Other neglected aspects of consumer-resource models
The simple models considered here lack many near-universal features of natural systems. A seldom-noted feature of the MacArthur model is its assumption that
Negativity, constancy, and continuity of competitive effects • 139 resources are substitutable; i.e.
the per capita growth rate is an increasing function of a weighted sum of the individual consumption of rates of each resource. Consuming two or more resources has repeatedly been shown to imply a nonlinear effect of the consumption rate of one resource on the consumer’s growth rate (Leon and Tump- son 1975, Tilman 1982). The addition of adaptive adjustment of intake rates (which is particularly likely for non-substitutable resources) further changes the nature of the interaction between consumers in such system (Abrams and Shen 1989). For the most part, competition theory that deals with interdependence of resource effects on consumer fitness remains focused on the simple 2-resource models considered by Leon and Tumpson (1975) and Tilman (1980, 1982). These are characterized by functional responses that are only affected by the abundance of one resource. Even if the resources are nutritionally substitutable, either adaptive switching behaviour (Abrams and Matsuda 2003, 2004) or digestive constraints (Abrams 1990a, b) are sufficient to cause the abundance of one resource to affect the consumption rate of another, and thus, to affect competitive interactions involving either resource. Properly accounting for these scenarios requires models of the dynamics of behaviours that determine resource choice. This is another largely neglected topic in competition theory (see Chapter 3 and Abrams et al. 2007).Another omission from most recent competition theory is that age and stage structure are lacking in most models, whether or not they contain resources. De Roos and Persson (2013) summarize the case for inclusion of population structure in models.
Determining the nature and outcome of competition becomes more complicated in these cases, and there is a possibility of alternative stable equilibria in many models, even with a single consumer species (Abrams and Quince 2005; Schreiber and Rudolf 2008; de Roos and Persson 2013). Population structure makes the representation of competition more complicated, even in the absence of explicit resource dynamics. As a result, the many potential impacts of different types of population structure have received little attention. The presence of between-individual variation in resource use, even if this is not related to size or age, also alters competitive interactions.Most traditional models of competition also neglect temporal variation in the parameters of the model. The majority of the theory regarding competition in variable environments has not been based on traditional differential equation models of seasonal environments; Li and Chesson (2016) is one of the few exceptions to this. Chesson has produced a large body of important work on competition in temporally variable environments (Chesson and Warner 1981; Chesson 1994, 2000b, 2003, 2018), but most of this lies outside of the ordinary differential equation framework that has dominated the rest of competition theory. (The theory developed by Ches- son has recently been reviewed by Barabas et al. (2018), Barabas and D’Andrea (2020), and Chesson (2020a, b).) Chapters 8 and 9 will take a closer look at the impacts of seasonal variation in parameters in the context of simple consumer-resource theory.
A final neglected element of competition theory is the impact of changes in nonneutral parameters—those that affect the per capita growth rate of more than one species. Once a consumer-resource framework is adopted, there are usually a number of such parameters. In MacArthur’s very simple consumer-resource model, every
per capita resource capture rate parameter (denoted cij, or Cij in the models considered here) is non-neutral.
A beneficial (i.e., fitness-increasing) change in any of these parameters need not produce the same direction of change in population size as a beneficial change in a neutral parameter. (The same is of course true of detrimental changes.) Some of the effects of changes in capture rates in competitive systems were discussed in Abrams (2002, 2003, 2004a, 2009d) and Abrams et al. (2003). One of the most important features of a predator’s (consumer’s) capture rate is that adverse changes can increase the predator’s population size; this happens when the resource is biotic and is ‘overexploited’. The latter term simply means that the resource abundance is low enough that the reduced resource abundance (R) from increasing capture (C) is sufficient to reduce the product, CR. In multi-resource systems, the meaning of overexploitation is that increases in one or more capture rates will result in a lower total intake at equilibrium. In some competitive systems, this means that a beneficial change to one consumer can increase the population size of its competitor, or that a detrimental change to the first can decrease the population size of its competitor.The 2-consumer-2-resource MacArthur model (eqs 6.1a) displays these counterintuitive changes when the equilibrium resource densities are below half their carrying capacity in the original system (Abrams 2002, 2003). Figure 6.9 provides an example. Each consumer in the system illustrated initially has a fourfold greater consumption rate of its preferred resource, but the consumers are otherwise equivalent. This yields a competition coefficient at equilibrium of 0.4706, which is significantly
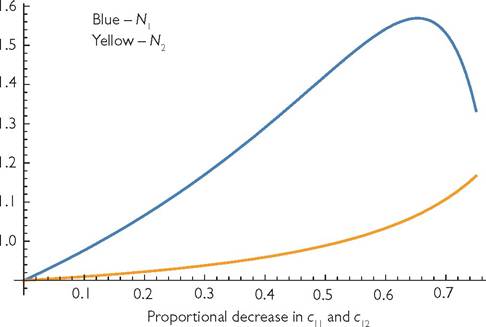
Fig. 6.9 The changes in the abundances (N1 and N2) of two consumer species in a MacArthur model (eqs (6.1a) with two consumers and two resources. The initial parameters are: r1 = r2 = k1 = k2 = b1 = b1 = 1, c11 = c22 = 0.8, c12 = c21 = 0.2; d1 = d2 = 0.1.
The x-axis gives the fractional reduction in consumer 1’s capture rates, c11 and c12. The graph ends at the x-value (0.75) where consumer 1’s equilibrium drops to zero, as explained in the text.Negativity, constancy, and continuity of competitive effects • 141 greater than that of the traditional case of multiple resources and Gaussian utilization curves separated by one standard deviation of the curves (see Figure 7.1). Figure 6.9 shows how the two equilibrium consumer abundances change when consumer 1 experiences equal proportional declines in its consumption rates of both resources. The fractional decline is given on the x-axis. Consumer 1 actually increases in abundance with proportional decreases in its consumption rates, and increases much more than its competitor, whose consumption rates remain constant. Consumer 1 benefits from reduced overexploitation of its preferred resource. Consumer 1 begins to decline once its proportional decrease in consumption rates exceeds approximately 65.3%. The graph ends at a proportional decline of 75%, because, at exactly that point, several discontinuous changes occur. Consumer 1 goes extinct, consumer 2 more than doubles in abundance to a density of 2.5, and consumer 2’s preferred resource goes extinct. (Note that both species had initial abundances of 0.9.) The exact pattern of change depends on the degree of initial resource partitioning and the efficiency of the consumers, as measured by the extent to which they reduce the resource population sizes. Abrams (2002) provides some additional examples.
The example in Figure 6.9 is a good illustration of Lewontins statement from the first paragraph of this chapter. The pattern of changes in abundance shown here would be totally incomprehensible using a Lotka-Volterra model, or MacArthur’s model with resource extinction assumed not to occur.
More on the topic Other neglected aspects of consumer-resource models:
- Important aspects of consumer-resource relationships
- Competition in other 2-consumer-l-resource models
- 3.4 MacArthur’s connection of LV to consumer-resource models
- Inherently unstable consumer-resource interactions
- A consumer-resource framework
- Arguments against resource-based definitions and models
- A simple model of resource partitioning envisions each type of resource available in a community as varying along a “resource spectrum.”
- In 1995, I published an article on humility, and observed that philosophers had neglected that virtue.1
- Consequences of non-logistic resource growth
- Differences in nonlinearity with seasonal resource growth
- Reasons for including resource dynamics
- Behavioral Violence in a Consumer Society
- Resource extinction and quasi-extinction in MacArthur’s model
- The principal judicial organ of the United Nations, the tribunal that has been widely regarded as a potential World Supreme Court, sits at The Hague in a state of august indolence, neglected and underemployed.
- International Studies of Consumer Adoption of E-Banking
- What distinguishes a single resource from others?
- Consumer Protection, Genetic Discrimination, and Privacy — The Issues
- Other Aspects of Climate Change
- A Brief Overview of the Important Aspects