Who interacts with whom makes all the difference in community membership
Even if species can disperse to a community and cope with its potentially restrictive abiotic conditions, the final cut to community membership is coexistence with other species (see Figure 19.4C).
Clearly, if a species depends on other species for its growth, reproduction, and survival, those other species must be present if it is to gain membership in a community. Equally important, some species may be excluded from a community by competition, predation, parasitism, or disease. For example, returning to our thought experiment, we might assume that lakes are suitable habitats for many fish species, but could those species all live together in one lake, given that resources are limiting? A simple view suggests that the best competitors or predators should dominate the lake, thus excluding weaker competitors and resulting in a low-diversity community. But we know that most communities are full of species that are actively interacting and coexisting. So what allows this coexistence? There are many important mechanisms that allow species to coexist, and we will spend the next two sections considering them. But first, let's ask how species might be excluded from communities by biological interactions—a question that is a bit different, but equally relevant.The invasive species literature provides some of the best tests of whether species interactions can exclude species from communities. The failure of some nonnative species to become incorporated into communities has been attributed to interactions with native species that exclude, or slow the population growth of, the non-native species—a phenomenon that ecologists call biotic resistance. Multiple studies in a variety of communities have shown that native herbivores have the ability to reduce the spread of non-native plants in substantial ways. Maron and Vila (2001) found that mortality of non-native plants due to native herbivores can be quite high (about 60%), especially at the seedling stage (up to 90% in some studies).
But while native herbivores can kill individual non-native plants, it is still unknown how important native species are in completely excluding non-native species from a community. For example, Faithfull (1997) found that in Australia, adults and larvae of the native lucerne seed web moth (Etiella behrii) breed and feed on the seedpods of the invasive gorse shrub Ulex europaeus, but the plant still continues to spread (FIGURE 19.6). This lack of knowledge about biotic resistance may be an artifact of ecologists being more likely to study why a particular non-native species does or does not spread once it becomes a provisional member of the community than to study all the cases in which it is unable to gain a foothold because of interactions with native species. It may also be true that most failed introductions of non-native species go completely undetected.
FIGURE 19.6 Stopping Gorse Invasion? Herbivory by adults and larvae of the native lucerne seed web moth (Etiella behrii) has slowed, but not stopped, an invasion of the non-native gorse shrub Ulex europaeus (the plants with yellow flowers) in Australia. View larger image
Climate Change Connection
How are Species Invasions Enhanced by Climate Change?
There is growing evidence that climate change—and, in particular, rising temperatures—may facilitate the invasions of species that would be unable to survive under cooler conditions. As you might guess, climate change can play a role in mediating the ability of species to pass through the three filters described in Figure 19.4, thus potentially exacerbating the arrival, spread, impact, and management of invasive species.
Hellmann et al. (2008) outline five potential consequences of climate change for invasive species (FIGURE 19.7A). The first consequence arises when climate change alters the pathways (transport and introduction) of non-native species (see Figure 19.7A, consequence 1).
Such alterations could occur if climate change better links areas that are geographically separate prior to climate change. For example, Sylvia Behrens Yamada and colleagues (2005) have shown that non-native European green crabs (Carcinus maenas) (FIGURE 19.7B), introduced to San Francisco Bay from the east coast of North America, were able to colonize northern Pacific estuaries during warm El Nino years. At these times, crab larvae were transported in stronger, warmer northward-flowing coastal currents to new estuarine locations in Oregon and Washington, where they were able to survive as adults. Thus, changes in coastal currents caused by global warming could create new pathways of dispersal for other non-native species.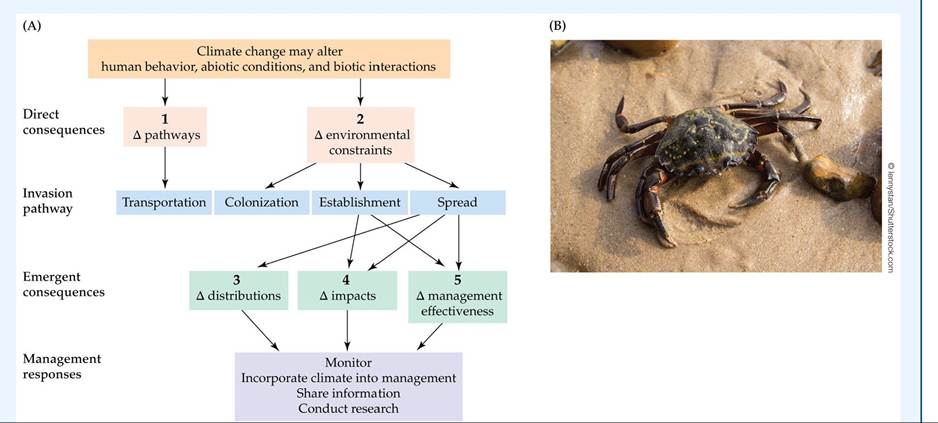
FIGURE 19.7 The Five Consequences of Climate Change for Species Invasions
(A) Consequences 1 and 2 directly affect the invasion pathway for new non-native species. Consequences 3, 4, and 5 are emergent after an invader has become established and spread, and they have management implications. Delta (Δ) means “change in.” (B) The European green crab (Carcinus maenas) has invaded estuaries along the U.S. Pacific coast. (A after J. J. Hellmann et al. 2008. Conserv Biol 22: 534-543.) Vi6W larger image
A second consequence of climate change is the alteration of environmental constraints on non-native species that allows some species to overcome physiological or biotic constraints on their persistence outside of their native range (see Figure 19.7A, consequence 2). For example, in the green crab invasion mentioned above, it was predicted that green crabs would become locally extinct in northern estuaries once El Nino subsided, because of their intolerance of cold winter ocean temperatures (they are unable to molt and reproduce at temperatures below 10°C). In fact, the researchers found that green crabs persist as invaders where they experience occasional warm winters, during which they have much greater survival, growth, and reproduction.
A third consequence of climate change is the alteration of the distributions of existing invasive species (see Figure 19.7A, consequence 3). For non-native species that have gained a foothold outside of their biogeographic range, climate change could expand (or contract) their new range in dramatic ways. For example, one could imagine that with warming of estuarine water temperatures above 10°C, green crabs would not merely exist in small populations but, instead, increase their numbers through enhanced survival and reproduction.
The fourth consequence of climate change occurs when the impacts of non-native species are altered (see Figure 19.7A, consequence 4). The impacts of the green crab under the climatic conditions of today are minimal. Hunt and Behrens Yamada (2003) observed very little overlap in the distribution and resource use of the green crab and the larger native red rock crab (Cancer productus). The red rock crab is dominant in the colder and more saline portions of estuaries, while the invasive green crab occurs in warmer and less saline areas. With climate change, increased temperatures or more rainfall could lead to warmer and less saline estuarine conditions, thus favoring green crabs over red rock crabs and having a greater impact on the estuarine community as a result.
The fifth and final consequence of climate change is its effects on the management of non-native species (see Figure 19.7A, consequence 5). Current management, whether it involves removal of invasive species or restoration of habitats impacted by these species, will need to adapt to changing climate in ways that maintain its control and efficacy. In the green crab example, management has been minimal beyond destroying individuals found in traps. If green crabs expand in population size and range in response to climate change, however, active management may be required to keep this invasive species from becoming a pest to shellfishery and aquaculture operations. As you can see, climate change can act on invasive species in a multitude of ways that may be hard to predict.
Studying invasions gives us valuable insights into how species are included or excluded from communities, but how species coexist can be complicated. In the next two sections, we will consider theories of species coexistence and ultimately species diversity. We will start by revisiting the concept of resource partitioning (also known as niche partitioning), which relies on ecological and evolutionary “compromises” that result in divergence in resource use as a mechanism for coexistence (see Concept 14.2). We will then explore alternative theories and studies that consider the importance of disturbance, stress, predation, and even positive interactions to the coexistence of species and, ultimately, the species diversity of communities.
More on the topic Who interacts with whom makes all the difference in community membership:
- Reviewers
- Community-Academic Partnerships
- Exploring Transcendent Communication
- Musgrave’s Late Writing
- Human beings live in groups.
- Designing With a Dialogical Sensibility
- SEVEN WORKSHOP MODULES
- Narrative
- WORKING WITH MORAL CONFLICTS
- APPROACHES TO ADDRESSING INTRACTABLE CONFLICT: FIVE PARADIGMS