CONTRIBUTION OF VITAMIN D ON PROTECTION/RISK TO HIV INFECTION AND DISEASE PROGRESSION TO AIDS
Both direct effects on HIV replication and indirect effects through innate and adaptive immunity responses affect the complex interactions of vitamin D endocrine system on HIV infection (Table 3).
Table 3: Contrasting effects of Vitamin D on HIV infection and progression to AIDS.
| Vitamin D confers protection by: | Vitamin D confers risk by: | |
| Physiological evidences | Promoting polarization to Treg that facilitates a proper modulation of the initial response | Impairing Th1 cellular immune response |
| Promoting a tolerogenic status that favour viral persistence | ||
| Inducing defensin synthesis | Interfering with APOBEC3G synthesis Increasing viral replication Lower levels of CD4+ T cells and the mean | |
| Environmental evidences | Solar irradiation favours immune modulation | CD4+/CD8+ ratio in HIV+ patients during summer and spring |
| High steady-state prevalence of HIV infection in Southern European countries | ||
| Less functional VDR-allelic variants that are | ||
| Genetic evidences | associated with protection to HIV infection and slow progression to AIDS |
Action of Vitamin D on HIV Replication may Contribute to HIV Infection
Conflicting data have been reported concerning the effects of 1,25(OH)2D3 on HIV-1 replication in primary monocyte-derived MAC as well as myeloid cell lines; both enhancement and inhibition of HIV-1 replication in response to 1,25(OH)2D3 have been reported.
Studies of the role of 1,25(OH)2D3 in HIV-1 infection have been hampered by technical and methodological variability, as well as lack of homogeneity in the experimental strategies. Several parameters can have deep influence on the results: cell types, 1,25(OH)2D3 concentration, viral strains (X4 vs. R5), 1,25(OH)2D3 treatment before or after infection, co-treatment with other stimuli (such as TNF-α), chronic vs. acute infection, days after infection for viral load quantification or infectious dose.The main used models are cell lines of myeloid origin, although several authors have also analyzed the 1,25(OH)2D3 effects on primary cells.
HL-60 cell line was established from a patient with promyelocytic leukaemia. This cell line acquires morphology and markers of mature granulocytes in the presence of DMSO, retinoic acid, or cAMP, whereas differentiates into a MAC-like phenotype after 1,25(OH)2D3 or phorbol ester (TPA or PMA) stimulation. HL- 60 cells are CD4+/CXCR4+/CCR5- and consequently are resistant to R5 but susceptible to X4 HIV-1 strains [150]. However after 1,25(OH)2D3 differentiation (10-7 M) for 5 days, this cell line becomes susceptible to HIV-1JR-FL or HIV-1BaL (R5). In this study, the TPA also rendered the cells susceptible to R5 viruses; this observation correlates with the observed induction of CCR5 after PMA treatment of HL-60 [151]. There was no effect of 1,25(OH)2D3 or TPA on HIV-1JR FL virus production from HL-60 cells when treated immediately after infection, suggesting a delay time between the stimuli addition and the susceptibility to viral infection. In a model of chronically infected HL-60 cell line (HIV-1JR-FL), 1,25(OH)2D3 enhanced virus production more than 10-fold and induced the same phenotypic changes as observed in uninfected cells. HL-60 cells treated with 1,25(OH)2D3 (10 nM) showed increased CXCR4 mRNA expression after 3 days of stimulation [152,153] and 1 mM of 1,25(OH)2D3 increased the viral production of the HTLViiib strain (X4) [150].
On the other hand in HL-60 cells a 30% inhibition of CXCR4 surface expression has been reported after 3 days of 1,25(OH)2D3 stimulation at 600 nM without any change in mRNA amount [154]. However, although CXCR4 surface expression was decreased upon treatment with 1,25(OH)2D3, it enhanced the functional G-protein coupling to the receptor as demonstrated by the increase in SDF-1-mediated GTP binding [154].The U937 cell line was established from a histiocytic lymphoma [155]. This cell line shows a phenotype of myelo-monocytic lineage (CD13+, CD15+, and CD33+). Naive U937 cells, as HL-60 cells, are CD4+/CXCR4+/CCR5- being vulnerable to infection by X4 strains but resistant to R5 strains [156]. U937 cells exposed to 10-6 M 1,25(OH)2D3 for 1 or 2 days prior to infection with HIV-1bru (X4) displayed 5-fold increase in virus replication; however, shorter 1,25(OH)2D3 stimulation reduced the levels of viral replication and the simultaneous infection plus 1,25(OH)2D3 treatment (10-8 M) produced a 75% reduction in the viral load after 3 days of infection [157]. Skolnik etal. [158] reported 140-fold increase in viral replication in U937 cells pretreated with 24 nM 1,25(OH)2D3 for 1 day before infection. Opposed results were communicated by Kitano et al., showing that a regime of 10-7 M 1,25(OH)2D3 for 5 days prior to infection totally inhibited X4 replication and had no influence on the R5 strains HIV-1 and HIV-1BaL (no infection at all) [150]. The role of 1,25(OH)2D3 in modulation of HIV-1 production in chronically infected U937 cell line was in agreement with the data reported in HL-60 cell line; 1,25(OH)2D3 at 1mM induced an elevated release of infectious viral particles (14-fold) after 6 days in culture. The authors observed no differences in the amounts of endogenous TNF-α production in control and 1,25(OH)2D3 treated cells, although there was a clear upregulation of the TNF-α receptor [18].
X4 chronically infected U937 cells, named U1 cells, produces HIV- 1 particles only after induction with several stimuli such as IFN-γ, IL-6, GM-CSF or TNF-α. The 1,25(OH)2D3 treatment alone (10-8 M for 5 days) did not induce any increase in virus replication but cotreatment with TNF-α resulted in 3-fold increase in virus production above that induced by TNF-α alone. In clear contrast, 1,25(OH)2D3 blocked the viral replication induced by INF-γ, IL-6 and GM-CSF [159]. The so- called plus clones (UC12, UC14) supported viral replication of X4 strains more efficiently than original U937 cell line; in contrast the minus clone (UC11) showed only very limited viral replication [160,161]. The origin of this phenotype is controversial; it appears to be related at least to the density of CXCR4 in cell membrane, and lower in minus clones, although there were no clear differences in mRNA levels and functional Ca++ mobilization in response to CXCL12 between the two cell types [162]. Remarkably, stimulation with 1,25(OH)2D3 upregulated HIV-1 X4 replication in minus clones to the levels comparable to those observed in plus clones, however the plus clones did not show any changes in response to the treatment. 1,25(OH)2D3 selectively induces a slight increase in CXCR4 expression on the cell surface in minus clones [163].The A3.5 cell line was established from human bone cell cultures. This cell line shows a monocyte phenotype (HLA-DR+/CD15+/CD3-) and is susceptible to infection by X4 and R5 HIV-1 strains [158]. A3.5 cells exposed up to 240 nM 1,25(OH)2D3 for 1 day prior to infection with HIV-1iiib (X4) exhibiting enhanced virus replication in a dose-dependent manner; augmentation was up to 104 fold in the presence of 240 nM 1,25(OH)2D3 after 14 days of culture.
THP-1 exposed to 10-7 M 1,25(OH)2D3 for 5 days prior to infection with both R5 strains HIV-1jr-fl and HIV-1BaL and X4 strain HIV-1iiib, exhibited a boosted viral replication after 7 days of culture.
The effect of 1,25(OH)2D3 treatment on HIV-1 infection of primary monocyte-MAC has been evaluated in three different studies.Skolnik et al. [158] observed up to 12- fold enhancement of R5 replication in cultures treated with 1,25(OH)2D3 (10-8 M) after 14 days post-infection.
In contrast, Schuitemaker et al. [165] reported that primary monocyte derived MAC treated for 5 days with 1,25(OH)2D3 (10-8 M) were markedly protected from HIV replication. Finally, Pauza et al. [157] observed large variability and contradictory results in 1,25(OH)2D3 effects between MAC from different blood donors; in some cases the treatment produced a 50% reduction in viral replication but in others it induced a slight increase in viral production.
Another experimental strategy to evaluate the effects of 1,25(OH)2D3 signalling on HIV-1 replication parameters has been done by an evaluation of HIV-1 LTR promoter activity after 1,25(OH)2D3 stimulation in transfected HeLa, Cos-1 and U937 cells. 1,25(OH)2D3 stimulated the LTR transcription in a dose-response manner, with 4-fold induction with 100 nM 1,25(OH)2D3 [17]. We have observed the same effect in primary monocyte- derived MAC from healthy blood donors and more interestingly, the intensity of the induction showed a relationship between VDR BsmI genotype polymorphism (unpublished data from AC laboratory).
Data regarding the significance of 1,25(OH)2D3 in the course of HIV-1 infection are scarce; Arpadi et al. [10] evaluated the effect of 1,25(OH)2D3 supplementation in children and adolescents at a dosage of 105 IU every 2 months and they have found no difference in progression of HIV-1 disease as measured by CD4+ cell count, viral load, rate of disease progression or antiviral treatment failure.
In conclusion, the 1,25(OH)2D3 effects on viral replication depend on several parameters, such as the maturation stage of the cells, the timing of treatment and probably individual genotypes for VDR and the genes coding for enzymes regulating the metabolism of 1,25(OH)2D3.
Additional efforts will be necessary to unveil the specific role of 1,25(OH)2D3 signaling in HIV-1 replication and receptors expression especially in primary cells of monocyte/MAC lineage. The upregulation of CXCR4 by 1,25(OH)2D3 in cells of myeloid lineage has been demonstrated by several studies [163], although in this context the evaluation of the influence of VDR genotypes on the in vivo selection of X4 or double tropic R5X4 strains is warranted. MAC are capable to perform both hydroxylation steps of the 1,25(OH)2D3 metabolism suggesting a possible role of local 1,25(OH)2D3 synthesis by myeloid cells, although we cannot exclude the possibility that 1,25(OH)2D3 is already present in these cells, especially after an initiation of differentiation process, pathogen encounter or cytokines stimulation. A promising experimental strategy could be to test the effects of antagonists of 1,25(OH)2D3 or siRNA against VDR during the course of HIV-1 infection of primary MAC as well as the use of steroids deprived of serum (charcoal-dextran treated) or ketoconazole (24 hydroxylase inhibitor).Action of Vitamin D on Innate Immune Response has Contrasting Effects on HIV Infection
The most well characterised antimicrobial peptides in humans are defensins and cathelicidin (also named LL-37).
Defensins belong to the family of microbicide and cytotoxic peptides involved in innate defense. They are abundant in the granules of neutrophils (5% of total protein content) and are also found in epithelia of mucosal surfaces such as those of the intestine, respiratory tract, urinary tract, skin and vagina. Members of the defensin family are highly similar in protein sequence and distinguished by a conserved cysteine motif.
α-defensins and most of the β-defensins genes are clustered on chromosome 8p23.1 locus that exhibits copy number variation from 2 to 12 copies per diploid genome. β-defensins are widely expressed throughout human epithelia. More than 30 loci exist, but only 4 (HBD1 to HBD4) have been studied in detail. α- defensins 1-4 and β-defensins 2-3 show anti-HIV-1 activity in vitro. In vivo expression correlates with protection in mother-to-child transmission, as demonstrated by the strong association between high α- defensin concentrations and a decreased risk of partum and postnatal HIV transmission has been observed. On the contrary α-defensins 5 and 6 increase HIV-1 infectivity and may play a role in enhancing HIV-1 transmission through genital mucosa [166]. The relationships between vitamin D and defensin expression have not been studied in detail. One report described the upregulation of defensins expression in mouse skin after UV stimulation mediated by vitamin D production [167]; in humans the defensin-β2 promoter contains a VDRE, and the gene transcription can be slightly enhanced by 24 hours treatment with 1,25(OH)2D3 (1,5-2,0 fold) in epithelial cells but not in neutrophils or monocytes. On the contrary, 1,25(OH)2D3 partially blocks the defensin-β2 expression at 48 hours in head and neck squamous carcinoma cells and adult keratinocytes [49].
In humans there is only one cathelicidin gene that is expressed on all epithelial surfaces (mouth, tongue, esophagus, lungs, intestine, cervix and vagina, sweat glands, etc.) and circulating white cells, including neutrophils, monocytes, natural killer cells, and γδT cells. In general, the expression of cathelicidin in most epithelial sites is constitutive but can be increased by local injury or infection. The interaction between vitamin D signalling and cathelicidin expression has been described in detail; cathelicidin gene is a direct target of VDR, because its promoter contains a VDRE and vitamin D treatment induces a strong up-regulation of cathelicidin mRNA. Cathelicidin is able to block HIV-1 infection in vitro [168, 169] but the concentration required to reach 50% of inhibition of infection (30-50 mg/ml) is more than 2000- fold higher than the mean concentration observed in Cervicovaginal secretion (13 ng/ml, range 2-191 ng/ml) [170]. The same study observed that the presence of sexually transmitted infections was associated with higher risk of HIV-1 acquisition (especially N. gonorrhoeae, C. trachomatis or both) and increased levels of several defensins and cathelicidin.
Cathelicidin is an agonist of the formyl peptide receptors and induces increased cell migration, phagocytosis and release of proinflammatory mediators [171], being the neutrophils, monocytes, and T cells the main target cells; this peptide has also been related to neovascularization and wound healing [172]. We hypothesise that although cathelicidin shows minor antiviral effect at physiological concentration [168-170], it might induce a more permissive environment for HIV-1 replication through attraction of neutrophils, monocytes and T cells to the tissues exposed to viral infection [170]. New data concerning the functional relationship between antimicrobial peptides levels, HIV-1 susceptibility and polymorphisms in VDR or the enzymes of vitamin D synthesis and degradation might provide a more accurate model of the complex interaction between this arm of innate immune response and HIV-1 infection. In addition to microbicide and cytotoxic peptides, host factors with specific antiviral properties have been described. Among them, the human apolipoprotein B mRNA-editing enzyme-catalytic polipeptide-like-3G (APOBEC3G) has raised special interest as an interfering agent of HIV dissemination. APOBEC3G belongs to a family of cytidine deaminases that, in absence of the viral protein Vif, is incorporated in the nascent viral particle. Virions carrying APOBEC3G express cytidine deaminase activity during reverse transcription that induces deamination of cytosines to uracil, resulting in G to A hypermutation on DNA positive strand. Hypermutated viral DNA leads to aberrant viral products aborting viral propagation [173]. Antiviral APOBEC3G activity can be abrogated by the interaction with viral protein Vif that induces proteasoma-dependent degradation of APOBEC3G. Studies on exposed uninfected individuals (EUI) indicate that variations on APOBEC3G levels might influence vulnerability to HIV infection. APOBEC3G expression is up-regulated by IFN-α and IFN-γ, induced in response to viral exposure [174]. Expression levels of IFN-γ vary among T-helper subtypes, being mainly expressed after Th1-polarization. Following this, it has been shown that CD4+ T-helper type 2 lymphocytes express lower levels of APOBEC3G than T-helper type 1 lymphocytes. As a result, differences on infectivity among Th1- and Th2-produced HIV particles have been correlated with changes in APOBEC3G expression levels [175]. It is tempting to speculate that the 1,25(OH)2D3-mediated inhibition of IFN-γ production [63] contributes to inhibition of Th1 polarization and the subsequent displacement to a Th2 profile that could be reflected as a lower expression of APOBEC3G which favours the not restricted Th2- produced HIV particles. Evaluation of APOBEC3G response on CD4+ T cells to 1,25(OH)2D3 treatment could be of interest to assess this hypothesis.
Vitamin D Action on Acquired Immune Response: Tolerogenic vs. Activation Strategies on Persistence of HIV Infection
Although immunopathogenesis of HIV infection has been largely studied, mechanisms underlying viral persistence and inability of host immune response to control infection remain obscure. T cell immune responses, that are capital for the control of many viral infections, are often unable to control HIV replication. The incapability to mount an efficient T cell response seems to be a common feature of chronic viral infections, such as HIV, hepatitis virus B (HVB) and C (HVC) infections. It has been found that viruses causing persistent viral infection develop a variety of strategies to evade or overcome the host immune response. Among mechanisms developed on such a way, avoiding T cell activation by the promotion of a tolerogenic status [176] and induction of T cell exhaustion by T cell overstimulation [177], there are two strategies used by HIV to evade an efficient T cell mediated immune response. We can expect contrasting effects of vitamin D immunomodulation on these two mechanisms.
HIV infection is characterised by several disturbances in the cytokine expression pattern. Among these, a general decrease in the expression of pro-inflammatory cytokines, as a consequence of a reduced Th1 polarization, seems to contribute to the host-inability for mounting a proper T cell mediated response [107]. It is remarkable that vitamin D and HIV infection exert similar action on Th1- Th2-polarization, both of them favouring Th2 polarization. As for vitamin D action on Th cell polarization, HIV infection is characterised by a down-regulation of IL-12 and IFN-γ as a result of the interaction of HIV-1-specific T cells with APC inducing CTL dysfunction [178-180]. IL-12 is a key cytokine required for the correct priming of CD4+ T cells to provide help to CD8+ T cells to respond to HIV infection [181]. In addition, as for vitamin D mediated induction of IL-10, HIV Tat protein, a key viral protein involved in viral replication, interferes the host cytokine production by the induction of IL-10 [182]. IL-10 is up- regulated during HIV infection, contributing to maintain a tolerogenic status that favours HIV mediated immunodeficiency. Finally, it has been proposed by several authors that the Th2-polarization that characterises HIV infection could resemble an allergic response to viral antigens [183,184] and that gp120 envelope and Tat viral proteins could act as potent viral allergens [185-188]. Also, a high frequency of hypersensitivity reactions in HIV infected patients has been described to contribute to HIV disease progression [184]. As formerly mentioned, vitamin D promotes allergy [115] and predisposes to allergic syndromes [116]. Following this, vitamin D could contribute to reinforce the HIV-promoted allergy associated with disease progression.
Despite the presence of HIV induced immunodeficiency, almost all cellular components of the immune system, B cells, NK cells, T cells and MAC show evidence of immune activation. Early steps of HIV infection are characterised by an acute T cell response, mainly CD8+ T cells, which can be maintained during the chronic phase of infection, and with a lower magnitude CD4+ T cells, probably due by their preferential depletion by the virus [189]. In addition, microbial products, mainly derived from the gastrointestinal tract, contribute to HIV-related systemic immune activation [190]. In spite of this T cell response, HIV infection can be established and reach successful persistence. Paradoxically, T cell activation, that may be considered a positive response against viral pathogens, could be detrimental in the context of HIV infection, as new targets for viral replication, in particular CD4+ T cells, can contribute to promote HIV infection. This intuitive observation is supported by experimental data indicating that there is indeed a direct correlation between T cell activation levels and HIV disease progression [191,192]. This paradoxical effect of immune activation is exploited and promoted by HIV in a perverse strategy [193,194], inducing an altered homeostasis of T cells (i.e., T cell turnover and apoptosis). Continuous and persistent differentiation of naive T lymphocytes to antigen experienced cells and subsequent apoptosis reach boundaries on the regenerative capacity of the immune system. As a consequence HIV infected individuals lose their capability to replenish naive T cell pool, exhibiting characteristics of replicative senescence [195] that exhaust the capacity of the immune system to control HIV infection [177].
It has been proposed by several authors that suppression of the activated immune response in HIV infection can contribute to control infection. In fact, antiretroviral therapy (ART), which is until now the best strategy to combat HIV, inhibits virus replication by contributing to the deactivation of the immune responses in HIV infected individuals [196]. Treatment of HIV infection with immunosuppresent drugs as ART coadjuvants has been considered, assuming that a decreased immune activation associated with HIV infection may be beneficial [197]. However, concerns have been raised about this strategy; changing initial parameters may have effects on long-term infection, by establishing a new immunological set point that may affect the rate of disease progression [198].
It is intuitive to consider vitamin D treatment as an immunomodulatory adjuvant in HIV infection. As previously mentioned, vitamin D acts on T cell polarization promoting Treg cells while inhibiting T effectors cells, mainly Th1 cells. Thus, vitamin D may contribute to avoid the early exacerbating T cell response that HIV could have taken in advantage. Nevertheless, the contribution of Treg on HIV infection must be considered with caution, as beneficial and detrimental effects are expected [199]. In support for a beneficial effect of Treg on HIV infection is the negative association between Tregs and immune activation showed during HIV infection [200] that may contribute to the attenuation of HIV-specific T cell immune responses in the early stages of HIV disease [201]. On the other hand, in support for a detrimental effect is an association of Treg cells with more severe stages of infection and induction of Th2 polarization [202]. An increase in Treg function could impair cellular immunity that is able to block HIV spread. In the simian immunodeficiency virus (SIV) infection a premature induction of immunosuppressive regulatory cells has been shown to contribute to viral persistence by limiting early antiviral response [176].
Another concern about using vitamin D as an immunosuppressant agent on HIV infection comes from the expected promotion of T cell exhaustion and viral persistence induced by vitamin D, as consequent to an altered pattern of cytokines expression. As pointed before, vitamin D induces Th2 and Treg cells, with a IL- 4/IL-5/IL-10 and TGF-β∕IL-10 profiles. IL-10 is an immunomodulatory cytokine that attenuates inflammatory response by suppressing Th1 cytokine production and proliferation of CD4+ and CD8+ cells [203]. Recently, a key role of IL-10 cytokine on T cell exhaustion and viral persistence has been proposed [204]. Increased IL- 10 production has been reported during persistent viral infection of lymphocytic choriomeningitis virus (LCMV), whereas anti-IL10 receptor antibodies unleash the viral persistence [205]. Thereafter, IL-10 receptor antagonists have been proposed for the treatment of chronic viral infections in humans [206]. Thus, vitamin D-induced IL-10 production could contribute to virus persistency and favouring viral persistence.
Lesson Learned from Vitamin D Receptor Polymorphisms and Association Studies
Several VDR polymorphisms have been described in the regulatory, coding and 3’ UTR regions with functional effects (Fig. 8). Two common polymorphisms, Cdx (rs1568820) and A1012G (rs4516035), have been located in the 5’ regulatory region that influence binding of transcription factors. The Cdx polymorphism is a G to A transition located between exons 1f and 1e that alters the recognition site for the intestinal-specific transcription factor caudal-related homeodomain protein (Cdx)-2 affecting 1,25(OH)2D3-VDR mediated intestinal calcium absorption [207]. The A1012G polymorphism is an A to G transition located between 1e and 1a exons that modify the GATA binding protein (GATA)-3 transcription factor recognition sequence, involved in the regulation of Th2 polarization. Cdx-2 and GATA promoter SNPs are included in single haplotype block of the VDR promoter region [208].
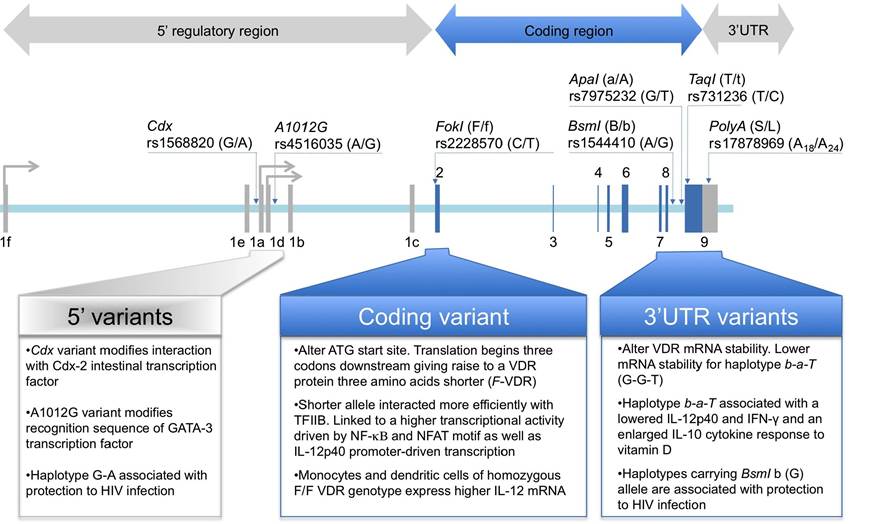
Figure 8. The most relevant VDR polymorphic markers are all along VDR gene structure. Two markers are located in the promoter-5'UTRs region, rs11568820 (Cdx) upstream exon 1e and rs4516035 (A1012G) between exons 1e and 1a. Marker rs2228570 (FokI) is located in exon 2. Finally, four additional markers are located in the 3’UTR region, rs1544410 (BsmI) and rs7975232 (ApaI), between exons 8 and 9 and rs731236 (TaqI) and rs17878969 (PolyA) in exon 9. SNP markers rs11568820, rs4516035, rs2228570, rs1544410, rs7975232 and rs731236 are biallelic. Marker rs17878969 is a mononucleotide repeats of adenines, showing a bimodal distribution of allele frequency [213] with short (S) allele expanding 18 A repeats and long (L) allele expanding 24 A repeats. Alleles have been coded according to coding VDR sequence. Restriction fragment length polymorphism (FokI, BsmI, ApaI and TaqI) alleles are also coded according to cutting (small letter) and un-cutting (capital letter) alleles. Functional effects of polymorphic variants on immune function are depicted at the bottom of the Figure.
In addition, a common FokI polymorphism (rs10735810) has been described in the VDR coding region that alters the first ATG start site to an alternate ACG sequence. By this polymorphism two potential starting sites for the VDR translation seems to appear [209]. Messenger RNA transcripts with the ACG sequence begin translation of three codons downstream, expressing a VDR protein that is three amino acids shorter (424 aa vs 427 aa). Functional differences among FokI genotypes show that VDR protein coded by the F-VDR (short, 424 aa) allele interacted more efficiently with Transcription Factor II B (TFIIB) and showed greater transcriptional activity than the full-length VDR protein coded by f-VDR allele [210]. The impact of FokI polymorphism on 1,25(OH)2D3-VDR mediated immune modulations has been recently evaluated [211]. The shorter F-VDR form was linked to a higher transcriptional activity driven by NF-Kβ and NFAT motif as well as IL-12p40 promoter-driven transcription.
Consequently, it was also shown that monocytes and DCs of homozygous subjects for short F/F VDR genotype express higher IL-12 mRNA and protein than cells with a long f/f VDR genotype, concluding that the short F-VDR allele is associated with a more active immune response.
Finally, three restriction fragment length polymorphisms (RFLP), designed as BsmI (rs1544410), ApaI (rs7975232) and TaqI (rs731236), and a PolyA (rs 17878969) microsatellite polymorphism, have been described in the 3'-UTR region of the VDR gene, showing strong linkage disequilibrium in Caucasian population [212,213].
Functional effects of 3,-UTR variants have been related to alterations in VDR mRNA stability [208]. Functional effects of 3,UTR haplotypes on VDR mediated immune modulation have been evaluated. Haplotype b-a-T (for BsmI and ApaI RFLP, cut allele, and TaqI RFLP, uncut allele) was associated with decreased IL-12p40 and IFN-γ and increased IL-10 cytokine response of 1,25(OH)2D3 treated peripheral blood mononuclear cells [214].
This seems to indicate that 1,25(OH)2D3-VDR mediated Th1 polarization is haplotype dependent with the b- a-T haplotype producing lower Th1 polarization effect.
The 3 ’ UTR polymorphisms have been associated with an increased susceptibility to infection by bacteria and viruses, including Mycobacterium tuberculosis [215,216], Mycobacterium leprae [217], Dengue Virus [218], HTLV-1 [219], HBV [220] and Respiratory Syncytial Virus [221].
1,25(OH)2D3-mediated signals appear to have a protective role in tuberculosis, in vitro [15] as well as in vivo [216, 222]. However, in vivo 1,25(OH)2D3 treatment exhibits a negative influence on the clinical course of both toxoplasmosis [223,224] and leishmaniosis [16] in a mouse.
On the contrary, a clinical trial evaluating the effect of vitamin D as a supplementary treatment for tuberculosis showed no overall effect on human beings [225].
In the context of HIV-1 infection, two VDR polymorphisms (BsmI and FokI) have been associated with susceptibility to faster progression towards AIDS-defining illness on a Spanish cohort of HIV-1 positive intravenous drug users. BsmI B/B homozygotes [22] and FokI F/f heterozygotes [21] were over represented among those reaching AIDS outcome. Both genotypes are associated with functional effects, enhanced VDR activity (FokI-F allele) and increased mRNA stability (BsmI-B allele).
Apart from the association of VDR gene variants with progression to AIDS, these gene variants could also influence the susceptibility to HIV-1 infection. Studies comparing Spanish HIV-1 positive intravenous drug users vs. HIV-1 exposed uninfected controls [20] and Indians HIV-1 positive vs. healthy controls [19] have suggested that the promoter Cdx-A1012G (G-A) haplotype and the 3’ UTR haplotypes containing the BsmI-b allele are associated with protection to HIV-1 infection. Since protective haplotypes confer a lower efficiency on the vitamin D pathway, it was inferred that hampering vitamin D signalling could confer protection to HIV-1 transmission.
More on the topic CONTRIBUTION OF VITAMIN D ON PROTECTION/RISK TO HIV INFECTION AND DISEASE PROGRESSION TO AIDS:
- Corynebacterium bovis Infection: Coryneform Hyperkeratosis; Scaly Skin Disease
- Clostridium piliforme Infection: Tyzzer’s Disease
- Different Responses
- Polarization and Collapse