Theory regarding the strength of competition
What is the level of competition in natural communities? Do most species experience large decreases in their own population size following large increases in the population sizes or population growth traits of other species with which they share resources? Are most species susceptible to complete elimination by a competitor with a small negative change in their own fitness or a small increase in the fitness of the competing species? More generally, how is similarity in resource use related to these population-level effects? One might think that these broad-brush questions about competition should already have been addressed in empirical studies.
While competition has been the focus of many field studies, they have not answered these questions, in part because of the limited duration and spatial extent of experimental treatments. In a relatively recent assessment, Adler et al. (2010, p. 1019) admit that, ‘Despite decades of research documenting niche differences between species, we lack a quantitative understanding of their effect on coexistence in natural communities’. One reason for this was stated many years earlier by a theoretical ecologist who chose the laboratory over the field for testing theory: ‘the direct study of... communities outdoors labors under crippling disadvantages... Environmental parameters vary uncontrollably in time and in space, inordinate numbers of... species are present, the individual life histories of most of these species and their interactions in pairs or in larger sets are poorly known, and experimental manipulations may be infeasible or doomed to yield ambiguous results' (Gilpin et al. 1986, p. 23).The answers to the questions raised in the previous paragraph can be approached theoretically as well as empirically. However, some level of theoretical understanding is a prerequisite for being able to refine the questions in a way that allows a meaningful approach by observation or experiment.
How should similarity be quantified? Which aspects of similarity have the greatest impact on competitive effects? How broad a spatial and/or temporal scale must be considered to obtain a meaningful estimate of effect size? These questions need to be understood from a theoretical perspective to determine what observations or experiments would be informative in measuring competitive effects. For example, is the presence of additional competing species likely to significantly change the per individual effect of oneCompetition Theory in Ecology. Peter A. Abrams, Oxford University Press. © Peter A. Abrams (2022).
DOI: 10.1093∕oso∕9780192895523.003.0007 consumer species on a second? The answer affects the applicability of any empirical measurement of competition between the two species. More generally, theory can suggest what types of organisms might be likely to experience stronger or weaker competition. And, because there are many potential measures of competitive strength, different perturbations to a system may change the various measures of competition in opposite directions. This first section deals with theory that discusses the relationships between measures of the similarity in resource use among co-occurring species and the population level interactions between them.
7.l.l Theory from the early l970s
Early natural history observations by Hutchinson (described in his 1965 book) suggested that significant differences between species were needed for them to coexist, and that competition between species might lead to a constant ratio of sizes in sets of three or more competitors. Much earlier, Volterra (1931) had argued that, if there were several competitors for a single limiting factor, the one capable of surviving and reproducing at the lowest level of that factor would displace all others. Hutchinson’s later observation suggested that a different sort of limitation might be needed for coexistence of species that consumed many resources (as most consumers do).
From 1967 through 1974 Robert MacArthur, Richard Levins, and Robert May, inspired by these and other early observational results, established what is still a widely accepted framework for thinking about the relationship between similarity in resource use and the strength of interspecific competition, and the consequences of that relationship for coexistence.The initial analysis by MacArthur and Levins (1967) was based on a Lotka- Volterra model with additional assumptions about how the difference between two consumer species in their mean values of a consumption-related resource characteristic translated into a competitive effect. These assumptions were also part of MacArthurs (1970, 1972) consumer-resource model. All three of these analyses assumed a constant homogeneous environment. All argued that coexistence was very unlikely (although not strictly impossible) for species that had a high enough overlap in resource use, but one that was significantly below complete similarity. The requirement for substantial differences in resource use was later suggested to be more stringent by May (May and MacArthur 1972; May 1973,1974), using Lotka-Volterra models that included environmental variation in consumer per capita growth rates. In these works, a level of similarity well below complete identity always led to exclusion. This result was the basis of a large amount of later theoretical work on the idea of a ‘limiting similarity’ of competitors.
All these initial studies contained a core set of assumptions that were not based on any supporting evidence. First, the resource use of each species could be characterized by plotting their maximum resource capture rates as a function of a single variable (often a measure of the size of the resource item). Secondly, this function was Gaussian. These two assumptions seemed to make it possible to obtain a simple analytical formula for the amount of competition between species just by measuring the distance between the peaks of the ‘utilization curves' of the two competitors.
Figure 7.1 illustrates the basic idea. Here, the trait distinguishing resources is given on the x-axis, and the maximum per capita consumption rates of three evenly spaced consumers is given by the curves, Ci(x) for consumer species i. In the figure, these are Gaussian curves with a standard deviation of 1, and with mean values also separated by 1 unit on the resource axis. The competition coefficient (formula (3.2)) proposed by MacArthur and Levins (1967), and still widely used today is:
This expression has a maximum value of 1 when the two utilization curves are identical. The assumption of Gaussian utilization curves having a common mean and a common standard deviation then makes eq. (7.1) equivalent to exp(-D2/ W2), where D is the distance between the curves of the two species, and W is the (common) standard deviation of the curves. In the situation illustrated (with D - W - 1), the competition coefficient between adjacent species is approximately 0.368. This allows coexistence for a reasonably broad range of other parameters when there are only two competitors, but that range is greatly reduced in the case of three or more species as in Figure 7.1. Even coexistence of two species rapidly becomes less likely when the D/ W declines below unity; α = 0.779 when D/ W - 0.5.
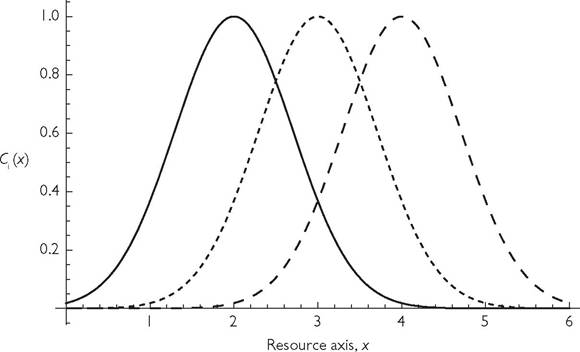
Fig. 7.1 A plot of standard resource utilization curves for three competitors as envisioned by MacArthur. The curve plots the value of the consumption rate of resources having the quality x (e.g., size) given on the x-axis. In the figure the standard deviation of these Gaussian curves is unity, as is the spacing between the curves. This spacing was MacArthur and Levins’ (1967) ‘limiting similarity’.
The above formula from MacArthur and Levins' (1967) Lotka-Volterra model turned out to be identical to the Lotka-Volterra approximation to MacArthur's (1970, 1972) consumer-resource model under the assumptions of an infinite number of substitutable resources having identical growth parameters and identical nutritional values to the consumer.
This is the extension of eqs (3.4) to a large number of resources. The effects of unequal values of the parameters on the competition coefficients are shown in Chapter 3 immediately below those equations. MacArthur also used the further (unstated) assumption that extinction of one or more resources did not occur. Neither uniform growth and nutritional value nor absence of resource extinction is likely to apply generally, as has been noted in preceding chapters. The derivation of the formula for competition coefficients required assumptions about independent logistic resources of equal value to the consumer that were consumed by linear functional responses, and whose consumption rates combined additively to determine consumer fitness. All of this was required to reduce the model to Lotka-Volterraform (Schoener 1974a; Abrams 1983b).A key feature of extensions of the 2-species Lotka-Volterra model to multiple competitors was that the interaction of any pair of species was independent of the presence of all the other consumer species. The three-or-more consumer version of the MacArthur consumer-resource model suggested that this was indeed the case. However, it should be noted that this is again dependent on the linear per capita growth rates of that model and on the continued existence of all the resources. A third consumer species often increases the total consumption rate of some resources, which has the potential to cause their extinction.
For the case of similar Gaussian utilization curves for all consumers, eq. (7.1) means that the range of a neutral consumer parameter (e.g., its per capita death rate) that allows coexistence is narrow when the distance between the utilization curves is significantly less than the standard deviation of the curves, particularly with three or more species. Early work (May and MacArthur 1972, May 1973, 1974) added stochastic variation to the consumer per capita growth rates and argued that this would ensure extinction of one competitor in an array of three or more, if the species had utilization curves separated by slightly less than one standard deviation of the curves.
The stochastic forms of the models assumed that conditions near an equilibrium point also applied when one or more of the competing species was close to extinction (Abrams 1976, 1983b; Turelli 1978).The evolution of traits related to resource use is also expected to influence observed levels of competition. However, in the early 1970s the evolutionary response to competition had not been analysed rigorously. Brown and Wilson (1956) argued that competitors would diverge in characteristics related to resource use. Grant (1972) suggested the possibility of convergence, but did not present a mechanism based on resource use. MacArthur did not analyse the evolutionary aspects of competition, and mathematical theory for those aspects was not developed until later in the decade (Lawlor and Maynard Smith 1976; Slatkin 1980). The evolution of competitors will be discussed in Chapter 11. However, early evolutionary models (Slatkin 1980; Taper and Case 1985; Abrams 1986a) did not predict that species would evolve
Resource use and the strength of interspecific competition • 147 to the limiting similarity predicted by MacArthur and Levins (1967). Regardless of the direction of evolutionary responses, the end result of those responses must be consistent with any ecological restrictions on the coexistence of very similar species. MacArthur’s ecological theory seems to imply that species could only achieve their ‘limiting similarity’ by convergent evolution of initially more different forms, but there was no mechanism suggested for such a change. If two species initially violated the ‘limiting similarity’, it is unlikely that they would have diverged evolutionarily at a rapid enough rate to achieve coexistence prior to exclusion. This problem was not noted in the literature from the 1970s. Even if divergence was rapid enough to prevent extinction, there is no theoretical reason why it should stop at the exact amount that was sufficient to produce long-term coexistence.
1. ∖.1 Early questioning of MacArthur’s limiting similarity
It did not take long for the above early results based on MacArthur’s approach to be challenged. Theory from the decade following MacArthur’s works showed clearly that there were many possible relationships between ‘similarity’ (however it was measured) and any chosen measure of the ‘strength’ of competition. Various works (Schoener 1974a, 1976, 1978; Abrams 1975, 1977, 1980a) showed that similaritycompetition relationships were affected by: (1) the nature of resource growth, and differences between resources in their population growth functions; (2) the intake rates of each resource as a function of all variables affecting that rate (consumer functional responses); and (3) the effects of those intakes on consumer demographic rates (numerical responses).
The first factor (resource growth function) was acknowledged in MacArthur’s original (1970, 1972) work, but was hidden by his decision to concentrate on the simplest case where all resources had identical logistic growth functions. MacArthur briefly noted that abiotic resources produced a different formula for the competition coefficient, but he chose to emphasize that the expression (7.1) was approximately correct when abundances were very close to their equilibrium values. Unfortunately, questions about coexistence depend on what happens far from equilibrium. MacArthur also failed to consider the possibility that extinction of some resources would be caused by efficient consumers. However, Hsu and Hubbell (1979) showed that that competition was changed by such extinctions. A final problem was that the most commonly assumed differentiating factor was the size of the resource items. However this usually leads to unidirectional change in the weighting factors; for example, larger items have a larger nutritional value (a larger b in eqs (3.4) or (6.1)). The values of the resource growth parameters are also known to change systematically with size, and it is highly unlikely that all of these unidirectional changes would exactly cancel each other. Figure 6.3 showed the effects of nonlinear resource density dependence on the competition coefficient.
The second factor limiting the applicability of expression (7.1) was the form of the functional response. The problem with the underlying assumption of type I functional
responses was explored in Abrams (1980a), who showed that type II responses made competition coefficients change greatly with consumer population sizes (see Figure 6.4). Most subsequent models continued to assume linear functional responses or to use MacArthur’s formula without noting its dependence on this assumption.
The third factor affecting the strength of competition was the nature of the numerical response. Leon and Tumpson (1975) initiated theoretical work on one of the most important aspects of numerical responses; i.e. how different resource intake rates combined to determine per capita growth rates. Leon and Tumpson considered the effects of non-additive contributions of different resources to per capita population growth; Tilman (1982) popularized their approach—like the original work by Leon and Tumpson (1975) he restricted his modelling to the case of exactly two limiting resources. Both works also shared assumptions of independent functional responses to each resource, and linear numerical responses to the intake rate of the limiting resource(s). A broader exploration of the range of numerical responses or their effects on competition has yet to appear.
In any case, studies from the 10 years following MacArthur’s (1972) book demonstrated that the effects of the abundances of competing species on each other’s growth rate were usually nonlinear. A sustained perturbation to the per capita growth rate of one consumer species that did not affect any consumer-resource interaction directly would have a different effect on its competitors’ per capita growth rates depending on the initial abundances of the perturbed species and the competitor(s), and depending on the magnitude of the perturbation. This was one of the main points of the theoretical articles from the 1970s by Schoener and by Abrams. It means that any single measure of the strength of competition may only be a good approximation over a narrow range of initial abundances.
The theoretical arguments for a limiting similarity were also shown to be weak in the 1970s (Schoener 1976, 1978; Abrams 1975, 1977; Turelli 1977, 1978). The results depended on assumptions about population dynamics of consumers and resources that were seldom satisfied, and the stochastic versions of the theory had been improperly analysed. The work suggesting an absolute limit to similarity was based on extrapolating very limited simulation results (May 1973,1974). This extrapolation turned out to be invalid (Turelli 1977, 1978, 1981). The original May and MacArthur results also assumed uncorrelated environmental effects that were independent of similarity, an assumption that was both unlikely and could reverse the qualitative results (Abrams 1976). Finally, the limiting similarity results with or without stochastic variation assumed the underlying linear functional response/logistic resource growth model of MacArthur, as well as a single resource at each position on the resource axis. Relaxing either of these assumptions could allow coexistence of similar species over a much wider range of relative mortality rates of the consumers (Abrams 1975, 1983b). However, some other potential assumptions made coexistence more difficult than in the MacArthur model.
The idea of simple Gaussian resource utilization functions was also far from a good approximation in the real-world systems where it had been closely examined (Wilson 1975). Thus, although a requirement for coexistence does limit the similarity
Resource use and the strength of interspecific competition • 149 of species that differ in mean fitness-related parameters, there was no generally applicable rule for the nature of these limits (Abrams 1983b). The difficulty of achieving coexistence for any given measure of similarity depended strongly on the details of the consumer-resource interactions involved. A general problem with using the Lotka- Volterra model for sets of three or more species was that it assumed that pairwise interactions were independent of the presence and abundance of additional competing species. This was shown to almost never be true for simple models based on consumer-resource interactions of homogeneous populations (Abrams 1983a). The presence of these ‘higher-order’ interactions meant that the relationship between similarity and any measure of competition for any pair of species was almost always changed by the presence of additional competing species.
If most organisms were close to a limiting similarity with their competitors, extinctions should be very frequent, as new mutations or environmental changes with species-specific effects would likely provide a slight advantage to one of the species. There is no general reason why being reduced in population size should increase selection for traits that would save the population. In almost all cases, a trait that provided an individual advantage would have been at least equally advantageous before the process of exclusion began. Mutations with such a trait would become less likely as population size decreased.
Whether or not consumers are close to a limiting similarity, it is important to try to understand the causes of any differences in the strength of competition that are observed for different taxonomic groups or in different ecological circumstances. All of this depends on having a quantitative description of the interaction. Here again, theory from the previous millennium provided a variety of reasons why the assumption of linear competitive effects (from both the Lotka-Volterra and MacArthur consumer-resource models) would often be misleading.
7.1.3 Recent and potential future theory on overlap and competition
The preceding section concentrated on theory published between 1970 and the early 1980s. Some later work has continued to explore the relationship between similarity in resource utilization and competition. In consumer-resource models with living resources, the departures from Lotka-Volterra dynamics and MacArthur’s competition coefficients are particularly large when one or more resources may become extinct because of a perturbation to one or more of the consumer species. Abrams (1998) showed that the increasing relationship between similarity in resource use and magnitude of the competitive effect on equilibrium abundance (assumed to be universally true) did not hold for competition between predators in 2-predator-2-prey models when the predators were able to survive on resource abundances that were sufficiently low relative to their carrying capacities. This efficiency of the predators could produce apparent competitive exclusion of one resource in a system with a single predator type, if that predator was moderately specialized. Exceptions to the supposed rule of competition increasing with similarity were even more common
in models with competing prey species (Abrams and Nakajima 2007) and in models with an infinite array of independent prey species (Abrams et al. 2008a).
The resource extinctions in these models all represent cases of exclusion due to apparent competition, which is itself a widely accepted phenomenon (Holt and Bonsall 2017). So far as I know, however, the implications of recognized cases of apparent competitive exclusion of resources for competition between two or more consumers have yet to receive any empirical study. This is at least potentially due to the almost complete focus on scenarios with a single predator species when apparent competition is discussed (Holt and Bonsall 2017). The presence of a second predator in a system with two prey does, in fact, reduce or eliminate the apparent competitive effect between the prey, if apparent competition is measured by the effect of a small change in a neutral parameter of one prey species on the abundance of its ‘apparent competitor' (see below or Chapter 5). It is possible that apparent competitive exclusion is rare in cases with multiple predator and prey species; if so, determining why this is the case would represent a major advance in our understanding of the nature of both competition and apparent competition in natural communities. However, apparent competitive exclusion in situations with two or three consumers and three or more resources is common in simple models (Abrams et al. 2008a, Abrams and Rueffler 2009). Returning to cases with a single predator, one possibility is that consumers in natural communities seldom decrease their biotic resources by the relatively large amounts required for resource extinction. However, Abrams et al. (2008a) pointed out that empirical work on the effects of predators on prey included many examples with prey reductions of a factor of 10 or more (Shurin et al. 2002).
Apparent competitive resource exclusion is not the only factor that limits the applicability of MacArthur's competition-similarity relationship. Another problem is the idea that the sets of maximal per capita resource capture rates of two consumers are sufficient to determine the competitive effects between them. Almost all functional forms and parameter values in a standard consumer-resource model affect any measure of the strength of competition. The determinants of competitive effects include the form of and parameters characterizing the consumers' functional and numerical response functions, and the resource growth rate functions. The latter include any interactions between different resources. The presence of resource choice behaviour on the part of the consumers also affects the shapes of their functional responses (Abrams 2010a, b) and the amount of competition between them (Abrams 2006a, b). These works primarily discuss behavioural effects on competition in the context of two resources. The theoretical effects of adaptive consumer foraging in the context of a continuous array of resources have not been studied. This is probably because of the lack of a simple closed-form solution for any reasonably widely applicable sets of functional forms for the behavioural model components in these systems. In any case, the lack of uniform applicability for any of the potential functional forms, and the relatively small number of resources per consumer species identified in food web models (Cohen et al. 1990) both suggest that exploring cases with a relatively small number of resource types is likely the most useful approach.
MacArthur’s consumer-resource model showed that if there were different nutritional values for different foods, a formula for the competition coefficient in the Lotka-Volterra approximation to that model would entail higher weighting for the more valuable foods. However, most of his analysis of similarity and competition assumed all foods were equally valuable, which was already known to be inaccurate in almost all systems. Schoener (1974a) later re-emphasized the importance of the weighting terms in the full version of MacArthur’s formula. Nevertheless, they continued to be ignored. In part this was due to a lack of experimental study of their values at that time. However, the variable that was most commonly assumed to differentiate the use patterns of competitors was resource-item size (e.g. seed size in seed-eating birds). Caloric or nutrient content (which would likely be the most important property influencing effects on demographic rates) are often roughly proportional to weight. However, the handling times usually are not. Thus, the lack of terms modifying the products of attack rate (Ci(x)Cj(x)) in eq. (7.1) is a serious deficiency. The weighting terms also include resource growth parameters, which are also likely to differ between different resource types.
Including weighting terms for different resources in a model of competitive dynamics is even more important when the resources are not linearly substitutable in their effects on fitness. The extreme case of two nutritionally essential resources implies that the resource intake vs per capita growth rate relationship differs qualitatively depending on which resource is limiting. Leon and Tumpson (1975) and Tilman (1980, 1982) discussed these sorts of essential resources, but did not have a proper analysis of how they affected a Lotka-Volterra approximation to the system. Tilman (1982) presented an erroneous derivation that purported to show that systems with two nutritionally essential resources could be reduced to a Lotka-Volterra form with fixed coefficients of the interaction terms. Abrams (1987b) later re-examined this case, and a related model in which adaptively flexible behaviour resulted in nonlinear competitive effects at all consumer densities. Chase and Leibold’s (2003) widely cited book had a thorough discussion of the importance of relative resource conversion efficiencies in determining competitive effects. However, they assumed linear functional responses for each resource. Adaptive functional responses with nutritionally interacting resources can have a variety of forms (Abrams 1987c), and the models of competition between species with such adaptive functional responses can have a variety of unusual dynamics, some of which were touched upon in Chapter 6. Even without nutritional interactions, the presence of different ratios of energy or nutrient content to handling time in different resource types makes it advantageous to ignore lower-quality foods when the abundance of better foods is high (Pyke et al. 1977). This greatly changes the form of the functional response (Fryxell and Lundberg 1998; Ma et al. 2003). The resulting effects on interspecific competition have largely been ignored.
Most of the above discussion has assumed that competition was purely exploitative. Early empirical studies (reviewed in Schoener 1983) showed that this was far from universally true. Much evidence and purely logical arguments suggest that interference should frequently exist, both within species (DeAngelis et al. 1975; Abrams 1984b, 1992c, 2010b; Abrams and Ginzburg 2000), and between species (Schoener 1983; Abrams 1986a, 2010b). The already wide variety of potential competitionsimilarity relationships is increased by the possibility of an interference component to the interaction. Interference can affect any or all of the separate functions that make up the per capita growth rate of a consumer. In addition, interference is likely to have a wide range of functional forms for any single component of consumer growth. However, relatively little theory has been developed to describe this range of possibilities.
One of the more common approaches to modelling interference has been to make functional responses ‘predator-dependent’. In the case of a type II response this usually involves adding a term proportional to predator abundance in the denominator of the response. This formulation arises when increased predator abundance interferes with the capture process (DeAngelis et al. 1975). Interference reduces capture rates generally in the model, which seems to imply that there is no benefit to the successful ‘interfering’ individual. However, ‘stealing’ a food item should increase the per capita food intake of the ‘thief’, even though it may decrease the mean capture rate for the consumer population. Any model that seeks to understand when interference will be observed or how it will evolve over time should consider these individual-level costs and benefits. However, they are seldom included in ecological models with predator interference. An even more important omission is the scarcity of theory dealing with the combined effects of both intra- and interspecific interference; the preponderance of experimental studies of functional responses deal with a single consumer and single resource (Jeschke et al. 2002).
Cases of two competing predators with interference have occasionally been modelled explicitly, but most examples have simply assumed an additive combination of independent effects of the two predators in the denominator of the functional response. Another possibility included in some models is that the interference increases per capita death rate (e.g., Abrams 1986a; McPeek 2019a), and here again, independence of effects is usually assumed. A broader approach would be to derive mathematical representations of the interference by assuming adaptively variable behaviours of the different species involved. In most cases, intraspecific interference enables coexistence of species having greater overlap in resource use, with interspecific interference having the opposite effect.
Another aspect omitted from most early theoretical works on competitionsimilarity relationships is the impact of different forms of spatial and temporal variation in resource uptake rates by the various consumers. These had been recognized as influencing the ‘strength’ of competition relatively early in the modern history of competition theory (Haigh and Maynard Smith 1972; Chesson and Warner 1981; Abrams 1984a), but are still relatively poorly understood (as documented in the next three chapters). In most cases, the measure of‘similarity’ between species must take differences in temporal responses between consumers in all resource- related parameters into account. The effects of such differences will usually depend on other aspects of the resource dynamics that affect the rate of response of resource
Resource use and the strength of interspecific competition • 153 abundances to the temporal changes in interaction parameters. More details are provided in Chapters 8, 9, and 10.
7.1.4 Continued use of outdated similarity-competition relationships
Returning to the simpler case of a single homogeneous and temporally constant environment, the studies from the 70s and early 80s reviewed earlier in this section had shown that no single function based solely on the sets of per capita capture rates of all consumers (e.g., that of MacArthur 1972) would accurately represent competitive effects. The nonlinearity of effects also means that competition between two species is seldom measurable by a single number. Nevertheless, MacArthur’s proposed function for an all-encompassing ‘competition coefficient’ is still being widely used, as noted by Leimar et al. (2013) and Falster et al. (2021).
This raises the question of why the early critiques were and still are being disregarded by so many. The answer to this question is not obvious. There is, of course, no universal answer. The widespread prior use of the Gaussian form has been given as the basis for its use by some of the few authors who have acknowledged that there are other possibilities (e.g. Day 2000). Once a certain amount of theory has accumulated, it gains a life of its own, and continues regardless of demonstrations of its inadequacies or limited range of applicability. And that is particularly true when (as is true here) there exists a broad range of alternatives, none of which has universal applicability. This situation is similar to the use of the logistic and chemostat equations to represent resource growth (as in many of the models discussed in this book).
In a recent article, Falster et al. (2021) cite D’Andrea and Ostling (2016) as showing that ‘there are few models that relate traits to fitness via competition for a common resource without including a phenomenological competition function’ (Falster et al. 2021, p. 264). This is definitely not the case, unless one adopts an abnormally narrow definition of‘trait’. Every consumer-resource model has a relationship between traits and fitness, at least if demographic and ecological parameter values are regarded as traits. Even if more easily measured traits such as body size are the focus, determining their effect on fitness requires knowledge of their relationship to consumer-resource parameters. The framework Falster et al. (2021) are attacking seems to be the one that I criticized above, consisting of independent (non-interacting) resources that can be arrayed along a one-dimensional axis, and that all have logistic growth and are consumed by predators with linear functional responses. However, there have been a large number of published models that do not assume this. Even by the time that the one-dimensional framework was first being applied by scientists other than MacArthur’s students and collaborators, Schoener’s (1974b) review of field studies had documented the fact that multi-dimensional niche separation was the norm in nature. While May (1973, 1974) had briefly discussed extending the Gaussian utilization-curve framework to two or more resource dimensions, this has yet to inspire a large body of theory based on multi-dimensional partitioning, in spite
of its known predominance in natural systems (Schoener 1974b). At about the same time Roughgarden (1974) showed that different shapes of the consumers’ utilization functions produced different shapes of the competition coefficient as a function of similarity. The shapes considered in that work often led to much greater permissible similarity of competitors than suggested by MacArthur (1972). Wilson (1975) showed that the MacArthur formula was inaccurate for several empirical systems.
The point of the Falster et al. (2021) article is that Lotka-Volterra models with MacArthur’s competition function do not provide a reasonable description of similarity and competition in plants, which is certainly true in most if not all cases. However, this point (and the remedy of using consumer-resource models rather than the Lotka-Volterra) had been put forward 50 years ago in Grenny et al. (1973) and Williams (1972). Falster et al. (2021) state, ‘The more promising approach will be to model depletion of resources by populations in relation to their traits, with its consequences for fitness landscapes and competitive exclusion’. However, nearly 50 years earlier, in the same journal W.E. Neill (1974, p. 407) had stated, ‘it may be more profitable now to set the Lotka-Volterra models aside, and to begin to emphasize mechanisms and dynamic aspects of resource utilization as a reasonable alternative’ It is clear that history has been forgotten.
7.2
More on the topic Theory regarding the strength of competition:
- Resource use and the strength Ofinterspecific competition
- The first revival of competition theory
- Choosing articles to represent current competition theory
- Theory’s roles in ecology and competition
- The need for resources in competition theory
- Abrams Peter A.. Competition Theory in Ecology. Oxford University Press,2022. — 336 p., 2022
- Competition between species in theory and reality
- Forgotten results in ‘modern competition theory’
- Competition theory past and present
- Competition theory its present state
- The theory of vague propositions outlined in chapter 6 appealed freely to the orthodox Bayesian theory of credences and learning, which assumes, among other things, that the correct theory of rational credence is one governed by the classical axioms of probability theory.
- Species interactions vary greatly in strength and direction
- The strength of competitive interactions affects coexistence