SELECT DEFINITIONS AND CONCEPTS
Prior to detailed discussions of wildlife disease topics, it is useful to consider some basic definitions.
Health and Disease
Two approaches commonly have been used to provide a conceptual base for health and disease in wildlife (Wylie 1970).
In one, health is viewed as a concept analogous to temperature. As such, while there may be lower limits (death or absolute zero, respectively), there is no true upper limit for either health or temperature.In such an analogy, health is the concept to be understood and measured, and one seeks to assess how far from death an organism may be. Among humans, features used to assess health can entail physical, mental, emotional, and spiritual components. Considering just physical features, states of human health have been assessed by body mass index, body fat, erythrocyte and serum enzyme measures, and urine metabolites (Berkow and Fletcher 1992). For wildlife and other nonhuman animals, far less is understood regarding what normal physiological values constitute physical health. Measures used also may include body weights, fat indices, as well as a variety of red blood cell, serum enzyme, and urine metabolite values (Malpas 1977, Warren and Kirkpatrick 1978, Seal et al. 1981, DelGuidice et al. 1990, Harder and Kirkpatrick 1994).
In contrast to viewing health on a purported linear scale, health also may be assessed as adaptability to change (Wylie 1970). Less important here is the number of parasites or problems an animal or population encounters than its response and adaptability to these stresses. One working definition of human health is the capacity to achieve socially determined goals (mental, physical, and social well-being, vigor, resilience, productivity, flourishing) within a set of socioecological constraints, only one of which is disease (Murray et al. 2002). Munson and Karesh (2002) view disease as any disorder of body functions, systems, or organs; such a disorder is not necessarily confined to one caused by an extrinsic factor such as viral or bacterial infection, infestation with parasites, or exposure to toxins (Munson and Karesh 2002).
In a very broad outlook, disease has been defined simply as a reduction in fitness (Clayton and Moore 1997).For purposes of this text, we use the definition that wildlife disease is “any impairment that interferes with or modifies the performance of normal functions, including responses to environmental factors such as nutrition, toxicants, and climate; infectious agents; inherent or congenital defects, or combinations of these factors” (Wobeser 1981). Such a definition can be applied to both individuals and populations. Our primary emphasis in this text is on infectious and, secondarily, noninfectious causes of disease.
Parasitism
Parasitism is a term used to describe one set of interspecific relationships along a broad continuum of relationships between species (Dindal 1975). Broadly, the term symbiology (literally, “living together”) has been used to describe the study of any persistent relationship (symbiosis) between two different species; parasitism is one part of that larger set of relationships (Read 1970). It sometimes is argued that while parasites may elude clear definition, they generally are “known primarily when we see them” (Moore 2002).
Broadly, parasites are organisms living partly or completely at the expense of another organism (its host). Parasitic lifestyles are regularly represented among four of the major kingdoms: Animalia, Fungi, Protista, and Monera (with a much less common occurrence among Plantae). Viruses raise an interesting problem; they have some features of living systems (e.g., genome, replication, evolution), but are not functionally active outside their host cells and generally are not considered to be living microorganisms (van Regenmortel and Mahy 2004); however, they commonly are treated as highly specialized parasites and included with other microorganisms in this text. Prions, infectious proteins lacking nucleic acids, also raise interesting problems and are addressed as a special topic in Chapter 11.
Parasitic relationships can overlap with some forms of predation (e.g., parasitoid wasps and some fly maggots that kill their hosts) as well as certain mutualistic relationships (e.g., many intestinal bacteria).
Generally, parasites live in relatively long contact with the host for part of their life cycle; in contrast, predators and prey generally have a relatively short period of contact that ends in death and consumption of the prey by the predators. Further, parasites typically relinquish the role of regulating their relationship with the external environment to the host during part of their life cycle.Endoparasites live within a host's body during at least part of their life cycle and depend completely on the host to regulate their relationship with the external environment during that time; examples of endoparasites include some helminths, fungi, protozoa, and many infectious microorganisms. Parasites living on the exterior of the host are termed ectoparasites; these have a partial dependence on the host to regulate their relationship with the external environment when they are present on the host; examples include parasitic arthropods such as ticks, fleas, lice, and mites. Finally, parasites generally are characterized as competing with the host for its resources, and often reduce host fitness (e.g., survival, fecundity) (Clayton and Moore 1997).
Many parasites are transmitted directly between susceptible hosts; these species have direct life cycles and are termed “monoxenous” (Gr. mono = one, xenous = host). However, others, including many parasitic helminths and protozoa, require two or more hosts to complete their life cycle; these have indirect life cycles and are termed “heteroxenous.” For parasites with an indirect life cycle, the host in which the sexually mature stage of the parasite occurs is called the definitive host. An intermediate host is an additional required host for those parasites to complete their life cycle. In the intermediate host, the parasites undergo some developmental changes and may multiply, but do not reach their sexually mature stage; the intermediate host typically is a different taxonomic group from the definitive host. A less common type of host is a paratenic (transport) host, an organism which serves to transfer a larval stage or stages from one host to another but in which little or no development takes place (Anderson 1992).
A paratenic host is not required for completion of the life cycle, but often is a prey species of the definitive host and facilitates completion of the life cycle; a paratenic host often is considered an “optional” host. The term vector is defined by some as any host that transmits parasites, including intermediate, definitive, and paratenic hosts (Clayton and Moore 1997); in contrast, other scholars tend to restrict the term primarily or exclusively to arthropods (Wobeser 2006).The persistent presence of a parasite in a host is termed an infection (Pratt 1963); a related term, infestation, is used to describe the persistent presence of ectoparasites. Prevalence is the number of animals infected by a parasite divided by the number of animals in the population examined, and commonly is reported as a percent value (Margolis et al. 1982, Bush et al. 1997). In contrast, incidence is the number of new hosts that become infected with a particular parasite during a specified time interval, divided by the number of uninfected hosts present at the start of that time interval (Margolis et al. 1982, Bush et al. 1997); incidence often is reported as number per 1,000 in the population. Intensity is the number of parasites of a particular species in or on a single infected animal; mean intensity is the total number of parasites of a particular species found in each host, divided by the number of hosts infected with that parasite (Margolis et al. 1982, Bush et al. 1997).
Mortality refers to the death of a host and morbidity is the condition of having an illness, weakness, or other disability. In contrast, necrosis refers to a localized area of death in a tissue or organ rather than the death of a whole organism.
Virulence is a measure of the impact of parasites upon their host. Broadly, virulence can be defined as any collective effects on host fitness, including mortality, morbidity, and reduced fecundity (Clayton and Moore 1997). Virulence also has been described more precisely as a complex property comprising three characteristics: infectivity, invasiveness, and pathogenicity (Frobisher 1962, Pratt 1963).
This latter definition has been applied primarily to microorganisms and is considered here in further detail. Infectivity is defined as the ability to initiate and maintain an infection in the host (Pratt 1963). This trait is dependent on the capacity of a parasite to establish a persistent presence by evading or overcoming local defense mechanisms of the host. For example, bacteria causing plague (Yersinia pestis), typhoid fever (Salmonella typhi), or shigellosis (Shigella spp.) do not have high infectivity to laboratory workers under normal circumstances (Pratt 1963). In contrast, Francisella tularensis, the cause of tularemia, is a highly infectious bacterium readily transferred to workers within the laboratory (Pratt 1963, Hopla and Hopla 1994).
Invasiveness is defined as the ability to progress further into the host from the initial site of infection (Pratt 1963). For example, many bacteria, including Pasteurella multocida, the cause of avian cholera, commonly invade the blood stream, causing a septicemia (presence of pathogenic bacteria or their toxins in the blood) among infected wildfowl.
Pathogenicity is the ability to injure a host (Pratt 1963) by damaging host tissues. For larger parasites such as helminths and arthropods,
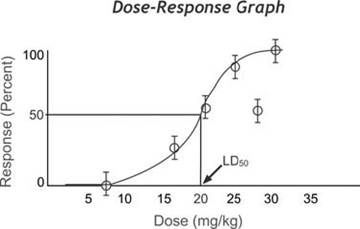
FIGURE 1.4 Sigmoid curve typical of an LD test (Courtesy of National Library of Medicine. Based on original from http://aquaticpath.umd. edu/appliedtox/images=toxtutor/ chart-5.gif. Drawn by Patient Education Institute).
physical damage and blood loss are common causes of morbidity and mortality. Among bacteria, toxins affecting the nervous system, heart, or kidney are more typical causes of pathogenicity (Pratt 1963). In the laboratory, pathogenicity in living animals commonly has been measured by an LD test; an LD is the dose of an infective agent or toxin lethal for 5°% of a test population (Fig. 1.4) (Reed and Muench 1938).
The LD test has been highly regarded as a measure of pathogenicity because of its consistency and its requirement for relatively few test animals. However, the LD test also 5° is limited to assessing mortality and cannot assess more subtle influences such as morbidity, reduced reproductive success, or increased susceptibility to predation; further, it also is useful only for single, isolated mortality factors and is not readily adapted to assessing syner- gystic relationships between two or more factors influencing mortality. In recent years, the LD (and the consequent killing of laboratory animals) increasingly has been replaced by alternative tests to measure pathogenicity, such as assessment of cellular pathology through the use of tissue cultures. However, it still is used in toxicology, where every registered pesticide must have at least three avian LD tests 5° (A. Fairbrother, pers. comm.).
In summary, for a microorganism to be considered virulent, it must simultaneously be infective, invasive, and pathogenic for a given host (Pratt 1963). For example, Mycobacterium tuberculosis, a cause of human tuberculosis, is very invasive and highly pathogenic to guinea pigs (Cavia porcellus) in the laboratory (Wamoscher and Stocklin 1927, Dorr and Gold 1932, Wilson and Miles 1964); yet there are no evident reports of its occurrence among guinea pigs in their natural habitats (Shope 1927, Wilson and Miles 1964, Williams 2001). Thus, while very invasive and pathogenic, an apparent absence of infectivity would result in the bacteria not being considered virulent for guinea pigs. Likewise, although Pasteurella multocida, the cause of avian cholera, is virulent to at least 180 species of birds (Samuel et al. 2007), there is no evidence that P. multocida can invade the blood stream (cause a septicemia) or cause a clinical disease among turkey vultures (Cathartes aura) (Botzler 1991, Samuel et al. 2007); thus by definition P. multocida is not invasive, and consequently not virulent, for turkey vultures.
Historically, microparasite virulence was viewed as a sign of recent association between a host and parasite, and it was argued that subsequent host-parasite co-evolution would lead to a reduction of virulence and even the development of commensalism or mutualism (Burnet and White 1972). A more recent hypothesis is that virulence also can be maintained by natural selection and may increase or decrease in evolutionary response to environmental conditions or the density and behavior of hosts (Levin 1996). Thus the level of virulence expressed by parasites may result from the strategy developed by the infective agent for optimal transmission and survival (Ewald 1994). Alternatively, it also has been proposed that the virulence of microparasites is coincidental to parasite-expressed characters that evolved for other functions, or emerged as the product of short-sighted evolution in infected hosts (Levin 1996). All of these factors may play a role in different circumstances.
Diseases in Populations
Several key terms are used in describing diseases in populations. The term epidemic (epi: upon; dem: people) refers to a disease affecting many people within an area at one time, at a significantly greater occurrence than expected. Examples include outbreaks of bubonic plague and human influenza. The term epizootic has been used to refer to epidemics within nonhuman animals and the term epornitic occasionally is used to refer to epidemics among avian populations. The term pandemic refers to a worldwide epidemic (among humans, since most other species don't have the same broad distribution). For example, human influenza is estimated to have caused the death of >20 million humans worldwide during the pandemic of 1918-19 (Slemons and Brugh 1994). Bubonic plague also has caused numerous pandemics among humans (McNeill 1977).
In contrast, the term endemic refers to a parasite or disease with a low incidence, but one that is regularly present in a host population. Enzootic is a similar term that has been used in reference to diseases characteristic of nonhumans. For example, Yersinia pestis, the cause of bubonic plague, is maintained among some rodent populations such as the California vole (Microtus californicus) and the deer mouse (Peromyscus maniculatus) in western North America; Y. pestis is considered enzootic in these species (Poland et al. 1994). One potentially confusing aspect of the term endemic is that in ecological literature it commonly refers to a species that evolved solely in a limited area or region, as on certain islands (Van Dyke 2003); use of the alternative term “enzootic” in disease literature can help reduce that potential confusion.
The terms endemic and enzootic also have been used to signify a parasite or disease characteristic of a geographic region (rather than a particular host species). Thus, plague also can be characterized as enzootic to dry grasslands, mountain meadows, and some deserts of western North American and other regions of the world. As another example, avian cholera, caused by Pasteurella multocida, regularly causes epornitics among wildfowl of North America; while found in all North American flyways, it generally is considered enzootic to northern California, Nebraska’s Rainwater Basin, and the Texas playa lakes (Friend 1999).
The term reservoir has been used by authors in several ways. Simply stated, it can be defined as the sum of all sources of infection—the natural habitat of the parasite (Pratt 1963). More specifically, a reservoir of infection has been defined as an ecologic system in which an infectious agent survives indefinitely (Ashford 1997); such an ecologic system would encompass all of the vertebrate and invertebrate host populations and encompass the pertinent environmental factors as well as any quantitative factors, such as critical community size, needed to maintain an infectious agent indefinitely (Ashford 2003). For our use, we generally refer to Ashford’s modified 2003 definition of a reservoir.
Examples of reservoirs range from red foxes (Vulpes vulpes) for rabies viruses in Europe (Rupprecht et al. 2001), to soil, mud, or water as reservoirs for the bacterium Listeria monocytogenes (Bille et al. 1999). However, the notion of reservoir for a parasite or disease may vary with geographic scale, such as that occurring within a specific watershed versus a more general assessment of the reservoir on a worldwide basis. Further, in describing reservoirs, researchers only address a limited number of aspects of the environment and thus may miss key criteria in their descriptions. The term reservoir species has been used in a specialized sense to refer to an introduced host that has artificially raised the size of the collective host populations or densities, consequently allowing pathogen transmission even when the endemic host population had been reduced below the density at which a pathogen is able to maintain effective transmission (Daszak and Cunningham 2002).
Time scale is another important factor. Arthropod-borne agents of vertebrates that survive in temperate regions often survive during a period of time (e.g., cold, dry) when arthropod survival is low. In some cases the infective agent survives primarily in the vertebrate population and the vertebrates could be considered the primary reservoir. However, our use of
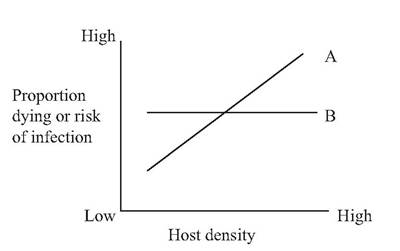
FIGURE 1.5 Contrast of density dependent (A) and density-independent (B) disease impacts.
reservoir (e.g., in Chapters 9 and 10) will generally take an annual or multi-year perspective, and thus we will refer to arthropod-vertebrate reservoirs in these chapters, even where some members (e.g., arthropods) play a smaller role during some parts of the year.
Density and Disease
Parasites and diseases whose risk of infection or impact varies consistently with the density (number per unit area) of their host populations are termed density-dependent, whereas those whose risk or impact do not change in response to differing densities are called densityindependent. Thus, for a density-dependent disease, the rate of transmission of a disease through a population, or risk of infection for the susceptible individual, varies consistently with the density of the host population (Fig. 1.5). Generally, parasites transmitted directly between hosts of a susceptible species, without the need for intermediate hosts or vectors, cause densitydependent diseases (Scott 1988).
Rabies often is cited as a classical densitydependent disease (Macdonald 1980, Bacon 1985). One would expect that a rabid fox would have a higher probability of encountering an uninfected, susceptible fox in a high-density fox population, compared to a low-density fox population. Thus the risk of rabies transmission would be higher in a high-density than in a low-density population of susceptible animals.
In contrast, among density-independent diseases, the rate of transmission of the causative agent through a population, or risk of infection for the host, is independent of the density of the susceptible host population. Pesticides and environmental toxins, infectious diseases with reservoirs in soil or water, severe weather, and accidents tend to have density-independent impacts. For example, the risk to an individual bird of flying into a power line during a migration would not be expected to change in a consistent fashion if the flock size increased or decreased. Also, one of the devastating impacts of some pesticides is that their lethal effects are unabated even as the host populations reach very low levels (Hickey and Anderson 1968, Risebrough 1978, Peek 1986).
It is important to note that the significant distinguishing feature between densitydependent and density-independent diseases is the proportion (rather than the actual number) of a susceptible population affected. For example, if the hosts in a susceptible population have a 5% risk of mortality (in a density-independent situation), one would expect approximately 5 hosts in a population 100, or approximately 50 hosts in a population 1,000, to die. Thus, while the total numbers of animals dying increases in larger populations, the actual proportion of animals affected (5%) is unchanged when transmission is density independent.
Density dependence becomes more complicated with parasites undergoing indirect life cycles. For example, among parasitic helminths, the intestinal tapeworm Echinococcus granulosus involves large canids (e.g., wolves, Canis lupus; coyotes, Canis latrans) as definitive hosts and ungulates (e.g., moose, Alces alces; deer, Odocoileus spp.) as intermediate hosts. For a given season, the risk of infection to wolves by ingesting the tapeworm infective stage (hydatid cysts) is more directly dependent on the density of hydatid-infected moose than on the density of the other infected wolves (who are shedding eggs infective to moose). Likewise, in a given season the risk of moose becoming parasitized by ingesting tapeworm eggs is more directly affected by the density of wolves shedding eggs than by the density of other hydatid-infected moose. Thus, over a shorter term, such as a single season, risks of acquiring indirect life cycle parasites tend to be density independent. However, over several seasons, one would expect that increases in wolves (shedding eggs) would lead to increases in infected moose, which, in turn, would lead to increases in the prevalence of infection among the wolves (eating infected moose); thus, over a longer term, risks of acquiring indirect life cycle parasites can become density dependent.
There is some similarity for arthropod- borne parasites (e.g., West Nile virus) to the relationship described above for indirect life cycle parasites. However, arthropod-borne parasite life cycles are complicated by additional factors such as the repeated feeding by some arthropods in a season, with their consequent increased likelihood of acquiring parasites. In such cases, the risk of infection to a susceptible vertebrate host is potentially dependent on both the density of infected vectors and the density of the other infected vertebrates. Likewise, the risk of infection to a vector is certainly influenced by the density of infected vertebrates, but also can be influenced by the density of other infected vectors that may infect vertebrates and make them available later in a season to the uninfected vectors. It is more difficult to break the cycle in this case as both vertebrate hosts and vectors can contribute to a condition more closely representing that of density dependence. While over a shorter term, such as a single season, arthropod-borne diseases often tend to be density independent, the influences of multiple feeding each season may lead to an additional density-dependent influence as well, over several seasons.
Disease Models
We present two models to use as foundations for conceptualizing key relationships in the field of wildlife diseases. One entails
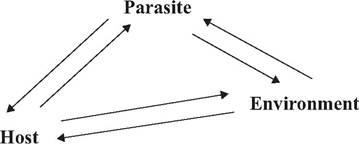
FIGURE 1.6 Parasites, hosts, and their environments are closely intertwined—each affecting the other two in varying degrees.
an understanding of three general components: parasites and other causative agents of diseases, the affected vertebrate and invertebrate hosts, and the environment in which these occur. In this model each component interacts with the other two, and must be described and understood in relation to the other two (Fig. ι.6). Any change in one can drastically alter the balance of a resulting disease.
An earlier and more complex disease model, proposed in 1939 by Soviet biologist Evgeny N. Pavlovsky, first was used to describe arthropod-borne diseases (Pavlovski 1966). It variously has been termed the Natural Nidality Doctrine of Transmissible Diseases, the Landscape Theory of Epidemiology, the Landscape Theory ofZoonotic Diseases (Pollitzer and Meyer 1961, Pavlovski 1966), and the Natural Nidality Theory (Nelson 1980).
Pavlovski believed that most transmissible diseases exist in nature as discrete foci or nidi (sing. nidus, “hearth,” “home”). A nidus is defined as that portion of a region with a definite geographic character, and would be similar in usage to that of the terms biotope, ecosystem, or habitat type. A nidus can be a small local area or a broad geographic region (Pavlovski 1966). Thus, rabies in the range of infected red foxes (Vulpes vulpes) in Europe would be an example of a broad nidus; likewise, plague would be seen as having nidi among dry grasslands (steppes), mountain meadows, and some desert habitats in western North America and Asia. A nidus also can be dynamic, as in the cases of a rabies nidus among red foxes moving across Europe (Macdonald 1980), or a plague nidus shifting in North America (Barnes 1982). However, most nidi are more permanently associated with specific regions and habitat types.
Within each nidus is an ecological association termed a biocoenosis, composed of the infective agent, the wild vertebrate reservoir species, intermediate hosts or vectors (typically arthropods or other invertebrates), and any wild vertebrate amplifying hosts. All of these organisms are limited in their geographic and ecological distribution by the environmental determinants of the habitat. These biocoenoses allow circulation of the parasites indefinitely and can be viewed as integrated wholes (Pavlovski 1966). Microscale disease foci are influenced by qualities of the entire ecosystem. Focal diseases in discrete sites may spread out from the nidi to cause epizootics among susceptible hosts. Susceptible wildlife, humans, or other domestic animals might become involved if they invade an active nidus. This holistic approach to the study of disease ecology has been used as a foundation for the study of plague in North America (Nelson 1980) as well as a number of other diseases (Pavlovski 1966).
More on the topic SELECT DEFINITIONS AND CONCEPTS:
- If definitions of ‘violence' are slippery, the concepts of crime and law offer some purchase.
- Select Bibliography
- “The Masses” versus “the Select”
- A Study of Select Ecoethical Narratives from Northeast India
- Three Classic Definitions
- Two Prominent Definitions
- Definitions
- Definitions
- AIDS case definitions and staging of HIV disease
- Why do definitions matter?
- 3. Models and definitions
- PROBLEMS OF DEFINITIONS