionary perspective on behavior help us to understand infanticide? It turns out that the males of many species kill the young of their potential mates. For example, male langur monkeys (Semnopithecus entellus) kill the infants of females in their social group. This behavior appears to increase the reproductive success of the murderous males: DNA paternity analyses showed that infanticidal male langurs were not related to the infants they killed but were related to the females' subsequent offspring (Borries et al. 1999). Infanticide by males has been documented in dozens of other species, including horses, chimpanzees, bears, and marmots. Infanticide by males appears to be adaptive in many cases: it reduces the time that females spend between estrous cycles, thus enabling the males to sire more offspring than they otherwise could.
But in some species, females commit infanticide. For example, female giant water bugs (Lethocerus deyrollei) and female wattled jacanas (a type of shorebird, Jacana jacana) destroy the eggs or young of their own species. While gruesome, this behavior makes evolutionary sense: in these species, the males provide most or all of the parental care, and the females have higher reproductive potential than the males. Thus, as is true for male lions and langurs, the infanticidal behavior of female water bugs and jacanas appears to be adaptive: by killing the young, the time before the male is willing to mate is shortened, thus potentially increasing her own reproductive success.
What about other puzzling behaviors mentioned in this chapter's Case Study? Recall that female fruit flies (Drosophila melanogaster) sometimes lay their eggs in foods that are high in ethyl alcohol. This behavior is not as strange as it first appears: evidence suggests that it provides a behavioral defense against the wasp Leptopilina heterotoma, which preys on the fruit flies.
Females of this wasp lay their eggs on fruit fly larvae; when an egg hatches, the young wasp burrows through the body of the fly larva, consuming and killing it. A fruit fly larva infected by this wasp will preferentially choose to eat foods that are high in alcohol content, such as rotting fruit, as it helps defend it from the wasp young. Even though consuming foods containing high concentrations of alcohol harms the fruit fly larvae, the benefits of this action outweigh its costs: wasps are more susceptible to the harmful effects of the alcohol than the fruit flies, increasing the overall chances that the fruit fly larvae will survive. In addition, Kacsoh et al. (2013) showed that adult female fruit flies altered their egg-laying behavior in response to the presence of wasps. In the absence of wasps, the fruit flies laid about 40% of their eggs in high-alcohol foods, but when female wasps were present, the fruit flies laid over 90% of their eggs in high-alcohol foods. This behavior increased the survival of fruit fly larvae exposed to wasps (FIGURE 8.23), suggesting that the behavior canbe viewed as a type of preventative medicine.
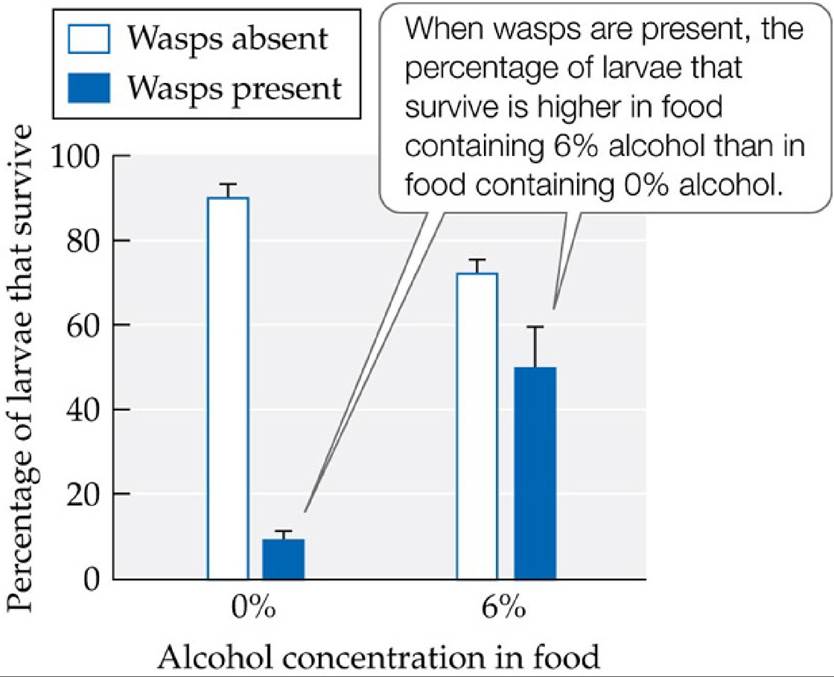
FIGURE 8.23 Fruit Flies Medicate Their Offspring Female fruit flies (Drosophila melanogaster) laid most of their eggs in foods containing alcohol when females of the wasp Leptopilina heterotoma were present. This behavior increased the percentage of fruit fly larvae that survived to adulthood. Error bars show one SE of the mean.
In the absence of wasps, what is the cost (in terms of reduced larval survival) of laying eggs on food containing 6% alcohol? When wasps are present, what is the benefit?
(After B. Z. Kacsoh et al. 2013. Science 339: 947-950.) View larger image
Connections in Nature
Behavioral Responses to Predators have Broad Ecological Effects
As you've seen, individuals often change their behavior in response to predators.
For example, in Concept 8.2, we saw that when exposed to recordings of sounds made by predators, song sparrows fed their young less often, built their nests in less desirable areas, and spent less time incubating their eggs (Zanette et al. 2011). What were the consequences of these behavioral changes?Zanette et al. found that when song sparrow parents altered their behavior in response to a perceived high risk of predation, their offspring lost body heat more rapidly (FIGURE 8.24A) and weighed less than did the offspring of sparrows exposed to recordings of nonpredators. These effects on individual offspring appear to have caused the number of offspring produced per year to decline (FIGURE 8.24B). Overall, the results of this study suggest that fear of predation alters the behavior of song sparrows in ways that decrease their reproductive success and may cause their population sizes to drop.
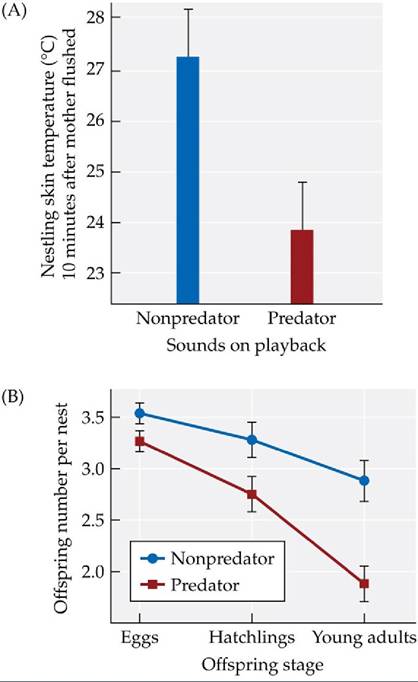
FIGURE 8.24 Costs of Fear (A) The rate at which young song sparrows lost body heat was higher for offspring of birds exposed to playbacks of predators than it was for offspring of parents exposed to playbacks of nonpredators. Error bars show one SE of the mean. (B) Fewer offspring were produced in nests exposed to playbacks of predators than in nests exposed to playbacks of nonpredators. Error bars show ± one SE of the mean.
Estimate the cost of fear on the number of offspring that survived to young adulthood.
(After L. Y. Zanette et al. 2011. Science 334: 1398-1401.) View larger image
Behavioral responses to predators can also affect ecosystem processes, such as the decomposition of leaves and other plant litter in soil and subsequently the availability of nutrients (Concept 22.2). The presence of spider predators can lead to a series of events in their grasshopper prey that ultimately slowed the decomposition rates of plant litter. How did this happen? Hawlena et al.
(2012) found this effect occurred indirectly via changes in foraging behavior. When the researchers raised grasshoppers in the presence of predators, the grasshoppers became physiologically stressed, one consequence of which was that they required more energy to maintain their basic body functions. This demand for additional energy appears to have altered their foraging behavior, leading the grasshoppers to increase their consumption of plants that are high in carbohydrates (and thus in energy) but low in nitrogen. Thus, grasshoppers stressed by predators had a higher carbon:nitrogen ratio in their bodies than did grasshoppers raised in the absence of predators. Although this change in nutrient content did not affect the decomposition of the grasshoppers’ own bodies, it did decrease the decay of plant materials in the soil. This probably happened because the altered carbon:nitrogen ratio in the decomposing bodies of grasshoppers affected the carbon:nitrogen ratio in the soil, which in turn affected the community of soil microorganisms that decompose leaves and other plant matter.