Experiments show facilitation to be important in early stages
A number of experimental studies like the ones we have just described, initially stimulated by Connell and Slatyer's 1977 paper, suggest that succession in any community is driven by a complex array of mechanisms (see ANALYZING DATA 17.1).
No one model fits any one community; instead, each community is characterized by elements of all three of Connell and Slatyer's models. In most successional sequences, especially those in which a pioneer stage is exposed to physically stressful conditions, facilitative interactions are important drivers of early succession. Organisms that can tolerate and modify these physically challenging environments will thrive and facilitate other organisms that lack those capabilities. As succession progresses, slow-growing and long-lived species begin to dominate. Those species tend to be larger and more competitively dominant than early successional species. For this reason, one might expect competition to play a more dominant role than facilitation later in succession.ANALYZING DATA 17.1
What Kinds of Species Interactions Drive Succession in Mountain Forests?
We learned in Concept 17.3 that successional patterns are often the result of complex species interactions. Such interactions are exemplified in a study investigating the patterns of succession in
mountain forests in Utah dominated by quaking aspen (Populus tremuloides) and subalpine fir (Abies Iasiocarpa) (Calder and St. Clair 2012).* In some cases, aspen can form stable and selfsustaining populations, but more commonly these trees occur in mixed stands with firs. Observations show that aspen initiate the earliest stage of secondary succession in open meadows created by fire or deforestation, using root suckers (underground shoots that produce clonal plants; see Figure 9.4) to colonize open meadows. Over time, mixed aspen-fir stands are formed as the shade-tolerant firs become established and increase in abundance while aspen decline.
The stands are eventually dominated by firs, which are more susceptible to fire than pure stands of aspen, thus increasing the chance of starting the successional cycle anew.To understand the transition from one successional stage to another, Calder and St. Clair counted the aspen suckers and fir seedlings in four successional stages—meadow, aspen, mixed aspen-fir, and fir—with the results shown in FIGURE A. To test for the type of interaction important in the transition from one stage to another, the researchers then measured the mortality of canopy aspen and subalpine fir trees as a function of their distance from the nearest neighboring tree of the other species. These results are given in FIGURE B. Error bars show one standard error of the mean.
(Â)
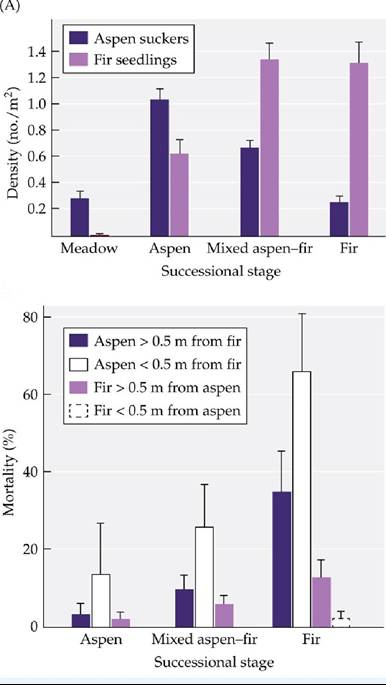
1. Based on the data in Figure A, what is the pattern of aspen abundance over the four successional stages? How does the abundance pattern of subalpine fir differ? Do these patterns of abundance of aspen and fir support the successional sequence described in the first paragraph above?
2. What type of interspecific interaction would you hypothesize could account for the difference between the number of fir seedlings in the aspen stage and in the meadow stage in Figure A? What type of interspecific interaction might explain the difference between the number of aspen suckers in the mixed and the fir stages?
3. Now consider Figure B. What happens to fir trees when they live close (of Styela dominance by suggesting that fish predation was spotty and that the tunicates themselves, once they reached a certain large size, might have acted as a natural “cage” or predator exclusion mechanism.
Lewontin (1969) and Sutherland (1974) both believed that multiple stable states existed in communities and could be driven by the addition or exclusion of particularly strongly interacting species. If those species were missing or ineffective, communities could follow alternative successional trajectories that might never lead back to the original community type (state) but might instead form a new community type.
We can visualize the theory behind alternative stable states by imagining a landscape in which different states are represented by valleys and in which a community is represented by a ball (FIGURE 17.18A). Just as the ball can move from one valley to another, the community can move from one state to another, depending on the presence or absence of strongly interacting species and how they affect the community (FIGURE 17.18B). For example, it may take only a slight change in the abundance of one or more dominant species to force the community (ball) into an alternative state (valley), or it may require complete removal of a species to cause this change. If we use Sutherland's work as an example, we can think of the Styela and Schizoporella community types as two different valleys. Whether the ball resides in the Schizoporella valley or the Styela valley depends on the presence of fish predators. Interestingly, in this system, the ball may not simply move back to the Schizoporella valley if access is restored to fish predators (FIGURE 17.18C). As Sutherland noted, Styela is able to escape predation once it reaches a certain size. Thus, this system might show hysteresis, an inability to shift back to the original community type even when the original conditions are restored.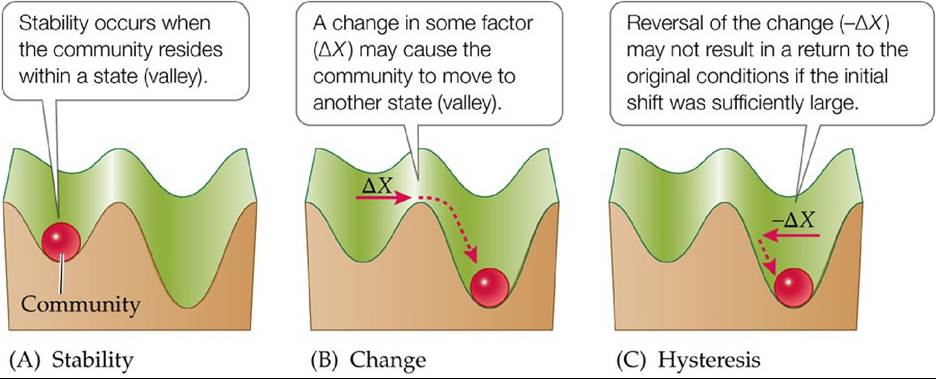
FIGURE 17.18 A Model of Alternative Stable States (A) A community is represented by a
ball that moves within a landscape of community states (valleys). (B) Note that some valleys can be deeper than others, suggesting the magnitude of change (ΔX) needed to shift the community from one state to another. (C) Hysteresis occurs when reversal of the change (-ΔX) does not return the community to its original state. (After B. E. Beisner et al. 2003. FrontEcol Environ 1: 376382.) View larger image
Connell and Sousa (1983) were skeptical that Sutherland had demonstrated the existence of alternative stable states, for several reasons.
First, they thought that his tile communities did not persist long enough, or have a spatial scale large enough, to be considered stable. If the tiles could be followed over multiple years, they asked, would they not all end up being dominated by one or the other species? In addition, they wondered whether the fouling communities could have been sustained outside of an experimental setting in which predators were removed. Their final argument, although it was not a criticism of Sutherland's study per se, was that alternative stable states could be driven only by species interactions and not by physical changes in the community. They argued that many of the examples Sutherland had used to bolster the importance of alternative stable states fell into the latter category. Their requirement that the physical environment not change is problematic because it excludes as drivers of succession all species that interact with other species by modifying their physical environment—that is, all ecosystem engineers. We know that ecosystem engineers can have strong effects on communities, so excluding them is unrealistic to most ecologists.