HIV-1 LATENCY AND PERSISTENCE
Interest in retroviral latency or, more generally, persistence preceded the acquired immunodeficiency syndrome (AIDS) epidemic. Although retroviral latency was initially studied in avian retroviruses, the understanding of these processes in vivo has significantly increased in recent years, utilizing HIV-1 as a model.1 It is critical, at the outset, to delineate the concepts of cellular latency and persistent replication for retroviruses in vivo.
Viral latency is a general property of many viruses, including herpes viruses. Retroviral latency, however, can be defined as integrated provirus with no active transcription. Low-level chronic yet productive viral expression may best be characterized as persistent or cryptic replication, as HAART seems to not fully ablate all replication from previously infected cells. This can take place after initially successful viral suppressive therapy or antiviral immune responses in the infected host. Such cryptic viral replication may also continue in immunologically privileged sites, such as the brain and testes. As well, transcriptionally active but nonproductive viral infections (i.e., certain viral mRNA are expressed but not intact virions) may also occur (see below).The HIV-1 replicative cycle contains many possible stages for latency, both pre- and postintegration into the human cell genome.1 It involves binding to cellular receptors and co-receptors (i.e., CD4 receptor and chemokine co-receptors, CCR5 and CXCR4), with subsequent viral core internalization and reverse transcription of the viral RNA template into a double-stranded DNA intermediate. The viral DNA is then integrated into the host cell genome as proviral DNA. Transcriptional activation of the integrated provirus is controlled by a complex series of interactions between HIV-1 regulatory proteins (e.g., Tat and Rev) and cellular transcription factors, controlled by the state of activation of the host cell.
The activated provirus produces various viral messenger RNA species and, in some cases, new virions (reviewed in O’Brien and Pomerantz1).Many studies demonstrated that “clinical” HIV-1 latency, when defined as no viral expression in an entire untreated infected individual, does not exist at the organismal level in any stage of disease.16-18 In the great majority of HIV-1-infected individuals, some cultivable virus may be
HIV-1 Interactions with Target Cells InVivo
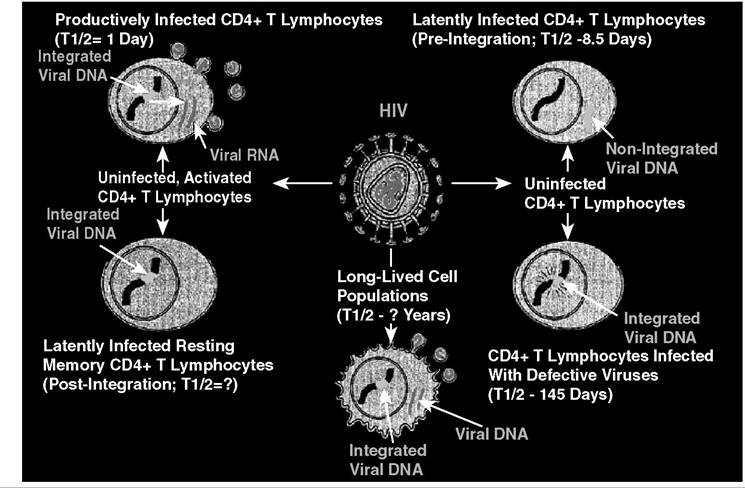
FIGURE 26.1 The in vivo dynamics and cellular reservoirs for HIV-1. This figure illustrates the different cell-types infected with HIV-1 in vivo and the possible sites for long-lived infected cell populations and latently infected cells. Red viral genomic RNA; Green, integrated or unintegrated viral DNA; Blue, host cell chromosome. Upper left, productively infected cell; Upper right, pre-integration latently infected; Lower left, integrated latently infected; Lower middle, long-lived population; Lower right, cells with defective proviruses.
recovered at all stages of disease.16-18 Nevertheless, data demonstrate that in the HIV-1-infected individual, some cells contain proviral DNA but express little or no viral RNA and produce few or no virions.16,19,20 In addition to postintegration latency, various states of preintegration HIV-1 latency were described both in vitro and in vivo.21-25 As such, latency at a cellular level exists in vivo, and the number of latently infected cells may vary based on the stage of disease. As HIV-1 infects, in vivo, CD4+ T cells, monocyte/macrophages, and other nonimmune-based cells,14,26,27 the virus may maintain cellular latency by different mechanisms in differing cell types (see Figure 26.1). This possibly represents important research opportunities in further elucidating mechanisms of HIV-1 latency in vivo.