Human actions have caused communities to shift to alternative states
The stringent requirements suggested by Connell and Sousa had the effect of delaying alternative stable state research for two decades. Recently, however, there has been renewed interest in alternative stable states, spurred by the increasing evidence that human activities, such as habitat destruction, species introductions, and overharvesting of wild species, are shifting communities to alternative states.
We have already seen examples of such changes in several of the Case Studies in this book, including the change from kelp forests to urchin barrens due to the decline of sea otters (see Chapter 9), the crash of the anchovy fishery in the Black Sea due to the introduction of Mnemiopsis (see Chapter 10), and the invasion of the aquarium strain of Caulerpa taxifolia in the Mediterranean, Australia, Japan, and North America (see Chapter 16). These so- called regime shifts are caused by the removal or addition of strongly interacting species that maintain one community type over others. Ecologists are uncertain whether the results can be reversed or whether hysteresis will occur once communities have been “manipulated” by human activities and new regimes are in place. Will recolonization by sea otters rejuvenate kelp forests? Will the cessation of nutrient enrichment in the Black Sea revitalize the anchovy fishery? And will the removal of Caulerpa restore seagrass communities? These are all questions whose answers may be found in a better understanding of the factors that drive alternative stable states and of the role restoration of the original conditions can play in reversing the effects of those factors.A Case Study Revisited
A Natural Experiment of Mountainous Proportions
On the twentieth anniversary of the eruption of Mount St. Helens, in 2000, a group of ecologists gathered on the once smoking and ash- covered volcano to participate in a week-long field camp.
They gathered their gear, including tape measures, quadrat frames, and maps, and visited the same sites they had explored two decades earlier. This visit, termed a “pulse,” was an opportunity to establish a 20-year benchmark of data comparable to those first collected in1980 and 1981. Many of the participants had spent the past 20 years—for some, their entire careers—studying recolonization and succession patterns in those once-devastated landscapes. When they departed, they agreed to write a book, the chapters of which would contain all that was known about the extraordinary ecology of this ecosystem, with the hope that young ecologists would be motivated to continue the research and carry on their legacy. The book, Ecological Responses to the 1980 Eruption of Mount St. Helens (Dale et al. 2005), was published 5 years later.
What does the book tell us about succession on Mount St.
Helens? First, the eruption created disturbances that varied in their effects depending on distance from the volcano and habitat type (e.g., aquatic vs. terrestrial). Although areas close to the summit, such as the Pumice Plain, were literally sterilized by the heat of the eruption, ecologists were surprised to discover how many species actually survived on the mountain (TABLE 17.2). Because the eruption occurred in spring, many species had been still dormant under the winter snows. Survivors included plants with underground buds or rhizomes, animals such as rodents and insects with burrows, and fish and other aquatic species in ice-covered lakes. In the blowdown zone, large trees and animals perished while smaller organisms survived in the protection of their larger neighbors. The opposite was true in areas outside the blowdown zone, where falling rocks and ash smothered smaller plants and animals, but not larger organisms.
TABLE 17.2 Surviving Organisms Found on Mount St. Helens within a Few Years after the Eruption
Disturbance zone
Mean vegetation
Average number of
Animals
| cover (%) plant species/m2 | Small mammals | Large mammals | bgcolor=white>BirdsLake fish | A | |||
| Pumice Plain | 0.0 | 0.0 | 0 | 0 | 0 | 0 | 0 |
| Mudflow zone | 0.0 | 0.0 | 0 | 0 | 0 | N/A | 0 |
| Blowdown zone | 8 | 0 | 0 | 4 | 1 | ||
| Preeruption clear-cut | 3.8 | 0.0050 | |||||
| Forest without snow | 0.06 | 0.0021 | |||||
| Forest with snow | 3.3 | 0.0064 | |||||
| Scorch zone | 0.4 | 0.0039 | 0 | 0 | 0 | 2 | 1 |
Source: C.
M. Crisafulli et al. 2005. In Ecological Responses to the 1980 Eruption of Mount St. Helens, V. H. Dale et al. (Eds.), pp. 287-299. Springer: New York, based on references cited there.
A second important research discovery from Mount St. Helens is the role survivors have played in controlling the pace and pattern of succession. In many cases, these species were thrust into novel physical environments and species assemblages without time to adapt over evolutionary time scales. Some species thrived, while others fared poorly, but their adaptability and unpredictability were surprising. Unlikely alliances were formed that hastened succession in particular habitats. For example, newly formed and isolated ponds and lakes were colonized by amphibians much faster than had been thought possible (FIGURE 17.19). Scientists discovered that frogs and salamanders were using tunnels created by northern pocket gophers (Thomomys talpoides) to make their way from one pond to another across the arid landscape (Crisafulli et al. 2005). The gophers were particularly successful on Mount St. Helens, both because they survived the eruption in their tunnels and because grassy meadows—their preferred habitat—expanded greatly after the eruption. Interestingly, the gophers were also responsible for facilitating plant succession: their burrowing activity brought to the soil surface organic matter, seeds, and fungal spores buried deep under the volcanic rock and ash (Crisafulli et al. 2005) (FIGURE 17.20).
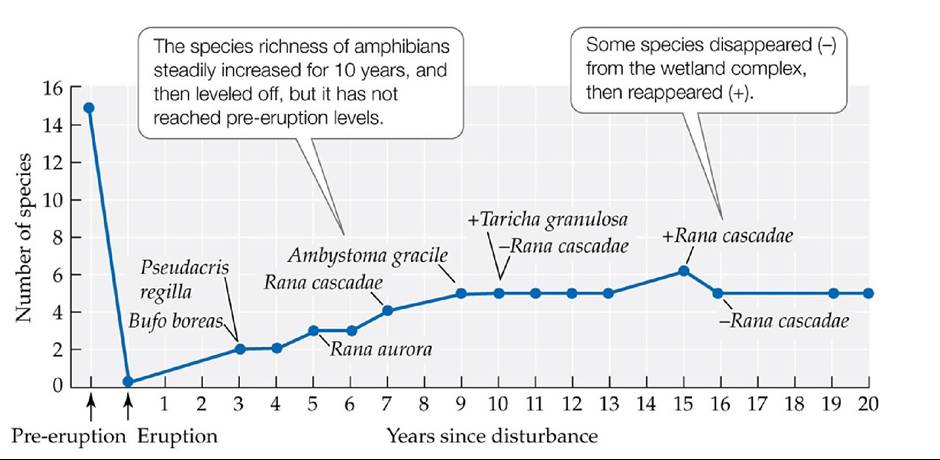
FIGURE 17.19 RapidAmphibianColonization Frog and salamander species rapidly colonized a wetland complex in the Pumice Plain on Mount St. Helens. (After C. M. Crisafulli et al. 2005. In Ecological Responses to the 1980 Eruption of Mount St. Helens, V. H. Dale et al. [Eds.], pp. 183-197. Springer: New York.) View larger image

FIGURE 17.20 PocketGopherstotheRescue The burrowing activity of northern pocket gophers, some of which survived the eruption underground, brought organic matter, seeds, and fungal spores to the soil surface, creating microhabitats, like this one in the Pumice Plain, where plants could grow.
View larger imageA third important discovery was the realization that multiple mechanisms were responsible for primary succession on Mount St. Helens. Facilitation on the Pumice Plain was exemplified by the dwarf lupine (Lupinus lepidus), the first plant to arrive there. Dwarf lupines trapped seeds and detritus and increased the nitrogen content of the soil though their symbiotic association with nitrogenfixing bacteria (del Moral et al. 2005). The lupines, in turn, were inhibited by multiple insect herbivores, which essentially controlled the pace of primary succession (Bishop et al. 2005). Tolerance was evident in some primary successional habitats, where Douglas fir lived in concert with annual herbs. The diversity of strategies species used, and the resulting community compositions, never ceased to amaze ecologists, who up to that point had been guided mostly by the models of Connell and Slatyer (1977).
Despite decades of data and a treasure trove of novel discoveries, research on Mount St. Helens has only just begun. Will communities there follow paths of succession that lead to predictable and repeatable outcomes? Or will they form alternative states that are highly dependent on their historical legacies? Geologic studies suggest that Mount St. Helens erupts roughly every 300 years. The life span of its community succession thus greatly exceeds our own life span by hundreds of years, so we must be content with the limited knowledge we have gained from studying what is arguably the most interesting phase of succession on Mount St. Helens and with the hope that ecologists will continue their research there for years to come.
Connections in Nature
Primary Succession and Mutualism
We saw in Chapter 15 that positive relationships can alter communities, and that they may be particularly important in stressful environments. Primary succession in terrestrial environments illustrates both of these effects: some of the examples presented in this chapter involve plants that interact in a mutualistic way with symbiotic nitrogen-fixing bacteria.
These bacteria form nodules in the roots of their plant hosts, where they convert nitrogen gas from the atmosphere (N2) into a form that is usable by plants (ammonia [NH4+]). The plants provide the bacteria with sugars produced by photosynthesis. This interaction appears to be extremely important for plants and animals colonizing completely sterile environments. We have seen that Dryas and alders, both species that form tight mutualisms with nitrogen-fixing bacteria, were some of the first species to colonize the till left behind by glaciers at Glacier Bay,Alaska. Similarly, Lupinus Iepidus was able to use the nitrogen produced by its bacterial symbionts to colonize the sterile Pumice Plain of Mount St. Helens after the eruption. Lupines were the major source of nitrogen for subsequent plants and herbivorous insects for many years. Thus, lupines and their symbiotic bacteria play a large role in controlling the rate of primary succession on Mount St. Helens.
The nitrogen-fixing bacteria involved in symbioses are extremely diverse. Only a few groups of bacteria live in root nodules; all the rest are associated with either the surfaces of roots or the guts of ruminants. The nodule-forming bacteria include the rhizobia, a taxonomic group associated with legumes (such as lupines), and actinomycetes of the genus Frankia, which are associated with woody plants such as alders and Dryas. Nodule formation involves a complex series of chemical and cellular interactions between the root and the bacteria (FIGURE 17.21). Free-living bacteria are attracted to root exudates that cause the microbes to attach to the roots and multiply. Sets of genes are activated in both bacterial and root cells that allow the bacteria to enter the root, the root cells to divide, and the nodule to be formed.
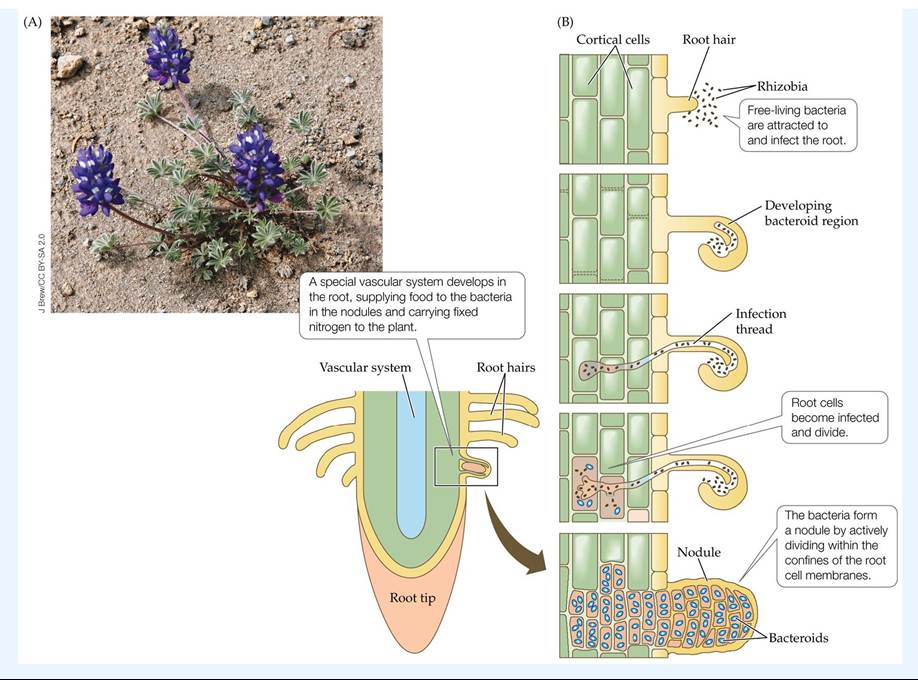
FIGURE 17.21 DwarfLupinesandNitrogen-FixingBacteria (A)Dwarflupine (Lupinus lepidus), a legume with symbiotic nitrogen-fixing bacteria, was the first plant to colonize Mount St.
Helens. (B) Root nodule development is the result of a strong interaction between the plant and the bacteria. View larger imageThe enzyme involved in nitrogen fixing (nitrogenase) is highly sensitive to oxygen and requires anaerobic conditions. Thus, wherever nitrogen-fixing symbioses occur, there are structural components to the interaction (such as membranes within the root nodules) that produce anaerobic conditions. The bacteria, however, need oxygen to metabolize, so a hemoglobin protein known as leghemoglobin, which has a high affinity for oxygen, is produced in the nodules to deliver oxygen to the bacteria in an essentially anaerobic environment. The nodules often have an eerie pink color that is associated with the leghemoglobin. In addition, the nodule develops a specialized vascular system that supplies sugars to the bacteria and carries fixed nitrogen to the plant.
Maintaining a symbiosis with nitrogen-fixing bacteria is costly to plants. Estimates suggest that creating and maintaining the nodules alone costs a plant 12%-25% of its total photosynthetic output. Plants may be able to shoulder this cost, especially if it allows them to live in environments free of competitors and herbivores. But as they increase the nitrogen content of the soils in which they live, plants with symbionts make conditions better for other plant species as well—some of which are likely to be competitors. Thus, these plants face a trade-off between improving the environment for themselves and competing with other species, which makes their role in early successional environments important, if somewhat ironic.