Resource extinction and quasi-extinction in MacArthur’s model
Beginning with MacArthur (1970, 1972), biotic resources in consumer-resource models of competition have usually been assumed to exist at positive abundances regardless of their level of consumption.
This assumption underlies the justification for the common practice of computing LV competition coefficients based on overlap in two consumer’s spectra of resource capture rates (e.g., Vincent and Brown 2005; Leimar et al. 2013), without considering the possibility that one or more resources could be excluded by increased consumer abundance. High consumer efficiency was shown to produce exclusion of one biotic resource in 1-consumer-2-resource models by Holt (1977), who referred to even earlier articles suggesting this possibility. Exclusion of more than one resource may occur via apparent competition when three or more resources are initially present. It is unclear why MacArthur ignored these cases.Complete resource exclusion can be avoided by a low rate of immigration of resources from outside the system being modelled. However, the distinction between systems with low levels of resource immigration and those that are totally closed to immigration is missing from most recent works on competition. This is due to the lack of a metapopulation perspective in non-spatial models of competition. Resources are usually more widely distributed than their consumers because resources can exist in some places where their equilibrium level is too low to support a consumer population. This is likely to cause some input from these resource-only patches into other locations that have consumers. If an analysis of competition only considers locations containing consumers (which is normally the case for studies of competing consumers), such input is most often simply assumed to be absent.
Most natural populations also experience some level of input of consumers, as documented by studies of island biogeography; e.g., Warren et al.
(2015). Having a very low level of immigration of competitors (i.e., consumers) in a LV model only has slight effects on abundances. For example, it changes outcomes of exclusion to presence at very low abundance. Minimal abundance of one species is effectively equivalent to exclusion in its effects on the population levels of other competitors. In cycling systems with heteroclinic (ever expanding) cycles (possible for LV models with three or more species), adding low-level consumer immigration can be essential for persistence (e.g. May and Leonard 1975), but such cases seem to be rare, even in theoretical models. In any event, continued low-level abundance of a competing consumer population that is maintained by immigration should be (and usually is) classified as functional or quasi-exclusion. Low-level immigration of the resources used by those competing consumers often has more profound effects than does consumer immigration. Including a low rate of external resource input can avoid some of the alternative outcomes predicted by spatially homogeneous models that lack this input. The impact of immigration of both consumers and resources is considered here, but the focus will be on the latter.Researchers have explored 2-consumer-2-resource models with biotic resources and low-level resource immigration to examine the effects of quasi-extinction of resources (e.g., Abrams 1998, 2001a, Abrams and Nakajima 2007). However, published work has not compared these to analogous systems without any resource immigration. The models with immigration have shown that greater overlap in resource use may lead to smaller effects of species addition or removal on equilibrium population size over a wide range of consumer overlap values (Abrams 1998). This is contrary to the usual assumption of theory based on the MacArthur model. Figure 6.1A shows how the proportional decrease in population size caused by a competitor can change as the similarity of the two competitors increases.
The unimodal change shown at low mortality rates is due to the fact that, for such efficient consumers, a relatively low consumption rate maximizes population size in a 1-consumer-1-resource system with logistic resource growth. Given an efficient consumer, the resource that is caught less rapidly can support a high enough population of consumer to exclude (via apparent competition) the more rapidly caught resource. If there is resource immigration, this is actually ‘quasi-exclusion, but the very low density of the almost excluded resource has a minimal effect on the population sizes of consumers. Sufficiently similar values of the two capture rates, ci1 and ci2, do not produce resource exclusion in the single consumer system. The range of values of C and d that produces resource exclusion in a 1-consumer system is shown in Figure 6.1B.McPeek’s (2019a) reanalysis of the 2-consumer-2 resource version of MacArthur’s (1970, 1972) model assumed no resource immigration and also assumed that each consumer must come to equilibrium with its resources before the other consumer is introduced, to judge whether coexistence will occur. Such an analysis maybe appropriate in a laboratory system, where complete exclusion is possible in a reasonable time frame (due to small population sizes) and immigration can often be prevented. However, when resource exclusion is a possibility, ruling out immigration drastically changes the nature of competitive relationships and coexistence. Furthermore, the underlying assumption of zero immigration is improbable for most natural systems, because without immigration, any empty patch would remain empty. Movement in
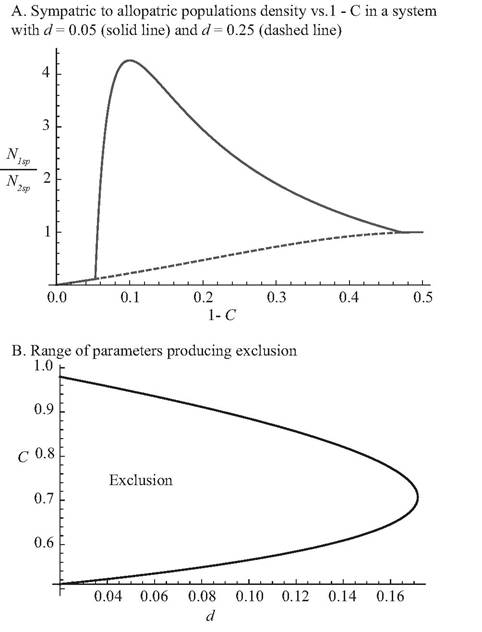
Fig. 6.1 Competitive effects in a 2-consumer-2-resource MacArthur model with the parameters given in the text. These are based on eqs (6.1a), with ‘mirror image' consumers, where C is the capture rate of the 'preferred' resource and 1 - C is that of the non-preferred resource.
Panel A shows the ratio of the consumer population size when present alone to that when it occurs with the ‘mirror image' competitor The solid line assumes d = 0.05, and the dashed line assumes d = 0.25. The smaller d produces apparent competitive exclusion over most of the range of C, while the larger d never does so.Panel B shows the maximum and minimum capture rates, C, of the preferred resource that allow coexistence as a function of the mortality rate, d. The ‘exclusion' zone is characterized by elimination of the earlier arriving consumer by the later arriving competitor when there is no resource immigration and the first consumer comes to its equilibrium density before the second is introduced. The other parameters are r = k = 1, and d1 = d2 = d. Values of d > 0.17157 never result in exclusion. The figure shows that for d < 0.12772, more than half of the possible values of C produce exclusion.some cases may be sufficiently rare that complete resource exclusion is possible, so it is worth exploring the outcomes that are produced in that case. However, McPeek (2019a) did not consider the final outcome of the process of sequential consumer invasion, except to note that it would not be coexistence. The actual result is an outcome that has apparently never been observed, as explained below.
The basic model analysed in this section was introduced in Chapter 3, but immigration of resources is added as a possibility and some changes in notation are also made here. The 2-resource version of this model is given by the following which was used in fig. 6.1 above.
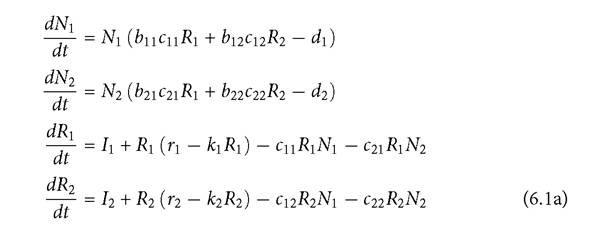
Equations (6.1b), below, represent the 3-resource extension of eqs (6.1a), in which each consumer has an exclusive and a shared resource, the latter being resource 2.
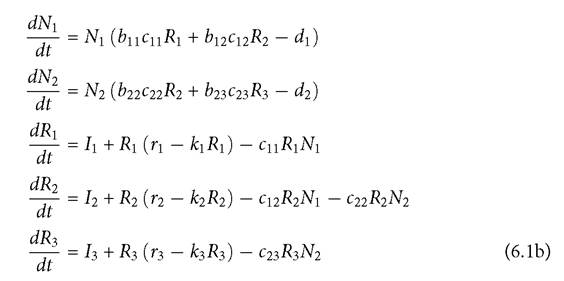
As in previous chapters, resource populations are R, and consumer populations are N.
The maximum per capita growth rate of resource i is ri, and the per capita reduction in that rate with greater resource abundance is given by ki. The per capita consumption rate of consumer i on resource j is cij, and bij is the corresponding resource conversion efficiency. Resource immigration is given by the subscripted I parameters. Comparable models with abiotic resource growth are discussed in Section 6.3. Abrams (1998, 2001a) explored eqs (6.1) but did not consider cases without resource immigration. Other than the presence of resource immigration, the 2-resource version of the model is equivalent to the 2-resource linear functional response model analysed by McPeek (2019a).I will first address the 2-resource case, eqs (6.1a). This provides a good illustration of the expanded range of outcomes arising from full consideration of resource dynamics. To simplify the analysis, assume that both consumers have the same initial mortality, d, and equal conversion efficiencies (all bij = 1, for consumer i and
Negativity, constancy, and continuity of competitive effects • 115 resource j), as well as equal mean consumption abilities. Differences between consumers in their ratios of bij∕bik can affect the outcome of competition (Schoener 1974a; Chase and Leibold 2003). Such differences can promote or inhibit coexistence, but we unfortunately have very few empirical studies of resource conversion efficiencies.
First consider a case in which the two competitors have symmetric partitioning of two resources. The two resources are assumed to have the same growth parameters. The two consumers’ resource consumption rates are mirror images; c11 = c22, and c12 = c21. Without loss of generality, I assume that ci 1 + ci2 = 1 and cii > cij. This allows cii to be replaced by C with 1/2 < C < 1.
The condition for apparent competitive exclusion of the resource consumed more rapidly by a single consumer is that the consumer would achieve a high enough equilibrium abundance on its low-capture-rate resource alone, to exclude the more rapidly caught resource. This requires that C not be too close to either 1/2 or 1. If C is close to 1/2, the resources suffer roughly equal consumption rates, so neither goes extinct. If C is too close to 1, then the consumption rate of the lower-c resource (1 - C) is close to zero, so the consumer population could not achieve a high abundance on that resource alone unless the consumer’s mortality was very close to zero. An intermediate C, slightly above 0.707107 is most likely to lead to resource extinction, as shown in Figure 6.1B.The outcome with resource immigration is coexistence as shown in Figure 6.1A. However, without resource immigration, and assuming complete equilibration of the 1-consumer system before introduction of the second consumer, there may be a different outcome. This is true in those cases where the first consumer causes exclusion of the resource it captures more rapidly. If consumer 1 arrives first and has a low enough mortality, resource 1 will be excluded. When consumer 2 is then introduced, it excludes consumer 1 because of consumer 2’s greater consumption rate of the only resource present (resource 2). Consumer 1 cannot reinvade this system, and the ‘no resource immigration’ assumption means that resource 1 never does so. This outcome is a posteriority effect (i.e., later arriving consumer species always excludes the earlier arriving one). Under the same assumption of no resource immigration, but both resources initially present, simultaneous introduction of both consumers would allow them to attain the 2-consumer outcome. Thus, three outcomes are possible, depending on the timing of consumer introductions.
A second simple scenario of competition under this model (eqs 6.1a) has one generalist (species 1) and one semi-specialized consumer (species 2) that share two resources. The generalist is assumed to have c = 1/2 for each resource, and is assumed to have a slightly larger mortality, d (a cost of generalization). In this case, the single-consumer system with only the generalist consumer always has both resources present. As a result, regardless of resource immigration, the semi-specialist can invade if it has parameters that allow it to increase. Coexistence is relatively easy to achieve when both consumers are introduced while both resources are present; it occurs over a broad range of relative values of di for the two consumers. However, if the semi-specialized consumer 2 is introduced first and allowed to achieve equilibrium with no resource immigration, it will cause extinction of its ‘preferred’ (higher c)
resource. Following this, introduction of the generalist will result in exclusion of the specialist, because the generalist has a much greater c for the one remaining resource. Thus, the outcomes in this sequential introduction case (with no resource immigration) are exclusion of the specialist, if it is introduced first, and coexistence, if the generalist is introduced first or both are introduced while both resources are still present. Resource immigration eliminates the resource extinction that keeps the specialist from returning in the scenario where the specialist is the first consumer present.
Resource exclusion in the no-immigration case becomes more likely (and occurs over a wider range of consumer efficiencies) if the resources themselves compete (Abrams and Nakajima 2007). Resource exclusion also becomes more likely in at least one of the single-consumer systems if the two resources are characterized by unequal r-values. It is necessary that each consumer have a death rate that is low relative to the maximum that allows it to persist in order to have resource exclusion in each of the single-consumer subsystems. Resource exclusion also occurs over a wider range of consumer efficiencies (i.e., d values) if the resources experience relatively little density dependence until they are close to their carrying capacities—e.g., θ-logistic growth with θ >1 (Abrams 1998, 2001a). While the extent of consumer-driven resource exclusion in natural communities is unknown, such exclusion is unlikely to be rare, given the high predator effects shown by experiments on trophic cascades (Shurin et al. 2002; Terborgh 2015), as well as the results of studies that have shown strong apparent competitive effects in systems with a single predator species (reviewed in Holt and Lawton 1994; Holt and Bonsall 2017). In the symmetric 2-resource model considered here, an intermediate level of consumer specialization on the higher-c resource leads to the greatest apparent competitive effect of the lower-c resource on the higher-c resource (Abrams 1998; Figure 6.1B above), and each consumer must have such an intermediate level of specialization for alternative exclusion outcomes to occur in the four-species system.
In a metapopulation with many patches experiencing different environmental perturbations, such variation (or underlying spatial heterogeneity) would increase the probability that resource species arrive via migration, and would increase the probability that different patches would have species occupying different dynamic attractors (assuming they exist). There may be some cases in which the system of interest has extremely low levels of resource immigration, or is totally isolated. In these cases, the alternative outcomes described for the no-immigration cases above may be more likely. It remains to be determined whether any such cases occur in natural systems.
Immigration also plays a key role in determining the outcome of the 3-resource system given by eqs (6.1b), in which each consumer uses one shared and one exclusive resource. This food web (with different resource dynamics) has a long history in competition theory (Schoener 1974a, c, 1976, 1978; Abrams 1975, 1977), but has been ignored in many recent studies (including all of the six focal articles identified in Table 4.1). The articles by Schoener and by Abrams from the 1970s concentrated
on abiotic resources, where resource exclusion is impossible, and resource input is indistinguishable from immigration. However, Abrams (1998) extended this shared- exclusive scenario to biotic resources with low external immigration of all three resources. Consumer extinction in the 2-consumer-3-resource model is impossible when a consumer can maintain a population on its exclusive resource alone. This means that it is possible to have very high overlap in resource use ability (i.e., high relative values of C for the shared resource) and large differences in neutral consumer parameters, but no possibility of consumer exclusion.
The outcomes for the 3-biotic-resource system described in the preceding paragraphs are altered if there is no immigration of resources. If one or both consumers produce apparent competitive exclusion of one resource in the 1-consumer system, that resource is lost permanently. In most of the discussion below, I make the same simplifications as in the above discussion of the 2-resource model; the resources have identical rj, and identical kj, and the consumers initially have identical di. The Bij are again set to unity. The resource utilization rates of the two consumers are again mirror images; c11 = c23 = C (for the exclusive resources, 1 and 3) and c12 = c22 = 1 - C (for the shared resource, 2). In this case, if C < 1/2, the shared resource is the only resource that could be excluded in a single-consumer system, and this outcome occurs under the same conditions as in the comparable 1-consumer-2-resource model discussed above (Figure 6.1b). Exclusion of the shared resource means that there will be no competition between the consumers after the second consumer invades and the two species will coexist if that second species can persist on its exclusive resource alone.
If each consumer eats its exclusive resource at a greater rate (C > 1/2), the exclusive resource in each single-consumer subsystem becomes extinct if d is sufficiently low (and C is not too close to the maximum of 1). When extinction of its exclusive resource occurs with consumer 1 present, the introduction of consumer 2 will always cause extinction of consumer 1. (There can be a neutrally stable line of equilibria after consumer 2's exclusive resource goes extinct, but population fluctuations would likely result in loss of the initially present consumer in most realistic scenarios having environmental variation that affects the two species differently.) Consumer 2's exclusive resource will eventually go extinct in any event, and the final state will be consumer 2 persisting on the original shared resource, which it captures at a rate 1 - C, which is < 1/2. An important feature of this 3-resource system with no resource immigration is that the qualitative outcome is changed by even a small difference from equality in the mortality rates (or a difference between consumers in their consumption rate of the shared resource). In these cases, the alternative outcomes are exclusion of the initially present species when this consumer has a higher R* (= d/C) for the shared resource, or coexistence when the initially present consumer has a lower R* for the shared resource. In both cases, the exclusive resource of the initial consumer goes extinct.
The symmetric 3-resource system with resource immigration was analysed in Abrams (1998). This scenario always allows coexistence ofboth consumers when each consumer is able to exist on its exclusive resource alone. However, as in the 2-resource
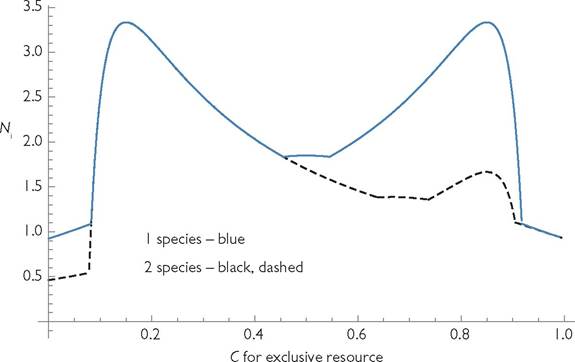
Fig. 6.2 Population sizes in 1- and 2-consumer systems as a function of the attack rate C on the exclusively used resource in a 3-resource system with shared and exclusive resources. The shared resource is consumed at a rate of 1 - C. The death rate of both species is d = 0.075, with b = r = k = 1. The lines are identical when the exclusive resources are extinct.
system, the possibility of quasi-exclusion of resources (one resource maintained by immigration) changes the relationship between similarity and the proportional reduction in population size caused by the competitor. This reduction is shown as a function of consumer similarity in Figure 6.2. As in the 2-resource case, maximal competitive reduction in population size occurs for low levels of similarity, in which each species has a much higher capture rate of its exclusive resource than of the shared resource.
The symmetric 3-resource case just discussed is not the only one in which the outcome of an invasion analysis differs depending on the presence or absence of resource immigration. Consider competition (again using eqs (6.1b)) between a semi-specialized consumer 1 (1 > c11 > 1/2) and a perfect generalist consumer 2 (c22 = c23 = 1/2). Also assume d2 is low enough that the generalist can exist when only resource 3 is present; i.e., d2 < r/(2k). If consumer 1 is the initial resident, it will exclude resource 1 if c11 and d11 lie within the range illustrated in Figure 6.1B. In this case, arrival of consumer 2 excludes consumer 1 provided d2 < d1ZC12. If, for example, consumer 1 is characterized by c12 = 1/4 and c11 = 3/4, then a four-fold or greater death rate disadvantage of the complete generalist consumer 2 is required for consumer 1 to persist after arrival of consumer 2. If the ratio of death rates is smaller than four, then exclusion of a resident consumer 1 occurs following introduction of consumer 2, and coexistence of both occurs when consumer 2 is the initial resident. Resource immigration ensures coexistence of both consumer species regardless of invasion order, as resource 1 will immediately increase following the introduction of consumer 2.
A final scenario in this 3-resource model (with the same resource parameter values and cij + Cji = 1) assumes that the utilization patterns of the consumers are identical but displaced along the resource axis; here c11 = c22, and c12 = c23. Here, I assume that species 1 has a higher attack rate on its exclusive resource, while consumer 2 has a higher attack rate on the shared resource. In a low-d scenario where exclusion of the more rapidly consumed resource occurs with a single consumer, the species having the higher attack rate on its exclusive resource (consumer 1) is easily excluded by consumer 2, when consumer 1 is the initial resident. When consumer 2, with a higher consumption rate of shared resource, is initially present alone, the shared resource is excluded (given a low d), resulting in coexistence with no competition after the first consumer invades. If the scenario is changed to have low- level immigration of both resources, the final state is similar to the final state under no resource immigration and consumer 2 arriving first (i.e., coexistence at unequal abundances). The only difference is a very low abundance of the shared resource due to immigration, but this has little impact on either consumer’s population size.
Coexistence is not the only outcome affected by resource exclusion in the two models described by eqs (6.1a, b). Another feature that is potentially affected is the phenomenon under which continuous directional change in a neutral parameter of one consumer species can produce a discontinuous change in the equilibrium densities of both consumers. (This is impossible in the Lotka-Volterra model with two species.) A discontinuous jump occurs when a resource either goes extinct or regrows from a near-zero density. The cause of this discontinuity is apparent competition, mediated by the consumers. For the systems considered above (eqs (6.1a, b)), agrad- ual change in the mortality of one species frequently produces an abrupt change in the equilibrium densities of both species. Figure 3.2 illustrated the 2-resource system (eqs (6.1a)) as a function of d1 in an example with mirror image competitors having c values of 3/4 and 1/4 (i.e., C = 3/4). The discontinuous drop in abundance to zero for consumer 2 is accompanied by the extinction of resource 1 at the critical d1 (= 1/30). Note that this disappearance occurs when the population of N2 is only approximately 11% lower than its equilibrium when both species have identical death rates and abundances. The loss of consumer 2 is accompanied by a more than threefold jump in the abundance of consumer species 1. The more general condition for discontinuous changes in equilibrium abundances of consumer species as the mortality rate of one competitor is reduced, is d < 2C - 1 in the symmetrical 2-consumer-2 resource model considered above. For example, if C = 3/4, a discontinuous change in the equilibrium of both consumers at a sufficiently low d1 occurs when the initial mortalities of both species are less than 1/2, which is coincidentally half the value above which no consumer can persist. Note that this qualitative result is not altered by a low rate of resource immigration, although immigration changes the N vs d relationship very close to the point of discontinuity. There is a second discontinuity in Figure 3.2, when d1 exceeds 3/10. This is characterized by the loss of both resource 2 and consumer 1, and by a discontinuous jump in the equilibrium density of consumer 2, which more than doubles its population size.
Another lesson from the system described by eqs (6.1a, b) is relevant to ‘modern coexistence theory, which interprets coexistence in terms of equalization and stabilization. The outcomes described above are not consistent with the view that coexistence always occurs when general competitive abilities are equal and there are differences in resource utilization rates (as is implied by at least some versions of ‘modern coexistence theory'). With no resource immigration and temporally spaced introductions of consumers, it is often more difficult for moderately different species to coexist than it is for more similar species. If they do coexist because of resource immigration it is often true that pairs of very different competitors produce greater reductions in each other's population size in sympatry than do pairs of consumers with more similar resource capture rates (Abrams 1998). Increasing the ability of consumer species 2 to obtain its resources, reproduce, or survive (i.e., increasing inequality of the consumers without changing overlap in resource utilization abilities) cannot exclude consumer 1 if the latter can persist on its own exclusive resource(s) alone.
These simple examples also show that ecologists have still not properly understood the role of resource extinction and resource immigration in determining the nature of competitive relationships between consumers. McPeek (2019a, b) calls for more attention to apparent competition, but, by ignoring resource immigration and focusing exclusively on consumer coexistence, his analyses failed to reveal either the posteriority effect or the possibility of coexistence in systems having different single-consumer subsystems, each characterized by the apparent competitive exclusion of one resource. The earlier analysis (Abrams 1998), that included resource immigration, found that a competitor's effect on equilibrium population was greatest for relatively low levels of similarity in resource attack rates, a possibility not mentioned in McPeek (2019a). Results for the 3-resource model (eqs 6.1b) with no resource immigration are also inconsistent with the widespread view that greater similarity always makes coexistence more difficult and greater differences make it easier. Chesson (2018, supplemental material) defends a Lotka- Volterra approximation for understanding consumer-resource models in spite of the existence of resource extinction with the following statement: ‘(A56) [an LV equation derived from MacArthur's consumer-resource equations] would still represent valid Lotka-Volterra dynamics, but their interpretation in terms of specific resources would be compromised by resource extinction'. This (correct) claim is problematic because almost all the conclusions that may be of ecological interest (including coexistence or not) change abruptly when one or more resources go extinct.
Both the LV model and MacArthur's flawed analysis of his corresponding consumer-resource model have led to the conclusion that competitive effects are decreased and the ability to coexist is increased by having a greater difference in resource attack parameters (i.e., reduced overlap in resource utilization, often termed ‘niche overlap'). The preceding results show that neither of these generalizations is valid in the case of two very efficient competitors that each uses two resources. Models having more consumers and/or more resources also fail to support these generalizations (Abrams et al. 2008a; Abrams and Rueffler 2009).
Apparent competition is the cause of the resource extinctions noted above. However, in a 2-consumer-2-resource system with no direct consumer density dependence, apparent competition (i.e., shared predation) is functionally absent over a wide range of parameter perturbations. The Yodzis (1988) method of measuring indirect effects (embodied in eqs (3.1) and (3.2)) implies that the total apparent competitive effect of one resource on the other can be measured by the population response of resource j to a change in the neutral parameter, ri of resource i in eqs (6.1a). However, this effect is zero in the 2-resource system when there is no self-limitation in either consumer species. Self-limitation is assumed to be absent in eqs (6.1a) and most published analyses of consumer-resource models of competition and apparent competition. In its absence, apparent competition does not produce any effect on equilibrium abundance provided that all resources and consumers are present. Small to moderate change in the ri of one resource species will not affect its own abundance or the abundance of the other resource—both resource abundances (assuming they are positive) are entirely determined by the equilibrium requirements of the two consumers. However, a sufficiently large parameter change affecting one resource in this 4-species system will produce extinction of one of the consumers. This will often be followed by extinction of one resource, depending on the efficiency of the remaining consumer. Just as in the case of between-consumer interactions in this 2-consumer-2-resource system, apparent competitive effects become discontinuous, with the equilibrium resource population jumping from a significant density to zero with a very small (growth-increasing) change in the parameter. Other implications of apparent competition in subsystems of this simple model, and in models with more resources are treated in Abrams (1998, 2001a, 2009b), Abrams and Rueffler (2009), and Abrams et al. (2008a).
Although it does not involve resource extinction, it is worth examining one of the possible outcomes of having a third consumer in the 3-resource system described by eqs (6.1b). The third species is a specialist on the shared resource (R2), which means that its presence fixes the abundance of this resource at d32∕(b32c32). For consumer species 3 to exist, this resource requirement must be lower than the equilibrium R2 that exists in the absence of consumer 3. Once it is present, species 3 eliminates any competition between consumers 1 and 2, as neither of these consumers affects the equilibrium abundance of the only resource they share. Thus, even without resource exclusion, the 3-consumer-3-resource MacArthur system shows that competition coefficients are not independent of the presence or abundance of additional consumer species, even when those species do not bring about resource extinction. In other words, the multi-species MacArthur model is again inconsistent with the Lotka-Volterra model.
The limitations of many previous analyses of the MacArthur model (eqs (6.1a, b), and examples having more species on one or both levels) go beyond the failure to take resource extinction into account. Many conclusions arising from the model depend on the exact linearity of its component per capita growth rate functions. Nonlinearity in resource density dependence results in very different competitive interactions, as does competition between the resources themselves. These possibilities are discussed below.
6.3
More on the topic Resource extinction and quasi-extinction in MacArthur’s model:
- 3.4 MacArthur’s connection of LV to consumer-resource models
- A simple model of resource partitioning envisions each type of resource available in a community as varying along a “resource spectrum.”
- Causes of Extinction
- Extinction is the end point of incremental biological decline
- Parasites can drive host populations to extinction
- Extinction and colonization rates often vary among patches
- Species richness is a balance between immigration and extinction
- Fluctuations in population size can increase the risk of extinction
- The diversity of life reflects both speciation and extinction rates
- Extinction of Usufruct
- Population Extinction
- Extinction of Praedial Servitudes
- A. Creation and Extinction of Secured Interests