Precision medicine-approaches to targeted therapy
Understanding tumour biology, identifying mutations and genomic abnormalities, and elucidating their role in cancer pathogenesis have important implications in the optimal clinical management of patients.
Disease biology influences tumour behaviour and this information can potentially be harnessed to plan therapy in a number of ways:1. Potential roles as prognostic or predictive factors. A predictive role is best exemplified when a genomic or proteomic alteration represents a biomarker of response or resistance to a specific target therapy.
The advent of genomic medicine has laid the foundation for predictive and prognostic markers. As a result, there is increasing use of these markers within the treatment planning of women with gynaecological cancers. Prognostic markers are clinical or biological characteristics that assist in estimating the likely outcome of untreated patients as well as the likelihood of disease recurrence. Predictive biomarkers are biological elements that can help predict likelihood of response to a specific treatment or describe how it contributes to increased risks of developing disease. Again, in gynaecological cancers these include mutations in DNA repair genes or mutations in the homologous recombination repair pathway; time to progression from the last chemotherapy; or HPV infection.
2. Identify novel potential therapeutic target (‘druggable’ mutations).
3. Identify individuals or family members at high risk to develop cancer and activation of specific screening and risk reduction strategies.
Molecular characteristics of gynaecological malignancies
Tumour-specific targets in gynaecological malignancies
The Cancer Genome Atlas Research Network (TCGA) has been responsible for important genomic data that have accelerated molecularly driven research in high-grade serous ovarian and endometrial cancers.
Findings from the TCGA in HGSOC illustrate the catalogue of mutations detected such as the universal mutation of TP53 in all tumours (96%); homologous recombination deficiency in almost half of tumours analysed; as well as the low prevalence but statistically recurrent somatic mutations in NF1, BRCA1, BRCA2, RB1, and CDK12; 113 significant focal DNA copy number aberrations; and promoter methylation events involving 168 genes (45) (Figure 67.3). TCGA findings and results published from subsequent studies clearly demonstrate that HGSOC is inherently genomically unstable and not characterized by a solitary driver mutation that is druggable. Furthermore, theTCGA has also been responsible for a similar in-depth examination of endometrial cancers identifying frequent TP53 mutations in high-grade endometrioid tumours and frequent mutations in PTEN, CTNNB1, PIK3CA, ARID1A, KRAS, and ARID5B DNA- binding protein (47) in most endometrioid tumours (Figure 67.3). The data also provide a strong rationale for a reclassification of these cancers into four distinct subtype—POLE ultramutated, microsatellite instability hypermutated, copy-number low and high—that may have clinical impacts at the time of postsurgical adjuvant treatment (47). Indeed, patients diagnosed with POLE ultramutated tumour have generally very good outcome. Additional investigations are needed to interpret the functionality of these observed genomic alterations and whether this can translate into therapeutic targets that could improve treatment outcomes (Figure 67.4).
Contemporary investigations into the molecular landscape of clear cell cancer and low-grade serous (LGSOC) ovarian cancers have also been published. Clear cell ovarian carcinomas harbour defects in PTEN(approximately 10% of cases), PIK3CA (approximately 50% of cases), roughly 50% of cases harbour ARID1A mutations, a minority harbour PPP2R1A mutations (48), and lastly, roughly 33% exhibit amplification of chr20q13.2 and do not typically harbour Wnt-activating CTNNB1 mutations (49).
Comprising 5-10% of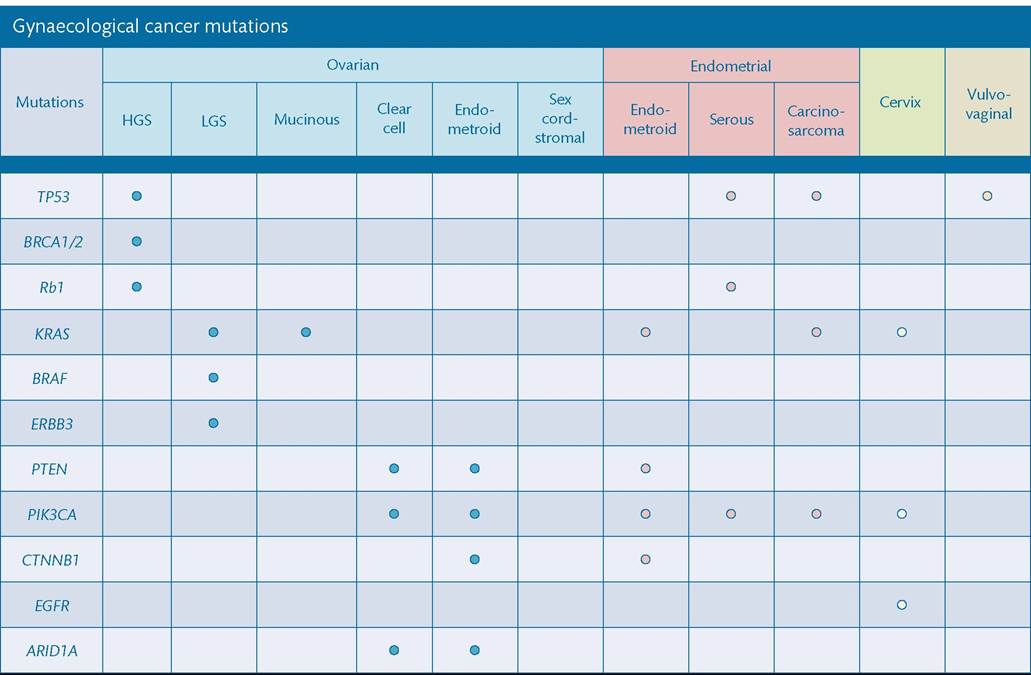
Figure 67.3 Common mutations according to gynaecological cancer subtypes. HGS, high-grade serous; LGS, low-grade serous.
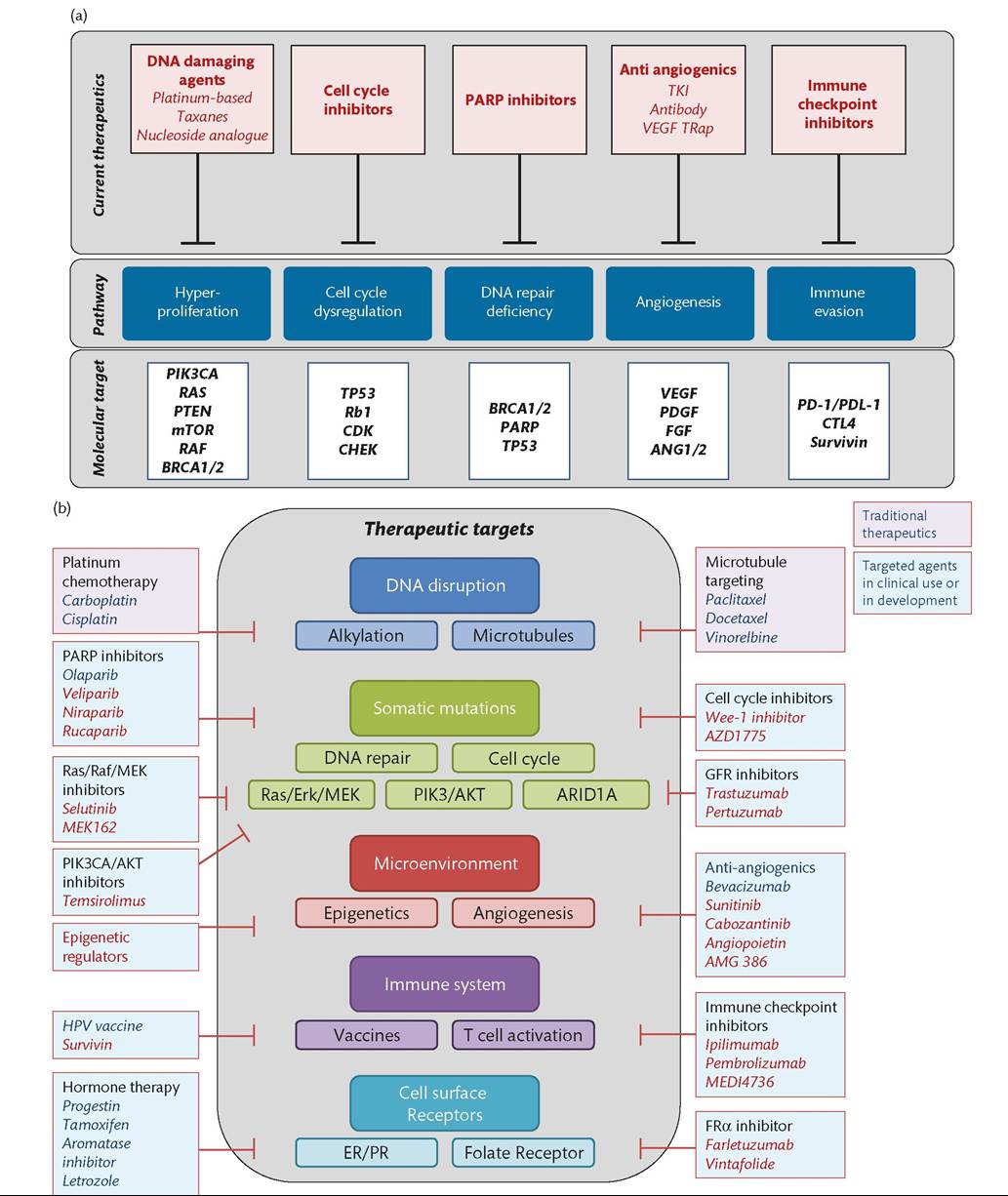
Figure 67.4 (a) Select therapeutic classes, pathways, and molecular targets in gynaecological cancers. (b) Therapeutic targets.
serous ovarian cancers, LGSOC tumours have documented prevalence of KRAS mutations (35-57% of cases) as well as mutations in BRAF during early-stage disease but rare in the advanced setting (50, 51).
Pathogenic HPV infection is the most important and potentially preventable causative factor in cervical cancer and responsible for more than 95% of cervical cancers. The molecular landscape of the disease was recently updated (52) and study findings show several mutations such as recurrent E322K substitutions in the MAPK1 gene (8%), inactivating mutations in the HLA-B gene (9%), and mutations in EP300 (16%), FBXW7 (15%), NFE2L2 (4%), TP53 (5%), and ERBB2 (6%) genes in primary squamous cell carcinomas, as well as somatic ELF3 (13%) and CBFB (8%) gene mutations in 24 adenocarcinomas. In cervical cancer, mutations in TP53, PI3K pathway, and KRAS are found with some different patterns between squamous carcinoma and adenocarcinoma. Furthermore, study gene expression data suggest that HPV integration into the host cell genome is a common mechanism for overexpression of target genes, and potential drivers of carcinogenesis (52).
Spatial and temporal heterogeneity
Therapeutic resistance remains a clinical and research challenge which has to be overcome to improve effectiveness of precision targeted therapies. There is significant genetic diversity within and between common tumours (53) which builds on recognition of morphological heterogeneity within tumours (54). This spatial and temporal heterogeneity may arise as a result of either random genetic drift or as a result of phenotypic advantage selected from a particular environment (55, 56).
Ovarian cancers exhibit branched patterns of evolution, whereby several subclones grow out driving disease progression and manifest as intratumoural heterogeneity (54). Treatment may also induce selection pressure for intratumoural driver mutations or between tumours. This means that tumours develop new patterns of resistance and emphasize the need for repeat assessment of patient samples throughout disease to accurately profile and understand tumour genomic architecture (54). Heterogeneity between primary and metastatic lesions impacts the efficacy of subsequent therapy.Spatial heterogeneity within HGSOC has been elegantly shown by several groups (57, 58). The degree of genomic diversity is evident within primary untreated tumours exhibited by extensive intratumoural variations in mutation, copy number, and gene expression profiles, with key driver alterations in genes present in only a subset of samples (e.g. PI3KCA, CTNNB1, NF1) (57). TP53 mutation is the most prevalent abnormality in HGSOC and essentially defines the disease, and is the only somatic mutation consistently present in all samples (57). Taken together, spatial and temporal heterogeneity in gynaecological cancers must be considered during treatment selection and throughout disease. Clinically relevant biomarkers may differ between primary and metastatic disease as has been shown in other cancers (59, 60).
Inherent and acquired resistance
Inherent and acquired forms of resistance impact treatment course whether in the upfront or advanced setting. Despite the lack of classical driver mutations in gynaecological cancers, frequently mutated genes exist within each subtype. The mutational landscape of gynaecological cancers is varied with several driver-like mutations identified; the current most widely cited being TP53 in high-grade serous cancers, mutations in DNA repair genes BRCA1/2, as well as homologous recombination repair pathway genes (45). Disease recurrence in a previously radiated area, commonly used in the treatment of gynaecological cancers, may also have acquired specific mechanisms of drug resistance.
Germline mutations
Mutations in germ cells are referred to as germline since these mutations occur in cells responsible for creating gametes and as such, mutations that can be passed on to offspring (61). With respect to gynaecological malignancies, well-documented mutations in heritable genes are those in BRCA1/2 and loss of DNA mismatch repair in Lynch syndrome. Mutations in these genes have also been suggested to contribute to several other malignancies such as prostate, pancreatic, and stomach cancers (62). Germline mutations in the DNA repair pathway beyond BRCA1/2 are under investigation, such as RAD51 or PABL2.
Key aberrations in gynaecological cancers
Ovarian cancer
EOC encompasses five distinct diseases, each with individual patterns of epidemiological and genetic risk factors, precursor lesions, patterns of spread, and molecular events during oncogenesis, response to chemotherapy, and prognosis (63). Low-grade serous ovarian tumours (LGSOC) commonly arise from precursor lesions and clear cell potentially from endometriosis. Conversely, TP53 mutations are ubiquitous in HGSOC; whereas mutations in BRCA1/2 and the homologous recombination repair pathway genes are key drivers in carcinogenesis (45). Beyond BRCA1/2 mutations in HGSOC, there are no validated predictive genetic biomarkers, though work is ongoing to evaluate the impact on outcome based on patterns of homologous recombination deficiency, loss of heterozygosity, and somatic mutations. The five different EOC subtypes are defined by particular molecular profiles that correlate with outcome:
• HGSOC is a disease characterized by genomic instability and ubiquitous TP53 mutations, frequent deficiency in homologous recombination pathway, and high prevalence of copy number alterations with few recurrent mutations.
• LGSOC harbours a KRAS mutation in 20-40% while only 5% have a BRAF mutation. This implicates the MAPK signalling pathway as a key node in LGSOC—in fact, 80% of LGSOC have activation of the MAPK pathway (64).
Although, at this stage, there remains insufficient data to use this mutation as a predictive biomarker for a treatment decision.• Mucinous ovarian cancer is a rare entity characterized by resistant disease and low response rate to platinum/p aclitaxel chemotherapy. Based on the analogy with gastrointestinal cancer, oxaliplatin and 5-FU chemotherapy has been used with modest activity. Approximately 60% of cases show a KRAS mutation and the presence of a KRAS mutation predicts resistance to anti-EGFR treatment, as already demonstrated in gastrointestinal cancer (65). HER2 is amplified in about 15-20% of cases and anti-HER2 agents may be active in this subgroup of patients but no data from clinical trials are available.
• Clear cell ovarian cancer is also associated with a low response to platinum-based chemotherapy. Molecular profiling of this disease is characterized by the lack of BRCA1/2 mutations as well as a high frequency of PI3K∕AKT∕mTOR pathway alterations (40-50%) (66). Interestingly, it has been documented that roughly 50% of clear cell ovarian cancer cases harbour mutations in the ARIDlA gene (48).
• Endometrioid EOC is characterized by mutations in the beta- catenin and PTEN genes, as well as the tumour suppressor genes PIK3CA and ARIDlA. Functional loss of PTEN through mutations in the PI3K∕AKT∕mTOR pathways has been the subject of considerable therapeutic research, with some promising results (67). Endometrioid ovarian cancer has histological and molecular aspects similar to endometrial adenocarcinoma.
Endometrial cancer
Findings from TCGA have helped illustrate the genomic landscape of this disease. In total, the collaborative analysed 373 endometrial carcinomas using array- and sequencing-based technologies. Data show uterine serous tumours and approximately 25% of high-grade endometrioid tumours harbour extensive copy number alterations, few DNA methylation changes, low ER/PR levels, and frequent TP53 mutations (47). Most endometrioid tumours had few copy number alterations or TP53 mutations; and findings also highlight frequent mutations in PTEN, CTNNBl, PIK3CA, ARIDlA, and KRAS, as well as novel mutations in the SWI/SNF chromatin remodelling complex gene ARID5B. Interestingly, a subset of endometrioid tumours analysed had a markedly increased transversion mutation frequency and newly identified hotspot mutations in POLE. As a result, a novel classification system which has prognostic value has been devised to include four categories: POLE ultramutated, microsatellite instability hypermutated, copy-number low, and copy-number high (47). Ongoing studies are evaluating the predictive value of this classification with targeted and immunological therapies.
Cervical cancer
While HPV infection contributes significantly to the development of cervical cancer, several genes have been documented to play a role in the development of disease. Involvement of TP53 gene mutations are documented related to HPV molecular transformation—HPVE6 gene-stimulated degradation interferes with p53 function (68). The HPV oncoproteins E5, E6, and E7 are the primary viral factors responsible for initiation and progression of cervical cancer. E6, E7, and to a lesser extent E5 play key roles in upregulating angiogenesis through the VEGF pathway through their effects on p53 degradation, hypoxia- inducible factor-1α (HIF- 1α), and inactivation of retinoblastoma protein (pRb). PIK3CA, STK11, KRAS, EGFR, NOL7, CDKN2A, PTEN, and binding protein p300 (EP300) genes have all been documented in the Cervical Cancer Gene Database, as well as the National Human Genome Research Institute, the National Cancer Institute, the COSMIC Database and TGCA (68-70).
Other cancers-vaginal and vulval cancers
The aetiology of vulval squamous cell carcinoma (VSCC) includes high-risk HPV-dependent infection manifesting with usual vulval intraepithelial neoplasia (VIN) as a precursor lesion, and an HPV-independent route associated with differentiated VIN, lichen sclerosus (a chronic dermatosis associated with autoimmune diseases), and genetic alterations such as TP53 mutations (71). As reported recently by Trietsch et al. (2015), a review of current literature confirms the hypothesis that HPV and TP53 mutations play almost separate, but key roles in the carcinogenesis of VSCC. Study findings go on to suggest that in the small number of articles with survival data available, tumours harbouring a mutation, which are most often HPV-independent VSCC, have a worse prognosis than VSCC without (epi)genetic changes (71). Other mutations described in VSCC and its precursor lesions include PTEN and CDKN2A gene mutations (72, 73). In a study of 107 formalin-fixed, paraffin- embedded primary surgically treated tissues from patients with VSCC, Sanger sequencing and mass spectrometry revealed somatic mutations in 62% of samples (74). In addition to HPV infection and TP53 mutations, CDKN2A(p16), HRAS, and PIK3CA mutations were frequently seen in HPV-negative patients (74). Interestingly, patients with somatic mutations, especially HRAS, have significantly worse prognosis than those patients lacking these changes (74).
Potential druggable mutations
TP53 mutations
The tumour suppressor gene TP53 (MIM#191117) is responsible for the production of p53 protein, which is charged with regulating cell division and death; and thus regarded as the guardian of the genome (75). During times of cellular stress such as DNA damage, hypoxia, oncogene activation, or∕and nutrient deprivation, p53 functions as a transcription factor that is activated and leads to cell cycle arrest, apoptosis, and metabolic adaptation (76). While non-heritable or somatic mutations in p53 are one of the most frequent alterations in cancer (77), germline mutations in TP53 have been documented (77). Germline mutations in TP53 are responsible for the rare Li- Fraumeni syndrome, a familial clustering of early-onset tumours including sarcomas, breast, brain, and adrenal cortical carcinomas (78, 79). Direct and indirect approaches are under investigation to target p53 alterations in gynaecological cancers.
BRCA1/2 mutations
With respect to gynaecological cancers, heritable or germline mutations in BRCA1 or -2 increase the lifetime risk of developing breast and ovarian cancers. Published findings show that while the lifetime risk of developing ovarian cancer is low in the general population, risk increases to between 40% and 60% for BRCA1 and 11-27% for BRCA2 germline mutation carriers, respectively (80). In all ovarian cancer, these particular mutations are associated with 10-15% of cases and in HGSOC, almost 20% of cases (81). Importantly, this includes women with HGSOC who do not have a family history of either breast or ovarian cancer. As such, NCCN guidelines suggest germline testing for BRCA1/2 germline mutation is made available for all HGSOC diagnoses.
Over the last several years, there are increasing data showing the benefits of targeting pathways necessary for maintaining∕ safeguarding DNA integrity, including BRCA1 and BRCA2 (82). Clinically, germline BRCA1/2 mutation has been shown to be predictive of sensitivity to platinum (83) as well as to agents targeting impairments in DNA repair pathways—typically conferred by BRCA mutations—as evidenced by recent approval of PARP inhibitors, such as olaparib, responsible for blocking a major DNA repair enzyme. Mechanistically, these agents operate via the parallel endorsement of DNA double-stranded breaks and interference of double-stranded break repair by PARP protein inhibition (84, 85) that collectively results in cell death. Germline and tumour testing for BRCA1/2 mutations gives important information to clinicians regarding the potential predictive benefit of PARP inhibitors in women with ovarian cancer and has led to the subsequent approval of these agents. Together, this is a successful example of targeted therapy development that showcases how understanding germline and somatic mutations, and their function, enables the strategic targeting of these vulnerabilities and leads to effective therapeutic agents.
Homologous recombination deficiency
There are numerous genes involved in homologous recombination repair pathways that also contribute to disease including ATM, CHEK2, BARD1, BRIP1, Mre11, RAD50, NBS1, RAD51C, RAD51D, and PALB2 (86). Other alterations affecting homologous recombination repair include amplification of EMSY (8%), deletion/mutation of PTEN (7%), hypermethylation of RAD51C (3%), mutation of ATM or ATR (2%), or mutation of other homologous recombination genes (5%). These are suggested to have the phenotype of ‘BRCAness’ in serous cancers and are predicted to behave like BRCA-deficient tumours despite normal germline BRCA1 and BRCA2 genes (86, 87). These patients may also benefit from PARP inhibitors.
An autosomal dominant, inherited cancer-s usceptibility syndrome in women with endometrial cancer, Lynch syndrome (LS) has also been found to harbour defects in DNA repair in MLH1, mutS homolog 2 (MSH2), MSH6, PMS1 homolog 2 (PMS2) (a mutations of DNA mismatch repair system component) genes, and others (88). Similar to HGSOC, BRCA1/2 germline history is insufficient to identify affected women and as such, changes in disease screening are gaining momentum. In addition to clinical criteria, immunohisto chemistry is universally used to screen for impairments in mismatch repair genes with a high sensitivity, which is followed by subsequent mutation testing for microsatellite instability (89). It has been hypothesized that these patients are more likely to respond to immunotherapy and prospective trials are ongoing to assess this question.
Targeting somatic mutations
DNA repair
BRCA1/2 and homologous repair deficiency
PARP inhibitors are a new class of agents developed and refined for clinical use over the last several years. Cancer cells with homologous repair deficiency are not able to repair double-stranded DNA breaks and if PARP is inhibited, the single-stranded breaks also cannot be repaired, which leads to cell death. This mechanism or concept is known as synthetic lethality. Olaparib, the first developed PARP inhibitor, has been approved in the United States as monotherapy for BRCA1/2-mutated patients that have received three or more lines of the treatment and as maintenance therapy in patient with relapse platinum-s ensitive HGSOC and more recently after the first line therapy for women diagnosed with BRCA1/2 mutation HGSOC. Several other PARP inhibitors have been approved since.
PARP inhibitors have shown activity in different trials as monotherapy or as maintenance, in BRCA mutated women but also in BRCA wild-type patients, leading to the definition of a special subgroup of HGSOC patients with a ‘BRCAness’ phenotype similar to BRCA-mutated patients (higher platinum sensitivity and benefit from PARP inhibitor treatment). Other trials are ongoing with different PARP inhibitors such as rucaparib, niraparib, and veliparib to address the benefit of this targeted therapy beyond BRCA1/2 mutation.
CDK12 (a member of cyclin-kinase protein family) is mutated in 3% of HGSOC and its activity is important for homologous recombination. When CDK12 is not active, depletion in HR enzymes occurs and reduction in BRCA expression has been observed. Cells harbouring a mutation in the CDK12 gene could be more sensitive to cisplatin and PARP-inhibitor, representing an interesting new therapeutic option.
The cell cycle
DNA repair pathways have cell cycle specificity, especially within the G2/M checkpoint (90). Agents targeting CHK1/2 and WEE-1 kinases have minimal single-agent activity in the absence of p53, or more likely, p53 and second (so far undefined) mutations. Efficacy is much greater when examined in a defined p53 mutant background in which the G1/S checkpoint is aberrant and also in combination. There is a subset of patients with ovarian cancer with wild-type BRCA1/2 function and cyclin E amplification and overexpression.
TP53
Somatic mutations in TP53 are the most documented mutations contributing to cancer and a defining characteristic of HGSOC (45) as well as in endometrial carcinomas (47). The majority of TP53 mutations are missense and cause single amino acid changes at many different positions and as such, vary in their structural impact (77). These typically result in high levels of dysfunctional p53 protein (91) and efforts remain underway to devise novel therapies targeting these proteins. Alternatively, in cervical cancers that have low mutation rates, p53 is inactivated by an alternate mechanism; specifically, it is targeted for degradation by HPV E6 (77, 92). Future studies are warranted to determine the precise manner in which p53 proteins or TP53 somatic mutations can be leveraged to devise novel therapies in gynaecological cancers.
KRAS/BRAF/MAPK pathway
Between 40% and 60% of LGSOCs have a RAS mutation (93). RAS is a family of different oncogenes and the most relevant are KRAS, NRAS, and HRAS and point mutations are frequently detected in codons 12, 13, and 61. Activating mutations in KRAS, NRAS, or BRAF promote tumourigenesis through a constitutive activation of the MAPK/ERK (mitogen-activated protein kinase/extracellular- signal-regulated kinase) pathway. MAPK inhibition with anti-MEK agents is being investigated. Further research is necessary to analyse the possible prognostic or predictive role of these mutations in order to select patients that may benefit from this treatment or define which tumours could be resistant to a specific drug.
PIK3CA/AKT/mTOR pathway
Aberrant activation of PI3K induces phosphorylation of AKT that subsequently activates mTOR. mTOR promotes cell proliferations and angiogenesis. Different mechanisms contribute to activate this pathway such as receptor tyrosine kinase activation or amplification, mutation, or silencing of negative regulators and activation or amplification of downstream kinase.
Approximately 70% of ovarian cancers are associated with activation of PI3K signalling, with a higher incidence in clear cell ovarian cancer and endometrial cancer. This activation may occur following mutation or amplification in one of the two subunits of PI3K (PIK3CA (12%) or PIK3R1 (3.8%)), AKT1 (2%), AKT2 (13.3%), or mTOR or through loss of function of PTEN that in normal cells acts as an inhibitor of the PI3K pathway (94). In HGSOC, activation of PI3K cascade is rare (<5%) but is more frequent in clear cell ovarian cancer and endometrioid ovarian cancer. Loss of PTEN is present in 40% of clear cell ovarian cancer.
Different clinical trials exploring the activity of PI3K, AKT and mTOR inhibitors are ongoing as a single agent or in combination with chemotherapy. Agents targeting this pathway have shown interesting activity in gynaecological cancers, starting with mTOR inhibitors; however, the ability to predict outcome based on somatic mutations in the pathway genes has been challenging, perhaps due to a redundancy of molecular pathways to allow cells to survive. As a consequence, combination studies are underway to understand and overcome resistance, such as combinations of PI3K inhibitors and RAS/RAF/MEK inhibitors, with potential synergy in blocking an overactive pathway but also limited by toxicity.
ARIDlA
ARIDlA encodes the protein BAF250α, a core subunit of SWI/SNF that acts by inhibiting cancer cell development. Mutations of this gene have been found in 46-57% of clear cell ovarian cancer, 30% of endometrioid ovarian cancer, and 40% of uterine endometrioid carcinoma and loss of ARIDlA expression is common in endometriosis (48, 95). Currently a specific treatment targeting ARIDlA has not been identified, as usually happens with loss-of-function mutations; however, a study by Bitler et al. has shown that inducing synthetic lethality by way of inhibiting EZF2 (enhancer of zeste homology 2) in ARIDlA-mutated patients results in clear cell ovarian cancer growth inhibition (96).
Targeting the microenvironment
Angiogenesis inhibition
Angiogenesis is required for cancer cell proliferation. Various mediators of angiogenesis have been reported, including VEGF. Hypoxia and the mechanisms that mediate hypoxic response are key drivers of physiological angiogenesis. Under hypoxic conditions expression of HIF-1α is induced in endothelial cells, resulting in VEGF-A, and vascular endothelial growth factor receptor 2 (VEGFR-2) expression. Targeted therapy against angiogenesis using bevacizumab, a humanized VEGF-neutralizing monoclonal antibody, improved the median progression-free survival rate in advanced ovarian cancer and the overall survival in cervical cancer (97). These two indications have been approved in gynaecological cancers. Despite several positive phase II trials with progression-free survival improvement, no antiangiogenetic has been approved in endometrial cancer. A crucial first step is to determine potential markers of response.
Epigenetic modulation
Epigenetics is a field that focuses on the study of heritable changes which do not result from changes in the DNA sequence, including DNA methylation. These are divided into histone acetylation, deacetylation, and methylation events carried out by histone acetyltransferases, histone deacetylases, and histone methyltransferases, respectively. Aberrant methylation of CpG islands which are found in close proximity to gene transcription initiation sites and are normally methylated has been linked to tumour initiation and progression. Because of their involvement in tumourigenesis, the biology of these changes is now being investigated so that its potential in the treatment of ovarian cancer is fully understood.
Immunotherapy
The ability of cancer cells to evade immune destruction has become recognized as one of the hallmarks of cancer. This has paved the way for the development of novel therapeutic agents that can enhance activation of antitumour immune responses or reverse immunosuppressive mechanisms through which tumours escape immune- mediated rejection (98).
Vaccines
The most notable example of this approach is vaccination for HPV which is now approved for the prevention of cervical cancer.
The identification of unique differentiation proteins expressed in gynaecological cancer has led to the exploration of various vaccination approaches, including simple vaccine preparations consisting of specific peptides and proteins, as well as more complex strategies, such as engineered cellular vaccines, dendritic cell vaccines, virus- vectored vaccines, and oncolytic viruses. The majority of studies have explored the cancer-testis antigens (e.g. NY-ESO-1) and proteins known to be overexpressed in EOC (e.g. p53, survivin, and MUC1). Although many studies have demonstrated induction of an immune response to the vaccines, very few have demonstrated clinical benefit. It is likely that these strategies are insufficient to overcome immune tolerance to self-antigens and to result in efficient activation of antigen-specific T cells, although they may prove to be valuable in combination with other therapies or be used in a different treatment schedule.
Therapies to enhance T-cell activation
The survival, proliferation, and activation of T cells are controlled by a variety of factors, including cytokines and a range of immunostimulatory and inhibitory receptors. Identification of the co-stimulatory and co-inhibitory receptors that regulate T-cell activation has led to the development of antibodies that target these receptors. Targeting such receptors, an approach termed ‘immune checkpoint blockade', has demonstrated activity in preclinical cancer models and in clinical trials. In particular, antibodies targeting the inhibitory receptors cytotoxic T-lymphocyte-associated antigen 4 (CTLA-4) and programmed death 1 (PD-1), as well as the PD-1 ligand (PD-L1), are the agents of this type that are most advanced in clinical development. Given the potential interest in gynaecological cancers, these strategies are being actively assessed. Indeed, tumour infiltrating lymphocyte (TIL) counts and peritumoural lymphocytes have been described as independent predictors for microsatellite instability high-status group in endometrial cancers (99). Moreover, the presence of TILs appears to be an independent prognostic factor (100). TIL infiltration and the presence of CD8 at diagnosis of ovarian cancer have been also described to be predictive of clinical outcomes in ovarian cancers.
Another approach under investigation is the adoptive cell therapies which rely on the infusion of large numbers of autologous tumour-reactive T cells that have been isolated from tumours and expanded in vitro.
Targeting cell surface receptors
Hormones
Hormonal therapy remains the sole approved targeted therapy in endometrial cancer. A significant proportion of endometrial cancer—in particular, type I tumours—express ER or PR, described to be predictors of favourable survival and rendering hormonal therapy an attractive therapeutic strategy. Current investigations are ongoing to understand mechanisms of resistance to hormonal therapies and the potential interest of combination therapies.
Folate
Several publications report that folate receptor alpha (FRα) is an attractive candidate for targeted biological therapy of ovarian cancer and potentially endometrial cancers (101-103). Indeed, approximately 90% of EOC express FRα and its expression correlates with stage and grade of malignancies (104). In contrast, FRα has a low expression on the apical surface of most normal cells (105). This difference in expression leads to high tumour-to-normal ratios and makes FRα a very attractive target for therapeutic and imaging purpose (106).
Combination approaches
Incorporating a novel targeted or immunological therapy into a therapeutic algorithm requires careful thought related to the current evidence-based standard of care. Therefore, early-phase studies are investigating strategies to combine as well as sequence novel agents in conjunction with chemotherapy, radiation, or other targeted agents. These early studies need to integrate strong translational studies to understand mechanisms of response and resistance that will guide future drug development to optimize treatment for our patients with a balance between drug activity and toxicity. Early examples of the efficacy of this approach have emerged from combinations incorporating antiangiogenics such as bevacizumab and cediranib as well as subsequent sequential use as a single agent.
The ability to develop combinations is inherently related to the potential for synergy as well toxicity—PARP inhibitors have been more challenging to combine with chemotherapy because of overlapping haematological toxicity, but proven to be synergistic in sequence (21, 107). Many studies are currently evaluating evidence-informed combinations which will be evaluated in prospective trials.