No one model fits any one community
To illustrate the types of successional mechanisms that have been revealed by experiments, we will focus on three studies: communities that form (1) after glacial retreat in Alaska, (2) after vegetation disturbance in salt marshes in New England, and (3) after wave disturbance in the rocky intertidal zone of the U.S.
Pacific coast.Primary Succession in Glacier Bay, Alaska
One of the best-studied examples of primary succession occurs in Glacier Bay, Alaska, where the melting of glaciers has led to a sequence of community change that reflects succession over many centuries (FIGURE 17.9). Captain George Vancouver first recorded the location of glacial ice there in 1794, while exploring the west coast of North America. Over the last 200 years, the glaciers have retreated up the bay, leaving behind bare, broken rock (known as glacial till). John Muir, in his book Travels in Alaska (1915), first noted how much the glaciers had melted since Vancouver's time. When he visited Glacier Bay in 1879, he camped among ancient tree stumps that had once been covered by ice and saw forests that had grown up in previously glaciated areas. He was impressed with the dynamic nature of the landscape and how the plant community responded to the changes.
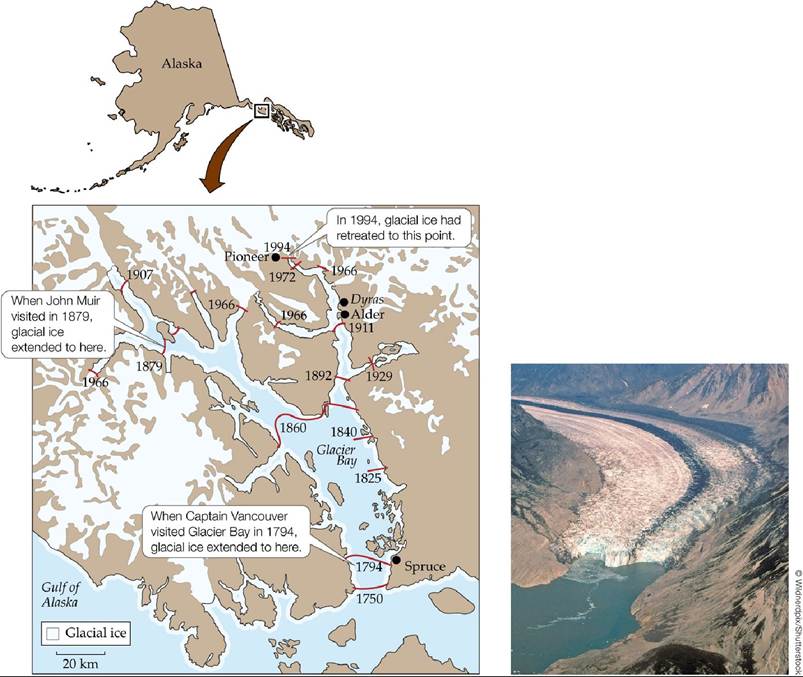
FIGURE 17.9 Glacial Retreat in Glacier Bay, Alaska Over more than 200 years, the melting of glaciers has exposed bare rock to colonization and succession.
Based on the locations of the glaciers over time, describe where the oldest and youngest communities are located.
(After F. S. Chapin et al. 1994. Ecol Monogr 64: 149-175.) View larger image
Muir's book sparked the interest of William S. Cooper (1923a), who began his studies of Glacier Bay in 1915. A former student of Henry Chandler Cowles, Cooper saw Glacier Bay as an example of the “space for time substitution” so well documented by his advisor in the Lake Michigan dunes.
He established permanent plots (Cooper 1923b) that have allowed researchers to observe the pattern of community change along the bay from Vancouver's time to today. This pattern is generally characterized by an increase in plant species richness and a change in plant species composition with time and distance from the melting ice front (FIGURE 17.10). In the first years after new habitat is exposed, a primary or pioneer stage develops, dominated by a few species that include lichens, mosses, horsetails, willows, and cottonwoods. Roughly 30 years after exposure, a second community develops, named the Dryas stage after the small shrub (Dryas drummondii) that dominates this community. In this stage, species richness increases, with willows, cottonwoods, alders (Alnus sinuata), and Sitka spruce (Picea sitchensis) sparsely distributed among the carpet of Dryas. After about 50 years (or some 20 km from the ice front), alders dominate, forming the third community, referred to as the alder stage. Finally, a century after glacial retreat, a mature Sitka spruce forest (the spruce stage) is in place, which fosters a diverse array of lichens, low shrubs, and herbs. Reiners et al. (1971) documented that 200 years after exposure, species richness decreases somewhat as Sitka spruce forests are transformed into forests of longer-living western hemlocks.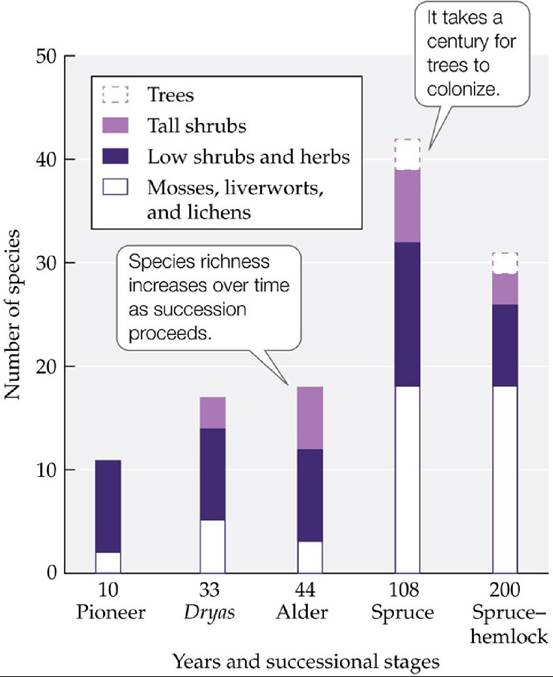
FIGURE 17.10 Successional Communities at Glacier Bay, Alaska Plantspeciesrichness has generally increased over the 200 years following glacial retreat. (After W. A. Reiners et al. 1971. Ecology 52: 55-69.) View larger image
The mechanisms underlying succession in this system have been studied extensively by F. Stuart Chapin and colleagues (1994). They wondered, given the harsh physical conditions experienced by most species in the pioneer stage, whether the facilitation model could explain the pattern of succession observed by Cooper and Reiners et al.
First, they analyzed the soils of the different successional stages. They found significant changes in soil properties that were coincident with the increases in plant species richness (FIGURE 17.11). Not only were there increases in soil organic matter and soil moisture in later stages of succession, but nitrogen increased more than fivefold from the alder stage to the spruce stage. (This increase resulted from the action of nitrogen-fixing bacteria associated with plant roots, which we'll describe in more detail in this chapter's Connections in Nature.) Chapin hypothesized that the assemblage of species at each stage of succession was having effects on the physical environment that largely shaped the pattern of community formation. The question remained, however, whether those effects were facilitative or inhibitory, and how they varied across the different successional stages.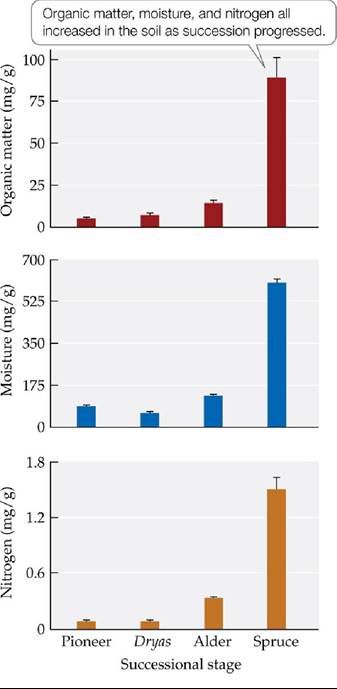
FIGURE 17.11 Soil Properties Change with Succession Chapinandcolleaguesstudied the properties of the soils in each of four successional stages at Glacier Bay. Error bars show one standard error of the mean. (After F. S. Chapin et al. 1994. Ecol Monogr 64: 149-175.) View larger image
To test their facilitation hypothesis, Chapin et al. (1994) conducted manipulative experiments. They added spruce seeds to each of the successional stages and observed their germination, growth, and survival over time. These experiments, along with observations of unmanipulated plots, showed that neighboring plants had both facilitative and inhibitory effects on the spruce seedlings but that the directions and strengths of those effects varied with the stage of succession (FIGURE 17.12). For example, in the pioneer stage, spruce seedlings had a low germination rate, but a higher survival rate, than in later successional stages. In the Dryas stage, spruce seedlings had low germination and survival rates due to increases in seed predators, but those individuals that did survive grew better because of the presence of nitrogen fixed by symbiotic bacteria associated with Dryas.
In the alder stage, a further increase in nitrogen (alders also host nitrogen-fixing bacteria) and an increase in soil organic matter had positive effects on spruce seedlings, but shading and seed predators led to overall low germination and survival rates. In this stage, alders had a net positive effect on spruce seedlings that germinated before alders were able to dominate. Finally, in the spruce stage, the effects of large spruce on spruce seedlings were mostly negative and long-lasting. Growth and survival rates were low because of competition with adult spruce for light, space, and nitrogen. Interestingly, seed production by adults was enhanced, which led to relatively high seedling numbers merely as a consequence of the many more seeds available for germination.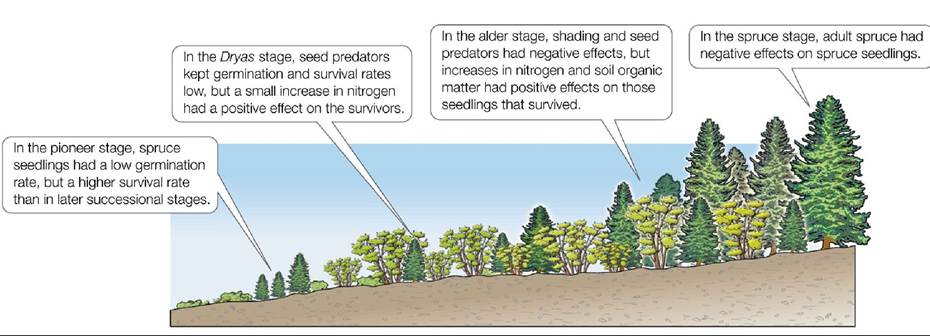
| Successional stage | ||||
| Effects on spruce seedlings | Pioneer | Dryas | Alder | Spruce |
| Positive | Higher survival | Higher nitrogen level Higher growth | Higher soil organic matter Higher nitrogen level More mycorrhizae Higher growth | Higher germination |
| Negative | Lower germination | Lower germination Lower survival Higher seed predation and seed mortality | Lower germination Lower survival Higher seed predation and seed mortality Root competition Competition for light | Lower growth Lower survival Higher seed predation and seed mortality Root competition Competition for light Lower nitrogen level |
FIGURE 17.12 Both Positive and Negative Effects Influence Succession Therelative contributions of positive and negative effects of other species on spruce seedling establishment changed across successional stages in Glacier Bay, Alaska. Positive effects equaled or outweighed negative effects in the first three stages, but the opposite was seen in the last spruce stage.
(After F. S. Chapin et al. 1994. Ecol Monogr 64: 149-175.) View larger imageThus, in Glacier Bay, the mechanisms outlined in Connell and Slatyer's models were operating in at least some stages of succession. Early on, aspects of the facilitation model were seen as plants modified the habitat in positive ways for other plants and animals. Species such as alders had negative effects on later successional species unless they were able to colonize early, supporting the inhibition model. Finally, some stages—such as the spruce stage, in which dominance was a result of slow growth and long life—were driven by life history characteristics, a signature of the tolerance model.
Secondary Succession in a New England Salt Marsh
What do other studies show with regard to Connell and Slatyer's three models? Mark Bertness and Scott Shumway studied the relative importance of facilitative versus inhibitory interactions in controlling secondary succession in a New England salt marsh. Salt marshes are characterized by different species compositions and physical conditions at different tidal elevations. The shoreline border of the marsh is dominated by the cordgrass Spartina patens, whereas dense stands of the black rush Juncus gerardii are found between the shoreline and the terrestrial border. A common natural disturbance in salt marsh habitats is the deposition of tidally transported dead plant material known as wrack (FIGURE 17.13). The wrack smothers and kills plants, creating bare patches where secondary succession takes place. Soil salinity is high in these patches because, without shading by plants, water evaporation increases, leaving behind salt deposits. The patches are initially colonized by the spike grass Distichlis spicata, an early successional species that is eventually outcompeted by Spartina and Juncus in their respective zones.

FIGURE 17.13 Wrack Creates Bare Patches in Salt Marshes Atidaldepositofwrackat
Rumstick Cove, Rhode Island, where Bertness and Shumway conducted their research on secondary succession. This dead plant material smothers living plants, creating bare patches with high soil salinity.
View larger imageBertness and Shumway (1993) hypothesized that Distichlis could either facilitate or inhibit later colonization by Spartina or Juncus depending on the salt stress experienced by the interacting plants. To test this idea, they created bare patches in two zones of a marsh and manipulated plant interactions shortly after the patches had been colonized (FIGURE 17.14). In the low intertidal zone (the Spartina zone, close to the shoreline), they removed Distichlis from half the newly colonized patches, leaving Spartina, and removed Spartina from the other half, leaving Distichlis. In the middle intertidal zone (the Juncus zone, closer to the terrestrial border of the marsh), they performed similar manipulations, with Juncus and Distichlis as the target species. Control patches, in which the colonization process was not manipulated, were maintained in both zones. In addition, they watered half the patches in each treatment group with fresh water to alleviate salt stress, and left half as controls.
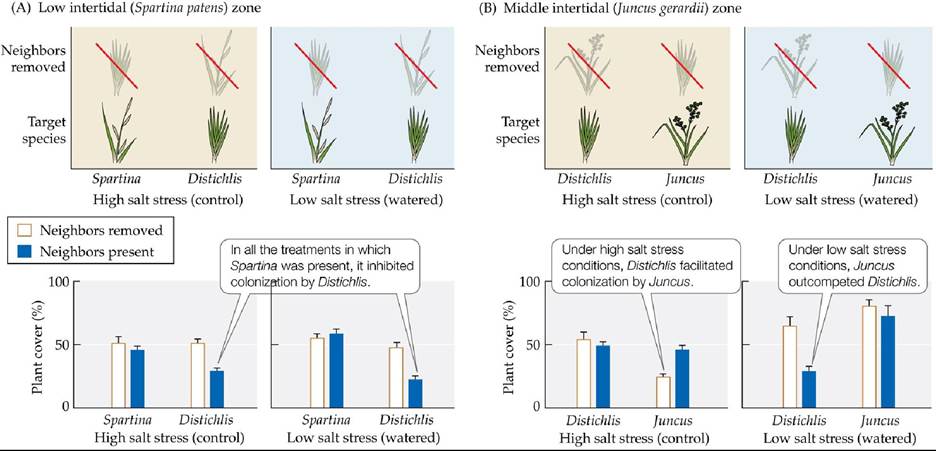
FIGURE 17.14 New England Salt Marsh Succession Is Context Dependent The trajectory of succession in salt marshes depends on soil salinity and the physiological tolerances of plant species. The kinds of interactions observed differed between the low intertidal zone (A) and the middle intertidal zone (B). Error bars show one standard error of the mean. (After M. D.
Bertness and S. W. Shumway. 1993. Am Nat 142: 718-724.) View larger image
After observing the patches for 2 years, Bertness and Shumway found that the mechanisms of succession differed depending on the level of salt stress experienced by the plants and the species interactions involved. In the low intertidal zone, Spartina always colonized and dominated the plots, whether or not Distichlis was present or watering occurred (Figure 17.14A). Distichlis was able to dominate only if Spartina was removed from the plots, so it was clearly inhibited by Spartina, the dominant competitor. In the middle intertidal zone, Juncus was able to colonize only if Distichlis was present or watering occurred (Figure 17.14B). Measurements of soil salinity confirmed that the presence of Distichlis helped to shade the soil surface, thus decreasing salt accumulation and reducing stress for Juncus. Distichlis, however, was able to colonize plots with Juncus only when salt stress was high—that is, under the control conditions. If plots were watered, Distichlis was easily outcompeted by Juncus.
These experimental manipulations confirmed that the mechanisms important to succession are context dependent. No single model is sufficient to explain the underlying causes of succession. In the middle intertidal zone, Distichlis was a strong facilitator of colonization by Juncus. Once this facilitation occurred, the balance was tipped in favor of Juncus, which outcompeted Distichlis (see Figure 17.14B). In the low intertidal zone, Distichlis and Spartina were equally able to colonize and grow in salty patches. If Spartina arrived first, it inhibited Distichlis colonization. If Distichlis arrived first, it persisted only if Spartina did not arrive and displace it (see Figure 17.14A).
Primary Succession in Rocky Intertidal Communities
Our final examples come from an environment where succession has been studied extensively: the rocky intertidal zone. Here, disturbances are created mainly by waves, which can tear organisms from the rocks during storms or propel objects such as logs or boulders into them. In addition, stresses caused by low tides that expose organisms to high or low air temperatures can easily kill them or cause them to lose their attachment to the rocks. The resulting bare rock patches become active areas of colonization and succession.
Some of the first experimental work on succession in the rocky intertidal zone was done on boulder fields in Southern California by Wayne Sousa, a graduate student at the time. Sousa (1979) noticed that the algae-dominated communities on these boulders experienced disturbance every time the boulders were overturned by waves. When he cleared some patches on the boulders and observed succession in those patches over time, he found that the first species to colonize and dominate a patch was always the bright green alga Ulva lactuca (FIGURE 17.15A). It was followed by the red alga Gigartina canaliculata. To understand the mechanisms controlling this successional sequence, Sousa performed removal experiments on concrete blocks that he had allowed Ulva to colonize. He found that colonization by Gigartina was accelerated if Ulva was removed (FIGURE 17.15B). This result suggested inhibition was the main mechanism controlling succession, but a question remained: If Ulva is able to inhibit other seaweed species, why doesn't it always dominate? Through a series of further experiments, Sousa found that grazing crabs preferentially fed on Ulva, thus initiating a transition from the early Ulva stage to other mid- successional algal species. In turn, the mid-successional species were more susceptible to the effects of stress and parasitic algae than the late successional Gigartina. Gigartina dominated because it was the least susceptible to stress and consumer pressures.
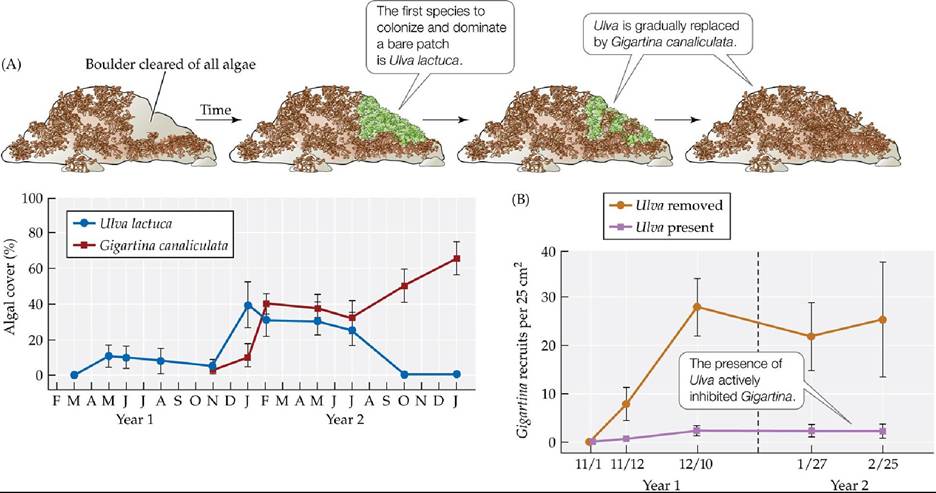
FIGURE 17.15 Algal Succession on Southern California Boulders Is Driven by
Inhibition (A) Drawings and data from a 2-year study of the successional sequence of algae in bare patches on boulder fields in the rocky intertidal zone of Southern California. (B) In a study
that lasted 4 months, Sousa performed removal experiments on concrete blocks to understand the mechanisms of succession in this ecosystem. Error bars show ± one standard error of the mean. (After W. P. Sousa. 1979. Ecol Monogr 49: 227-254.) View larger image
This view of rocky intertidal succession as driven by inhibition was the accepted paradigm for many years. Facilitation and tolerance were thought to be much less important in a system where competition for space was strong. More recent work by Terence Farrell and others (e.g., Berlow 1997) demonstrated that the relative importance of inhibition is probably much more context dependent than previously thought. In the more productive rocky intertidal zone of the Oregon coast, the communities include many more sessile invertebrates, such as barnacles and mussels, than Sousa's communities of the Southern California coast, where seaweeds dominate. In the high intertidal zone of Oregon, Farrell (1991) found that the first colonizer of bare patches was a barnacle, Chthamalus dalli. It was replaced by another, larger barnacle species, Balanus glandula, which was then replaced by three species of macroalgae, Pelvetiopsis limitata, Fucus gardneri, and Endocladia muricata. A series of removal experiments showed that Chthamalus did not inhibit colonization by Balanus, but that Balanus was able to outcompete Chthamalus over time, thus supporting the tolerance model. Likewise, Balanus did not hinder macroalgal colonization, but in fact facilitated it, lending credibility to the facilitation model.
But why and how would Balanus facilitate macroalgal colonization? Farrell suspected that Balanus protected the algae in some way, possibly from desiccation stress or grazing by limpets (herbivorous marine snails). To test this idea, Farrell created experimental plots from which Balanus, limpets, or both were removed, then observed macroalgal colonization in those plots. He found that macroalgae colonized all of the plots without limpets but had a much higher density in the plots with barnacles than in those without barnacles (FIGURE 17.16A). These results suggested that Balanus did indeed act to impede limpets from grazing on newly settled macroalgal sporelings.
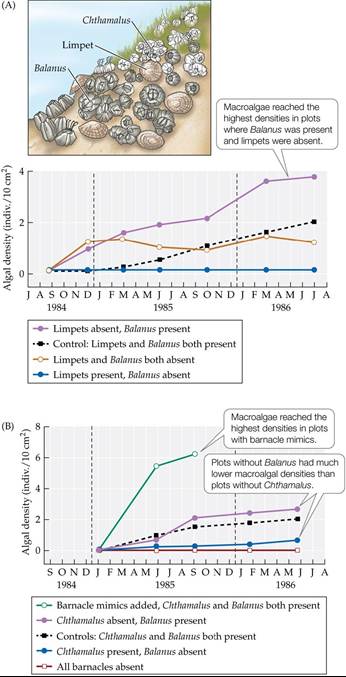
FIGURE 17.16 Algal Succession on the Oregon Coast Is Driven by Facilitation (A)
Changes in macroalgal densities over time were measured in plots from which Balanus barnacles, limpets, or both had been removed. The results suggested that Balanus facilitates macroalgae by reducing limpet grazing. (B) To understand the mechanisms of the facilitation, large barnacle mimics were added to some plots and compared with plots from which the real barnacle species —Balanus, Chthamalus, or both—had been removed. The results suggested that the larger the barnacle species, the better it protects macroalgae against limpet grazing and desiccation. (After T. M. Farrell. 1991. Ecol Monogr 61: 95-113.) View larger image
You might be asking yourself, Why doesn’t Chthamalus have the same facilitative effect on macroalgae that Balanus does? Farrell suspected that the reason was Balanus,s larger size (it is nearly three times wider than Chthamalus). By using plaster casts to mimic barnacles that were slightly larger than Balanus, Farrell found that these barnacle mimics had an even more positive effect on macroalgal colonization than did smaller-sized live barnacles of either species (FIGURE 17.16B). It seems likely that the smaller and smoother Chthamalus does not retain as much moisture, or block as many limpets, as the larger and more sculpted Balanus—or the mimics, for that matter.
More on the topic No one model fits any one community:
- Reviewers
- REVIEW OF FORENSIC ASSESSMENT INSTRUMENTS
- Sin, Impurity and Exorcism (4Q560)
- CHAPTER FOUR Town and Country Urban devotions and rural rituals
- THE THEORY AND PRACTICE OF EMPIRE-BUILDING