Historical definitions of competition
I will not attempt a complete historical account of the term ‘competition’ in the linked fields of ecology and evolution. For more details on the early history see Hutchinson (1965) or Kingsland (1995).
The term was already in common usage when Lotka and Volterra independently introduced the first mathematical model of interspecific competition in the 1920s (presented in Chapter 1). I will begin this history with the early 1970s, corresponding to the beginning of my own formal study of ecology. At that time, most universities did not include departments devoted to ecology and evolution, and the vast majority of the current body of published work on competition had yet to appear.The year before I began my graduate studies, a short book summarizing population biological theory for beginning students was published, ‘A Primer of Population Biology’, by two Harvard professors, E. O. Wilson and W. H. Bossert. Their definition of competition was, ‘the active demand by two or more organisms for a common vital resource’ (Wilson and Bossert 1971, p. 156). This may be a little too succinct, as there is quite a bit of leeway in what constitutes ‘a common vital resource’ and ‘active demand’. Subsequent authors usually have referred to ‘consumption’ rather than ‘active demand’ Nevertheless, this definition is close to the one that I will support here. Although Wilson and Bossert’s definition of competition applies to both within- and between-species forms, these two types were discussed separately in their book, and within-species competition was, in most instances, referred to as ‘density-dependence’ The mathematical models of both within- and between-species processes that they discussed failed to include any explicit representation of the abundance or dynamics of the resources. Their models expressed the population growth rate of the competing consumers solely as a function of their own abundances.
Shortly after Wilson and Bosserts book appeared, C. J. Krebs (1972) published the first comprehensive modern textbook covering the entire field of ecology. He endorsed L. C. Birch's earlier definition (Birch 1957), that ‘Competition occurs when a number of organisms (of the same or different species) utilize common resources that are in short supply; or, if the resources are not in short supply, competition occurs when the organisms seeking that resource nevertheless harm one or other in the process’. This definition maintains the centrality of resources, but it introduces the possibility that there are additional mechanisms, beyond simple consumption, which may influence the effects of one resource consumer on another. Interestingly, Birch (1957) was motivated to publish the above definition because he felt multiple definitions of competition were being used, and that ‘Some of these meanings are so ambiguous that the word has largely lost its usefulness as a scientific term' (Birch 1957, p. 5). Neither Birch nor Krebs has a detailed discussion of what constitutes a single resource. This is not really required for the definition, but it does have an important effect on the population-level consequences of resource consumption for the consumers and the resources. In addition, it is crucial for assessing whether species will be able to coexist, as will be discussed in later chapters.
The second part of Birch and Krebs' definition is the issue of‘harm... in the process’. This part of the definition is now usually described as interference competition. ‘Harm' may sound like a well-defined term, but several different metrics could be used to distinguish between harmful effects. Does the harm decrease the immediate per individual population growth rate or the immediate resource intake rate, or can it increase one or both provided that it decreases the ultimate population size? As will be explained below, these different ways of quantifying ‘harm' do not always result in the same classification of the interaction.
Harm is usually considered to include interference with intake of resources, interference with reproduction, or the killing or injuring of other individuals.Models of competition should reflect the definition of the term. Fifty years ago, most ecological researchers who thought about modelling competition would have employed the Lotka-Volterra (LV) model presented in Chapter 1. The defining feature of the interaction between two competitors in that model is that the immediate per capita growth rate of each species declines with an increase in the abundance of the other. By 1970, this had been the predominant model of competition for over four decades. It was the only model discussed by Wilson and Bossert (1971). Unfortunately, measuring the per capita growth rate at one point in time requires two population measurements that are close in time. This is difficult and entails large uncertainties. Thus, most investigators studied the long-term change in population size in one species following addition or removal of a second species. We will see later that both the linearity and the immediacy of the effects on population growth rate are problematic if one considers the dynamics of the resources that are the subject of the interaction. In any event, the LV model lacked resources, so it was at least somewhat inconsistent with definitions based on resource use.
As was noted in Chapter 1, Robert MacArthur was largely responsible for the major increase in the scientific study of competition that began in the late 1960s. In his final book in 1972 (p. 21), MacArthur proposed that, ‘two species are competing if an increase in either one harms the other. Any machinery that can have that effect will be called competition.' The ‘harm' was not defined, but it is clearly meant to apply at the level of the population rather than the individual. MacArthur's usage of the term strongly suggests that he was interpreting ‘harm' as a decrease in the size of one species' population caused by an increase in that of another species.
Notably, he mentions that the mutual negative effects of two species that came about by increasing the population of a shared predator would qualify as competition. This separates the definition from a requirement that it involve resources and puts the emphasis on the negativity of the effect on population size.While on the subject of definitions, readers should be aware that ‘predator' in this book will be used to describe any consumer of a living resource, with ‘prey' being any living entity consumed by a predator. Thus, ‘predator' should be understood to include herbivores, and ‘prey' includes plants. This book will largely ignore parasites and diseases, which live within their host (resource), and therefore require a different modelling framework.
The status of the interaction via shared predators is still somewhat uncertain. MacArthur did not attempt to model this interaction directly or discuss its role in consumer-resource interactions. It was later named ‘apparent competition' by Holt (1977) in an article that developed the first formal theory regarding the populationlevel effects of shared predation. Most recent articles seem to regard interactions via shared predators as a different category from ‘normal' competition (see Holt and Bonsall 2017). Apparent competition will be treated in detail in Chapter 5.
Another definitional grey area in the 1970s was the situation in which individuals of each of two species kill and consume individuals of the other species, i.e. mutual predation. The problem with mutual predation is that it is possible for one species to benefit more from its consumption of its competitor than it loses from negative effects of that competitor. This could produce (+,-) effects of the two predators on each other's abundance. It is even possible that it could produce (+,+) effects on population size. Neither mutual predation nor apparent competition is included in most recent discussions of interspecific competition, although Kuang and Chesson (2008) have argued that apparent and resource competition are identical in many respects.
Despite his definition emphasizing population-level effects, MacArthur (1970, 1972) was the first biologist to explore how shared resource use by two consumers could lead to the model of competition suggested by Lotka and Volterra. He showed that if four conditions were met, this model could approximate the consumers' dynamics in a consumer-resource model. These conditions were: (1) all of the resources had logistic population growth; (2) none of the resources interacted directly with others; (3) resource dynamics were fast enough relative to the consumer that all resources quickly reached a quasi-equilibrium state following any change in consumer population size; and (4) the change in consumer population(s) did not cause extinction of any resources. However, MacArthur did not actually investigate when conditions (3) and (4) would be satisfied, and he did not even acknowledge condition (4). A 2-resource version of his model is explored in Chapter 3, to examine the consequences of these omissions. In any case, this quasi-consistency of the LV model with one very simple resource-based model led MacArthur to develop additional theory in which resources were not considered explicitly (May and MacArthur 1972).
The derivation in MacArthur (1972) may have contributed to the later popularity of definitions of competition based purely on negative effects on equilibrium population size. Negative effects are in fact the only possibility under the LV model of two competitors. MacArthur had previously published theory based on the LV model (MacArthur and Levins 1967), so his interpretation of the consumer-resource models may have been biased towards finding consistency between the two approaches. In any event, most subsequent authors adopted MacArthur’s definition of competition in terms of effect on ultimate abundance rather than mechanism. Paired negative effects on abundance were assumed to be the definitive evidence for the occurrence of competition in subsequent experimental studies (Connell 1983; Schoener 1983) and were also thought to be a prerequisite for concluding that coupled evolutionary changes in two species were a consequence of competition (Schluter 2000).
MacArthur’s focus on competition helped inspire many subsequent theoretical and empirical works. A large number of articles in the 1970s expanded our understanding of competitive interactions beyond the confines of the LV model. Laboratory systems involving Drosophila showed that competitive interactions were highly nonlinear (Gilpin and Ayala 1973; Ayala et al. 1973). In those systems, the effect of adding a fixed number of individuals of species 1 on the growth rate of 2 was relatively smaller when the initial abundance of species 1 was larger. This contrasts with the density-independent effect assumed by the LV model. A series of theoretical works by Schoener (1973,1974c, 1976,1978) explored the role of resource dynamics in producing different types of nonlinearity, as did Leon and Tumpson (1975) and Abrams (1975, 1977).
Stephen Levine (1976) called into question the assumption that mutually negative effects always occur as a consequence of the joint use of resources. He analysed simple mathematical models of two important cases in which shared use of resources could produce positive effects of a change in the number of individuals of one species on the abundance of another. The first case involved two consumers that shared two resources, but the resources had negative effects on each other’s per capita growth rate. If consumer 1 had a strong negative effect on its preferred resource (resource 1), and that resource had a strong negative effect on consumer 2’s preferred resource (resource 2), then the effect of increasing the abundance of consumer 1 would be an increase in consumer 2. In other words, they would not ‘compete’ according to a definition based on effects on long-term population size. Vandermeer (1980) provided a more detailed treatment of this case.
The second of Levine’s examples used the LV model, but assumed three competing species. In this system, an increased abundance of consumer species 1 could have a positive effect on the long-term abundance of species 2. Positive effects occurred when greater numbers of species 1 caused a sufficiently large decrease in species 3, which was a stronger competitor of species 2. The indirect positive effect via competitor 3 could easily be larger than the direct negative effect on species 2, reversing the sign of the net effect of species 1 on species 2. Both of Levine's examples were common occurrences in the set of simple models he examined, and I am unaware of any subsequent work providing a good argument for why the mechanisms they represent should be rare in nature. However, there was no immediate response by the ecological community to resolve the problems that these cases represented for the definition of competition. Later work has revealed a large range of other mechanisms involving joint use of resources that have the possibility of (+,-) or (+,+) effects (e.g., Abrams and Cortez 2015a), as well as cases in which the interaction sign depended on the magnitude of the perturbation to the ‘initiating' species.
Grime (1979) addressed the conflict between definitions based on mechanism vs those based on effect (directions of change in abundance), in a book focused on plant ecology. He suggested the use of shared resources as the basis for defining competition. The early 1980s saw other works that promoted a resource-based view of competition. Tilman's 1980 article and 1982 book were particularly influential, although he made a point of referring to ‘resource' competition, implying the existence of other forms. A broader discussion of the proper definition and/or minimal model of competition was apparently cut short by a more prominent dispute, the debate over the ‘importance' of competition for determining the composition of natural communities. Chapters for and against competition may be found in the book organized and edited by Strong et al. (1984). The contributing editors, all from Florida State University, were all on the ‘anti-competition' side of the debate, but both sides were well represented in the individual chapters. The main issue of this controversy was whether geographic patterns in the co-occurrence of species could be interpreted as strong evidence for the operation of interspecific competition. A second issue was whether differences in the body sizes or feeding morphologies of coexisting species provided evidence that competition had caused these differences. The subsequent rapid decline in empirical research on competition (Grover 1997) suggested that the critiques of Strong and his associates might have discouraged studies of competition in the following years. Strong et al.'s book was based on a conference that took place in 1981; by the time the book finally appeared, two comprehensive reviews of field studies of competition (Schoener 1983; Connell 1983) had already appeared. Neither of these was based on the correlational evidence that was the subject of the controversy, and each of these reviews found that interspecific competition was observed in the vast majority of studies that were designed to assess whether it occurred. The evidence for competition in those studies was a significant decrease (increase) in population size (measured over some time frame) in response to an experimentally imposed increase (decrease) in the abundance of a co-occurring species on the same trophic level. This evidence of widespread competition was unfortunately not sufficient to prevent a loss of interest in studying the interaction.
The analyses in Connell and Schoener's review articles might seem to indicate a shift towards a definition of competition based on the effect on ultimate population size. However, Connell (1983) claimed to agree with Birch's (1957) definition of competition. Although this definition was resource-based, Birch seemed to assume, without explicitly requiring, mutually negative effects on population size. Schoener (1983) did not formally define competition, but did require that the species occupy the same trophic level and that at least one of a pair of competitors decreases in response to an increase in the other. This again seems to indicate a shift towards a double requirement of both mutually negative effects and a mechanism based on shared resource use. Schoener (1983) divided competition into six types; consumptive, preemptive, overgrowth, chemical, territorial, and encounter. Preemptive, overgrowth, and territorial interactions all involve space as a resource (or a proxy for the resource). Chemical and encounter both involve harm inflicted by individuals of one species on individuals of the other. Thus, this scheme is more of a subdivision of the two recognized categories of exploitative and interference competition, than a fundamentally new classification. Although it might seem that some interference effects are independent of resources, it does not make evolutionary sense for individuals of one consumer to kill, chase, or harm members of another species (which entails time, energy, and possibly risk of injury) unless there are increases in resource availability that subsequently result from these actions. Most of these scenarios argue that a proper definition of what constitutes a single resource needs to take location or time into consideration. It is possible that, in some cases of interference, the ‘harm’ is a side effect of some other activity, and has no real cost to the perpetrator. The latter possibility no doubt occurs in some cases (e.g., elephants stepping on insects), but is likely to be rare in systems with mutually negative effects.
While the definitions of interspecific competition assumed in Connell’s and Schoener’s 1983 reviews are not purely based on co-utilization of resources, it appears that almost all (if not all) of the studies they review that showed mutually negative effects would have also qualified as ‘competition’ based on such a requirement. However, in both of these reviews, the authors identified small numbers of cases that did not exhibit such effects. Were these few outcomes a consequence of Levine’s or some other mechanism, such as limitation of abundance by separate predators? So far as I know, this was never investigated. These two reviews also do not shed any light on whether cases that lacked significant negative effects on population density also lacked effects on resource abundances.
Proceeding to the 1990s, the range of views on the definition of competition was still quite wide. Nicholas Gotelli (1991) published a general introduction to ecological theory, aimed at advanced undergraduates and beginning graduate students. His definition (Gotelli 1991, p. 112) was based purely upon mutually negative effects: ‘Competitive interactions are those in which two species negatively influence each other’s population growth rates and depress each other’s population sizes’. James P. Grover, in his 1997 book ‘Resource Competition’, states on page 1 that ‘Competition refers to mutually negative interactions among two or more individuals or populations’. If it is between populations, Grover specifies that ‘population growth rate’ is the quantity that is negatively affected. This is clearly an effect-based definition using population density as the perturbation. Nevertheless, his book is restricted to what he considers to be a subset of competitive interactions in which mutual use of limiting resources is the mechanism producing the mutually negative effects. Gurney and Nisbet's (1998) theory text lacks an explicit definition of competition, but only represents the interaction using consumer-resource models. The examples they consider did not exhibit positive effects. T. J. Case's (2000) theoretical ecology textbook stated that competition occurs when ‘individuals of one species suffer a reduction in growth rate from a second species due to their shared used of limiting resources... or [due to] interference’. Case did not specify the time frame over which this reduction in population growth rate is to be measured. Keddy (2000, p. 5) defined competition as ‘the negative effects that one organism has upon another by consuming, or controlling access to, a resource that is limited in availability’ This combination of effect sign and mechanism again neglects the time frame over which the negative effect is to be measured, as well as not specifying the range of properties that could be used to define the negative effect.
Textbooks from the current millennium have mostly adopted a definition that includes both a requirement that resources be involved and a requirement that mutually negative effects occur, although the operational definition of a ‘negative effect' is usually not provided. For example, the 5th edition of C.J. Krebs' textbook (2001, p. 179) defines competition as a situation in which ‘two species use the same limited resource or seek that resource, to the detriment of both', a definition that requires both a resource-based mechanism and a mutually detrimental outcome. Another widely used textbook from the same period, Ricklefs and Miller (2000, p. 384, Table 20-1) suggests that interaction types are classified by the ‘effect of' one species on another. Competition is a case when ‘interactions between individuals... have negative consequences for all species involved’ However, the chapter on competition theory in this book defines competition (p. 403) as ‘use or defence of a resource by one individual that reduces the availability of that resource to other individuals'. Thus, both types of definition are supported, but the mechanistic one seems to ignore the Levine (1976) scenario with its positive net effects, and is generally vague about the time frame of measurement and about situations with more than one resource. Turchin's (2003) monograph on population dynamics separates indirect and direct interactions, and therefore equates interactions to interference competition. This does make his classification of interactions more logically consistent than that in most treatments. However, because interference is almost always present as a result of exploitative interactions with shared resources, neither can be fully understood without considering the resource-mediated pathways.
Textbook definitions have not been converging towards a consensus during the past decade. The most theoretically oriented general textbook available currently (Pasztor et al. 2016, p. 23) defines competition as ‘individuals sharing one or more regulating factors and thus having a negative effect on each other'. The current best-selling (as of 2019 on Amazon.com) ecology text is Bowman et al. (2017). Their definition (p. 318) is, ‘a nontrophic interaction between individuals of two or more species in which all species are negatively affected by their shared use of a resource that limits their ability to grow, reproduce, or survive'. This again combines mechanism and effect, while adding the somewhat unclear requirement of being
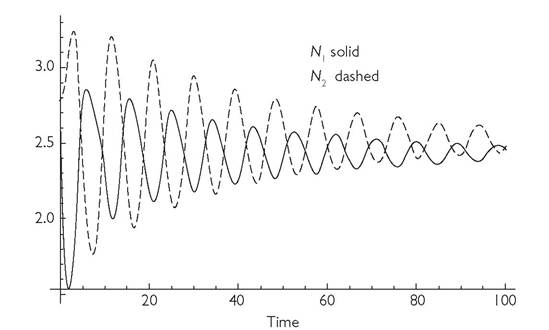
Fig. 2.1 Dynamics of two consumers in a MacArthur system (eqs 3.4), modified by having competition between the two resources. 'The competitors have opposite relative specializations on the resources, but are otherwise equivalent and the simulation is initiated at their equilibrium densities. The start of the simulation corresponds to an imposition of a 50% increase in the death rate of consumer species 1, from 1.0 to 1.5. The initial consumer abundances are both 2.784. The system will eventually come to an equilibrium in which N1 has been reduced from 2.784 to 2.435, while N2 has been reduced from 2.784 to 2.524. The basic parameter values are: bij = 0.125 for all i, j; di = 1; C11 = C22 = 0.4; C12 = C21 = 0.1; ri = 2; Ki = 0.02; α12 = α21 = 0.9 for both resources.
non-trophic, and apparently implying a single resource. (The latter requirement was clearly not meant, given the subsequent examples provided.) The only theory referred to by Bowman et al. (2017) is the LV model, which lacks resources. A somewhat more advanced recent textbook on community ecology (Mittelbach and McGill 2019) cites the works by Grover (1997) and Case (2000), discussed in the previous paragraph, for possible definitions.
In summary, most relatively recent definitions of competition mention resources and they usually require negative effects, although the nature and timing of these negative effects is often not specified. Timing is important because there is often an oscillatory change in the sign of the change in both population growth rate and population size of a second consumer species following the addition or subtraction of individuals of the first consumer (in a two-consumer-species system). For example, Figure 2.1 shows the changes in abundance over time following an instantaneous 50% increase in the death rate of individuals of consumer 1 at time 0. The system represented here is the system described by Levine (1976). This is based on the MacArthur model (eqs (3.4) in the next chapter) modified by having the resources themselves exhibit LV competition in the absence of the consumer. The initial dynamics seem to exhibit competition by many definitions—one species goes up when the other goes down. However, the end result is that harming consumer 1 reduces the abundance of consumer 2, reflecting a positive interspecific effect of 1 on 2.
The continued and widespread use of competition models that lack resources certainly detracts from research into the role of resource dynamics in shaping competition. The most-cited article on competition between species published during the past five years (Levine et al. 2017) mainly considers models that lack resources. (Note that this is not the Levine of the 1976 work referred to above.) Other highly cited works published in recent years have also concentrated on models of competition that lackresources (Saavedra et al. 2017; Mayfield and Stouffer 2017; Hart et al. 2018). One might be able to justify this in cases in which resource dynamics were very fast relative to consumer dynamics, and those resource dynamics were used to derive the form of the consumer-only model. Yet in most cases, this has not been done. O’Dwyer (2018) is a recent discussion of some of the limitations of representing within-and between-species competition for resources based on models without explicit resource dynamics. Lack of resources is particularly problematic when population sizes vary continually.
2.3
More on the topic Historical definitions of competition:
- Arguments against resource-based definitions and models
- Abrams Peter A.. Competition Theory in Ecology. Oxford University Press,2022. — 336 p., 2022
- What should the definition be?
- Competition is often asymmetrical
- Implications of the definition
- Intraspecific competition
- SELECT DEFINITIONS AND CONCEPTS
- Concept and content of state functions.
- Why the Lotka-Volterra and MacArthur models are insufficient
- Important aspects of consumer-resource relationships