The biotas of biogeographic regions reflect evolutionary isolation
The six biogeographic regions described by Wallace are the Nearctic (North America), Neotropical (Central and South America), Palearctic (Europe and parts of Asia and Africa), Ethiopian (most of Africa), Oriental (India, China, and Southeast Asia), and Australasian (Australia, the Indo-Pacific, and New Zealand) (FIGURE 18.9).
It is no coincidence that these regions correspond roughly to Earth's six major tectonic plates. These plates are sections of Earth's crust that move across Earth's surface through the action of currents generated deep within its molten rock mantle (FIGURE 18.10). Before scientists understood the processes driving the movement of these plates, they hypothesized that the continents drifted over Earth's surface; thus, the name continental drift was given to the early theory describing these movements. There are three major types of boundaries between tectonic plates. In areas known as mid-ocean ridges, molten rock flows out of the seams between plates and cools, creating new crust and forcing the plates apart in a process called seafloor spreading. In some areas where two plates meet, known as subduction zones, one plate is forced downward under another plate. These areas are associated with strong earthquakes, volcanic activity, and mountain range formation. In other areas where two plates meet, the plates slide sideways past each other, forming a fault.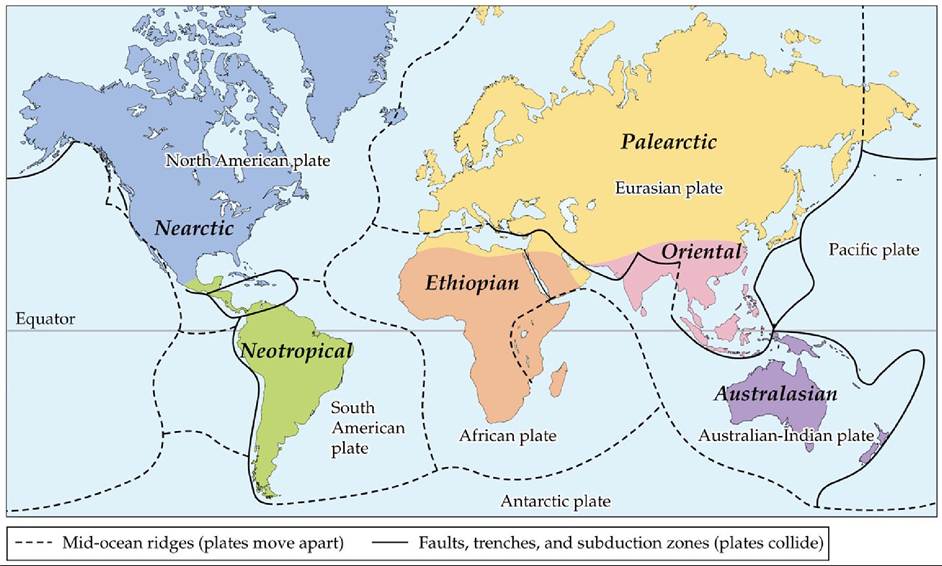
FIGURE 18.9 Six Biogeographic Regions Wallace identified six biogeographic regions using the distributions of terrestrial animals. These six regions roughly correspond to Earth's major tectonic plates.
Compare Wallace's 6 regions with the 11 biogeographic divisions shown in Figure 1.2. What types of data were used to expand the number of regions to 11?
(Based on A.
R. Wallace. 1876. The Geographical Distribution of Animals. Harper and Brothers: New York.) View larger image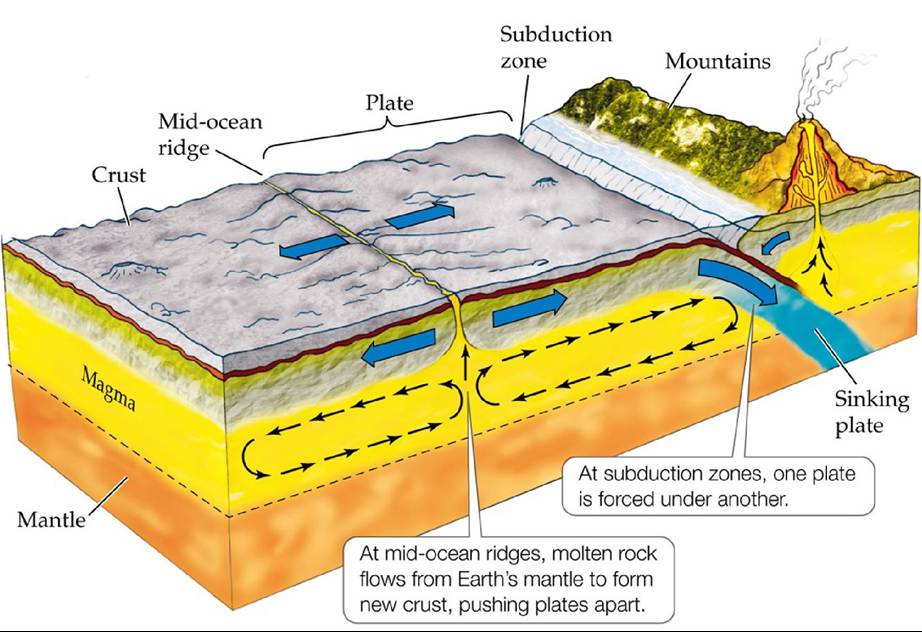
FIGURE 18.10 MechanismsofContinentalDrift Overgeologictime1Currentsgenerated deep within Earth's molten rock mantle move sections of Earth's crust across its surface. View larger image
As a result of processes such as seafloor spreading and subduction, the positions of the plates, and of the continents that sit on them, have changed dramatically over geologic time. For our purposes, let's consider the movements of the major tectonic plates since the early Triassic period (251 million years ago), when all of Earth's land masses, a single continent named Pangaea, began to break up (FIGURE 18.11A). At that time, there was a mass extinction (see Figure 6.19), which eventually led to the rise of the first archosaurs (precursors to dinosaurs) and the cynodonts (precursors to mammals). About 100 million years ago, during the mid-Cretaceous period, Pangaea had split into Laurasia to the north and Gondwana to the south. During that time, dinosaurs were in their heyday and mammals were small and a relatively minor component of the fauna. The end of the Cretaceous period was marked by another mass extinction, which resulted in the disappearance of dinosaurs. By the early Paleogene period (60 million years ago), Gondwana had separated into the present-day continents of South America, Africa, India, Antarctica, and Australia. Laurasia eventually split apart to form North America, Europe, and Asia. Most of these movements resulted in the separation of continents from one another, but some continents were brought together (FIGURE 18.11B). For example, North and South America joined at the Isthmus of Panama, India collided with Asia to create the Himalayas, Africa and Europe united at the Mediterranean Sea, and a land bridge formed between North America and Asia at the Bering Strait.
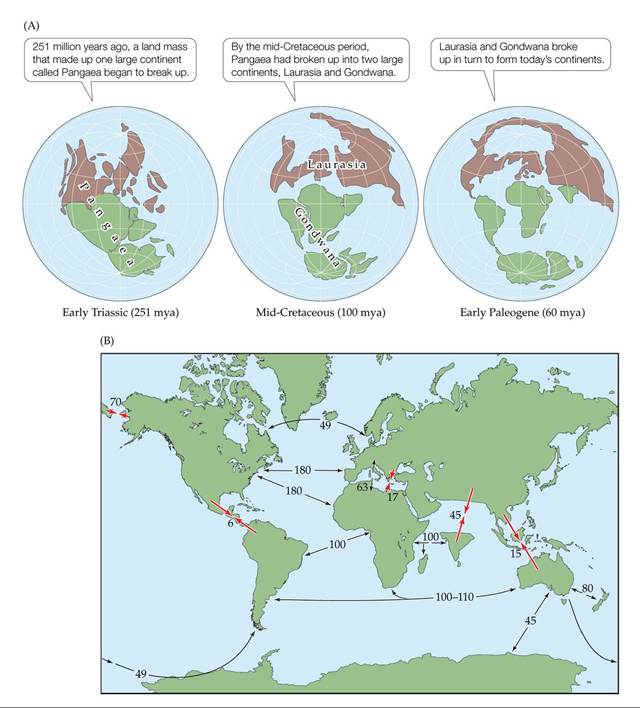
FIGURE 18.11 The Positions of Continents and Oceans Have Changed over Geologic
Time The locations of continents and oceans have changed dramatically over the last 251 million years because of continental drift. (A) The breakup of Pangaea. (B) A summary of the movements
that led to the configuration of the continents we know today. Larger, red arrows are labeled with the time (in millions of years) since land masses joined; Smaller, black arrows are labeled with the time since land masses separated.
As land masses separated, would you expect speciation to increase? Why or why not?
(After E. C. Pielou. 1979. Biogeography. Wiley: Hoboken, NJ.) View larger image
The movement of Earth's tectonic plates thus separated the terrestrial biota of Pangaea, united by geography and phylogeny, into biogeographically distinct groups of species by isolating them on different continents. The sequence and tempo of the continental movements has resulted in some biogeographic regions having very different flora and fauna than others. For example, the Neotropical, Ethiopian, and Australian regions, all once part of Gondwana, have been isolated for quite some time and have very distinctive forms of life. In other cases, however, distinct groups of species have been united. For example, the biota of the Nearctic region differs substantially from that of the Neotropical region despite their modern-day proximity. Because North America was part of Laurasia while South America was part of Gondwana, North and South America had no contact until about 6 million years ago. Within that time, however, many species have moved from one continent to the other (e.g., mountain lions, wolves, and the precursors of llamas spread to South America, while armadillos and opossums spread to North America), somewhat homogenizing the biotas of the two regions. Interestingly, there is also evidence that several families of terrestrial mammals went extinct once the two continents merged, suggesting that ecological coexistence was not possible for some species (Flessa 1975).
Finally, the Nearctic and Palearctic, both part of ancient Laurasia, have similarities in biota across what is now Greenland as well as across the Bering Strait, where a land bridge has intermittently allowed exchanges of species over the last 100 million years.The legacy of continental movements can be found in a number of existing taxonomic groups as well as in the fossil record. The evolutionary separation of species due to barriers such as those formed by continental drift is known as vicariance. Tracing the threads of vicariance over large geographic areas and long periods provided important evidence for early theories of evolution. For example, as Wallace began to amass knowledge of the distributions of more and more species and make geographic connections between them, his ideas about the origin of species started to solidify. In an 1855 paper titled “On the law which has regulated the introduction of new species,” he wrote, “Every species has come into existence coincident both in space and time with a pre-existing closely allied species.” Despite the biogeographic evidence of evolutionary connections among species, it took a few more years for one mechanism of evolution (i.e., natural selection) and its role in the origin of new species to be formally proposed by both Wallace (1858) and Darwin (1859).
Before we move on, it is important to consider contemporary research that updates and expands on the biogeographic regions first identified by Wallace. One recent study (Holt et al. 2013) used phylogenetic information acquired from DNA analysis and recent observations of global species distribution patterns to test whether Wallace's original biogeographic regions are supported by modern data. The researchers identified more biogeographic regions (a total of 11), some of which were the same and others of which were different from Wallace's original 6 regions (compare Figure 18.9 with Figure 1.2). This new analysis suggests that additional isolation mechanisms beyond continental drift are responsible for the different regions.
Interestingly, New Guinea and the Pacific Islands, separated from the Philippines by Wallace's line, emerge as a new biogeographic region, completely separate from the Australian or Oriental regions.Another recent analysis of biogeographic regions involved mapping the distribution of species in the oceans. After all, the oceans make up 71% of
Earth's surface area and, just as we have seen for continents, they are dynamic, in the sense that they are created, merged, and destroyed by the movements of Earth's tectonic plates (see Figure 18.11). The main question, then, is whether there are barriers to dispersal between oceans as there are between continents. Despite their appearance of connectivity, oceans do have significant impediments to the exchange of biotas: these impediments take the form of continents and currents; thermal, salinity, and oxygen gradients; and differences in water depth. Oceanographic discontinuities have isolated species from one another, allowed for evolutionary change, and created unique oceanic biogeographic regions (Briggs 2006). Unfortunately, delineation of marine biogeographic regions has been hindered by the extra complicating factor of water depth and by our basic lack of natural history and taxonomic knowledge of the deep oceans. One recent model by Adey and Steneck (2001) identifies 24 recognizable biogeographic regions for intertidal benthic marine macroalgae. Although it is hard to compare these macroalgal regions with terrestrial biogeographic regions, the analysis does suggest that the marine realm has much more biogeographic variation than previously realized.