AVIAN INFLUENZA
Leslie a. Reperant1, albert d.m.e. Osterhaus2 AND THIJS KUIKEN2
1Department of Virology, Erasmus MC, Rotterdam, The Netherlands
2Department of Virology, Erasmus MC, and Artemis Research Institute for Wildlife Health in Europe, Rotterdam, The Netherlands
AETIOLOGY
Avian influenza A viruses can be classified based on their pathogenicity in poultry into low pathogenic and highly pathogenic forms.
Nearly all isolates from wild waterbirds are low pathogenic avian influenza viruses (LPAIV). All known subtypes of LPAIV have been isolated from wild waterbirds, which are considered their natural reservoirs. All influenza A virus lineages occurring in other species, including established lineages of influenza A viruses in humans and other mammals, are thought to originate from avian influenza viruses of wild birds. LPAIV may be transmitted to poultry, where infection is subclinical or causes mild clinical disease. However, LPAIV of the H5 and H7 subtypes can evolve by selection in poultry into highly pathogenic avian influenza viruses (HPAIV) and cause systemic infection and severe, often fatal, disease in these species. HPAIV are rarely found in wild birds, and, if they are, typically they do not cause clinical disease. Only HPAIV H5N1 has caused disease in wild birds in Europe, sometimes in association with outbreaks with high mortality.Low pathogenic avian influenza has no synonyms. Highly pathogenic avian influenza is also called fowl plague or fowl pest.
EPIDEMIOLOGY
GEOGRAPHICAL DISTRIBUTION
LPAIV are widely distributed across Europe. Partly as a result of up- scaling of avian influenza surveillance as a result of the threat of HPAIV H5N1, LPAIV have now been isolated from wild birds in most European countries (Figure 2.1A). Some eastern and southern European countries have not yet reported LPAIV infection in wild birds, although recent studies have provided serological evidence of LPAIV infection.
Most LPAIV haemagglutinin (HA) and neuraminidase (NA) proteins described to date have been isolated from wild birds in Europe (Table 2.1) (4). Phylogenetic analysis of each of the eight genes of LPAIV isolated from wild
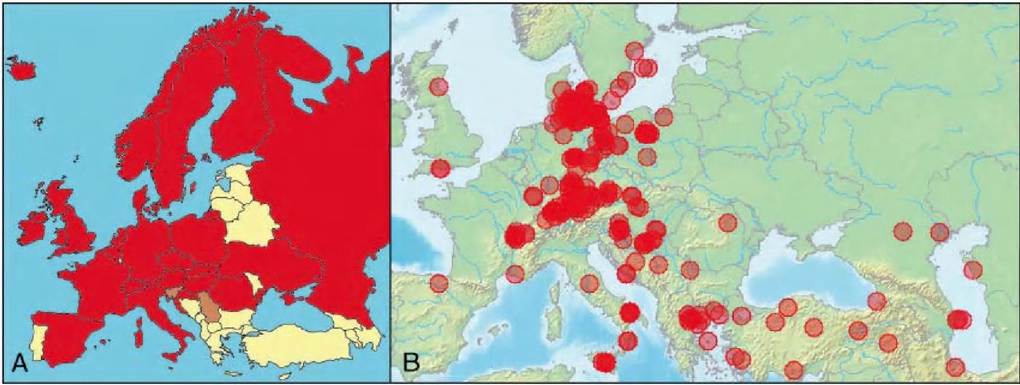
FIGURE 2.1 Geographical distribution of avian influenza viruses in wild birds in Europe. A. Geographical distribution of LPAIV. Red: countries where LPAIV have been isolated from wild birds and viral sequences deposited in a public database; brown: countries with serological evidence of LPAIV circulation in wild birds; yellow: countries with no published report of LPAIV infection in wild birds. B. Locations of wild birds found infected with highly pathogenic avian influenza virus H5N1 in Europe and the Near East between 2005 and 2009 (marked as transparent circles).
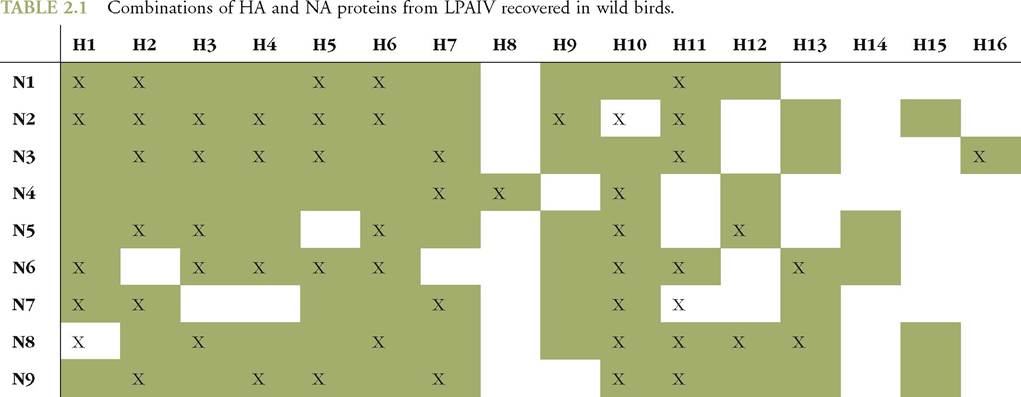
Green boxes: combinations of HA and NA proteins from LPAIV recovered in wild birds globally
X: combinations of HA and NA proteins from LPAIV recovered in wild birds in Europe
birds in Europe places them in a Eurasian clade. By contrast, LPAIV from wild birds in North America cluster in a separate, North American clade(5), and there may be a distinct phylogenetic clade for South American isolates1-6). Geographical separation of host species has probably shaped the gene pool of LPAIV into largely independently evolving lineages. Intercontinental dispersal of LPAIV of either clade has not been described to date, although there is growing evidence of reassortment of genes from LPAIV of different clades(7). For instance in Europe, genes of the North American clade have been recovered from LPAIV isolated in guillemots (Uria aalge) in the northern Baltic Sea(8). However, such intercontinental reassortment events remain rare. Therefore, despite shared migratory flyways used by LPAIV host species, e.g.
the East Atlantic flyway, LPAIVs mix infrequently between the Eurasian and North American clades(5).Since the autumn of 2005, HPAIV H5N1 originating from poultry in South-East Asia have infected wild bird populations across Eurasia (Figure 2.1B). HPAIV H5N1 has affected multiple wild bird species on several continents during the past 5 years. HPAIV H5N1 first reached Russia and south- eastern European countries in autumn
2005 and then spread to more western and northern countries during the following winter. Most outbreaks of fatal infection occurred during the winter and early spring of
2006 in multiple species of wild birds. HPAIV H5N1 reoccurred in wild birds in summer 2007, mostly in Germany, and were occasionally isolated in winter 2007— 2008 and winter 2008—2009, in the UK and central Europe.
HOST FACTORS
LPAIV have been recovered worldwide in at least 110 wild bird species from 26 families belonging to 13 orders. In Europe, LPAIV have been recovered in at least 35 wild bird species, from 10 families belonging to 8 orders (Table 2.2). Despite their wide host range, LPAIV typically infect bird species that live in wetland and aquatic habitats, in particular species of the orders Anseriformes (geese, ducks and swans) and Charadriiformes (waders and gulls), as well as Eurasian coots (Fulica atra) of the order Gruiformes in Europe (Figure 2.2A).
The host range of avian influenza viruses (AIV) is partly determined by the specificity of the HA protein for the receptor by which the virus attaches to the host cell. AIV typically bind to sialic acid moieties with a α2-3 linkage to galactose. These receptors are present on the surface of avian host cells, notably epithelial cells of the intestinal tract. However, the conformation of these receptors on intestinal epithelial cells of ducks, gulls and chickens differ(9). Such differences may contribute to the observed differences in susceptibility of different avian hosts to LPAIV infection.
Studies on prevalence of LPAIV infection in wild birds in Europe are based on isolation of LPAIV or detection of LPAIV nucleic acids in samples. The highest prevalence is
TABLE 2.2 List of wild bird species found infected with LPAIV in Europe. Prevalences are reported for species with more than 150 sampled birds.
| Order, family | Species common name | Species scientific name | Country | N | % |
| Anseriformes, | Greater white-fronted | Anser albifrons | The Netherlands, Sweden | 4325 | 2.2 |
| Anatidae | goose Greylag goose | Anser anser | Germany, the Netherlands, Sweden | 1432 | 1.5 |
| Pink-footed goose | Anser brachyrhynchus | The Netherlands, Sweden | 285 | 2.1 | |
| Bean goose | Anser fabalis | The Netherlands | 466 | 0.6 | |
| Barnacle goose | Branta leucopsis | The Netherlands | 1257 | 0.6 | |
| Brent goose | Brenta bernicla | The Netherlands, Sweden | 715 | 0.6 | |
| Northern pintail | Anas acuta | France, the Netherlands, Sweden | 920 | 3.3 | |
| Common teal | Anas crecca | France, Germany, Iceland, the Netherlands, | 2414 | 8.3 | |
| Eurasian wigeon | Anas penelope | Sweden France, the Netherlands, Sweden | 3596 | 2.5 | |
| Mallard | Anas platyrhynchos | Czechoslovakia, France, Germany, Hungary, | 15962 | 8.7 | |
| Garganey Northern shoveler | Anas querquedula Anas clypeata | Italy, Sweden, the Netherlands, Sweden France, Romania France, the Netherlands | 284 | 3.2 | |
| Common shelduck | Tadorna tadorna | France, Italy, Sweden | 1244 | 4.7 | |
| Tufted duck Long-tailed duck | Aythya fuligula Clangula hyemalis | The Netherlands Germany | 157 | 1.3 | |
| White-winged scoter Common scoter Common eider Bewick’s swan | Melanitta fusca Melanitta nigra Somateria mollissima Cygnus columbianus bewickii | Germany Norway The Netherlands, Sweden The Netherlands, Sweden | 153 | 2 | |
| Mute swan | Cygnus olor | Germany, the Netherlands | 1362 | 1.5 | |
| Charadriiformes, | Waders | Sweden, Italy | 3000 | 1.2 | |
| Caradriidae | |||||
| Charadriiformes, | Black-headed gull | Larus ridibundus | Germany, the Netherlands, Sweden | 2395 | 1.5 |
| Laridae | Common gull | Larus canus | The Netherlands | 226 | 0.9 |
| Herring gull | Larus argentatus | The Netherlands | 862 | 0.6 | |
| Greater black-backed gull Mediterranean gull Common tern | Larus marinus Larus melanocephalus Sterna hirundo | The Netherlands, Sweden France Germany | 875 | 1.5 | |
| Arctic tern Sandwich tern | Sterna paradisea Sterna sandvicensis | Germany Germany | 351 | 0.3 | |
| Ciconiiformes, | Grey heron | Ardea cinerea | Romania | ||
| Ardeidae | |||||
| Columbiformes, | Eurasian collared dove | Streptopelia decaocto | Hungary | ||
| Columbidae | |||||
| Galliformes, | Common pheasant | Phasianus colchicus | Hungary | 399 | 0.5 |
| Phasianidae | |||||
| Gaviiformes, Gavidae Gruiformes, Alcidae | Arctic loon Common murre | Gavia arctica Uria aalge | Romania Sweden | 843 | 0.7 |
| Gruiformes, Rallidae | Eurasian coot | Fulica atra | Germany, Hungary, Italy, the Netherlands | 2610 | 2.6 |
| Pelicaniformes, | Great cormorant | Phalacrocorax carbo | Germany, Romania | 4500 | 0.4 |
Phalacrocoracidae
typically recorded in dabbling ducks (Anas spp.), and notably in mallards (A.
platyrhynchos) and common teal (A. crecca) with a prevalence of 8% (Table 2.2). Interestingly, high prevalence was also reported in a small sample of estuary and marine ducks, such as shelduck (Tadorna spp.), scoters (Melanitta spp.) and eiders (Somateria spp.) (Figure 2.2A). In contrast to North America, where spring prevalence of LPAIV in waders is high, low prevalence is reported in a few species of waders in Europe, and no LPAIV has been isolated from waders in Europe to date.
FIGURE 2.2 Avian host range of LPAIV and HPAIV H5N1 in Europe. A. Prevalence of LPAIV infection in the most frequently infected wild waterbirds in Europe (N is the number of birds sampled). B. Wild bird species most frequently found infected with HPAIV H5N1 in Europe (the number of affected European countries experiencing outbreaks of HPAIV H5N1 infection in each wild bird category is indicated).
Higher prevalence of LPAIV infection is reported in juvenile birds than in adults, in particular just before the autumn migration when many waterbirds congregate. LPAIV prevalence was 13% and 6% in 3982 juvenile and 1363 adult dabbling ducks, respectively, sampled during autumn in the Netherlands and Sweden between 1998 and 2006(4,10). Juvenile birds are immunologically naive to LPAIV, and this probably contributes to the observed age difference in LPAIV prevalence1-4). Gender does not appear to affect LPAIV prevalence in birds.
Waterbird ecology probably contributes to the observed higher prevalence of LPAIV infection in birds of the orders Anseriformes and Charadriiformes. Feeding behaviour, notably of dabbling ducks, and social behaviour and population sizes of most waterbird species will probably favour virus transmission)4). Of particular interest is the difference in the prevalence of LPAIV infection in dabbling ducks and small diving ducks (Aythya spp.), possibly resulting from differences in diet and feeding behaviour.
Dabbling ducks feed on the surface of the water whereas diving ducks feed below the water surface. Feeding on the surface of water may result in greater ingestion of contaminated water. Conversely, lower prevalence in swans and geese may reflect a lower risk of infection from grazing. In this light, high prevalence in shelducks and marine diving ducks is rather surprising, as these birds respectively graze and feed in deep saline waters that do not favour LPAIV environmental persistence. Therefore, additional factors probably contribute to the host range of LPAIV in waterbirds.HPAIV H5N1 also have a wide avian host range, but infect some species that are rarely found infected with LPAIV (Figure 2.2B). In particular, birds not necessarily associated with aquatic habitats, such as corvids and birds of prey, were infected, this probably as a result of feeding on infected carcasses. More than 700 fatal cases of HPAIV H5N1 infection were confirmed in wild birds in Europe in 2006, mostly in mute swans (Cygnus olor), whooper swans (C. cygnus), common pochards (Aythya ferina) and tufted ducks (A. fuligula). In summer 2007, a massive outbreak reoccurred in wild birds in Germany and affected mostly black-necked grebes (Podiceps nigricollis) and greatcrested grebes (P. cristatus), killing more than 200 individuals^1). There are few data on potential correlations of HPAIV H5N1 infection with bird age or sex.
ENVIRONMENTAL FACTORS
LPAIV prevalence displays important geographical and seasonal variations1-1,12). In Europe, there may be a north— south gradient of LPAIV prevalence in wild birds, being typically higher in Scandinavian countries than in the more southerly located Netherlands1-4). However, prevalence data remain limited, and it is not known whether seasonal variations in wild bird demography and distribution play a role.
LPAIV infection in wild birds peaks in early autumn, when waterbird populations are composed of a high proportion of juvenile birds that congregate before migration, with a prevalence reaching up to 25%, e.g. in Sweden(10). As migration proceeds, LPAIV prevalence declines and reaches low to undetectable levels during late winter. LPAIV prevalence in spring is typically lower than in autumn, yet can reach relatively high levels in Sweden (up to 9.5%)(10). LPAIV prevalence in waterbirds at breeding grounds has rarely been reported. It was 3.5% in breeding dabbling ducks in northern Siberia(13).
HPAIV H5N1 emerged in wild birds in Europe at the time of autumn migration in 2005 and resulted in massive outbreaks during winter 2005—2006 and summer 2007. HPAIV H5N1 was isolated in wild birds during winter 2007—2008 and winter 2008—2009, displaying an unusual seasonality pattern. During winter 2005—2006, the temporal and geographical distribution of wild birds infected with HPAIV H5N1 coincided with the 0° C isotherm. This suggests that cold weather may have favoured HPAIV H5N1 transmission in Europe, either owing to the aggregation of waterbirds in ice-free areas, or to the persistence of the virus in the environment along the 0°C isotherm, or both(2,14).
Epidemiological role of the
AFFECTED SPECIES
Because of the observed high prevalence of LPAIV infection in birds of the orders Anseriformes and Charadrii- formes, these species (and possibly coots of the order Gruiformes) are typically referred to as the natural reservoirs of LPAIV(12,1). Many species within these orders migrate over long distances and have the potential to spread LPAIV over large geographical areas. Mallards and common teal have a higher prevalence rate than other waterbird species, and probably play a significant role as reservoirs. Annual turnover of mallards in Europe is estimated to be one-third, thus resulting in populations with one-third of juvenile naive individuals, which will favour the maintenance and spread of LPAIV(4,10). However, it is difficult to assess the role of other waterbird species: part or all of the waterbird community may play a role as the reservoir of LPAIV.
LPAIV of the H13 and H16 subtypes form a separate category. These viruses are phylogenetically distinguishable from other LPAIV, and are typically isolated from gulls, belonging to the order Charadriiformes(15). Interestingly, the receptor-binding site of the HA of subtypes H13 and H16 differs from those of other LPAIV subtypes, possibly in adaptation to slightly different cell receptors in gulls(9). Collectively, these findings strongly indicate that gulls are reservoir hosts for these subtypes.
Waders, of the order Charadriiformes, do not appear to play a significant role as reservoir hosts of LPAIV in Europe, contrary to what is observed in North America. Birds from orders other than Anseriformes and Charadrii- formes are typically considered accidental hosts. However, more comprehensive surveillance studies extending to a larger number of wild bird species are necessary to determine the relative role of the different avian hosts of LPAIV.
Wild birds potentially play different roles in the epidemiology of HPAIV H5N1: sentinel, spreader and reservoir species. Some wild bird species, e.g. mute swans, whooper swans and tufted ducks, proved highly susceptible to developing fatal disease, and thus were found to be sentinels. Other wild bird species, e.g. mallards, became subclinically infected and shed virus for several days upon experimental infection, and may thus play a role in spreading the virus. There also is field evidence for the role of wild birds as spreaders of HPAIV H5N1. First, a healthy pochard and a healthy mallard have been found infected in Switzerland and Germany. Second, outbreaks of HPAIV H5N1 in Western Europe during winter 2005-2006 occurred mainly in wild birds. Third, the temporal and geographical association between the locations of these outbreaks and the 0°C isotherm strongly suggest that wild waterbirds were spreading HPAIV H5N1, following its introduction (2,14). Whether wild birds can act as a reservoir of HPAIV H5N1 is an important but unanswered question, especially if eradication of HPAIV H5N1 in poultry succeeds.
TRANSMISSION
LPAIV are excreted mainly from the digestive tract of infected birds into the environment. This may involve large quantities of virus: cloacal shedding was estimated in one study at more than 108 EID50 (median egg infectious dose) per gram of duck faeces, for a total of 1010 EID50 per day(16). The viruses are thought to be mainly transmitted via the faecal-oral route, via ingestion of contaminated water (Figure 2.3A). Faecal-oral transmission is facilitated in aquatic habitats. LPAIV have been shown to persist for several months in environmental reservoirs, such as lake water(17). Birds feeding on surface water may be more likely to be infected. Taken together, these factors favour waterbirds over terrestrial birds as the main hosts of avian influenza viruses.
Alternative routes of transmission are via the cloacal and respiratory routes. ‘ Cloacal drinking’, i.e. uptake of water via the cloaca, has been suggested as a possible route of infection in waterbirds. The respiratory tract can become transiently infected by LPAIV, and excretion of LPAIV from the respiratory tract cannot be excluded.
The mechanisms of year-to-year persistence of LPAIV remain unclear, and several hypotheses have been proposed. On the one hand, year- round infection of wild birds may contribute to LPAIV persistence; LPAIV are usually undetectable during late winter in Europe, but this may be the result of low sampling efforts. On the other hand, environmental persistence of LPAIV may also contribute to their year- round persistence. Surprisingly, few studies have investigated LPAIV persistence in environmental samples, and none in Europe (for a review see Stallknecht and Brown(17)). LPAIV can persist for several months in faeces and water under adequate conditions. The duration of infectivity is highly dependent on LPAIV strains, yet is always higher in water at lower temperatures, with neutral to slightly basic pH and low salinity levels. LPAIV have been isolated from the surface water of lakes in North America and LPAIV RNA has been recovered from lake sediment in North America and from ice of frozen lakes in Siberia. Although infectious virus could not be isolated in the latter two, sediment and ice may form long-term environmental reservoirs of LPAIV LPAIV may survive in lake water during winter, and infect birds heading towards their breeding grounds during spring migration.
Transmission from the virus reservoir in the environment may be independent of the presence of infectious birds, i.e. density- independent, because infectious virus persists in the water after the infectious bird has left; or it may be density- dependent, necessitating the presence of infectious birds. Studies of LPAIV dynamics in wild birds are rare, and density- dependent transmission is typically used as the main mode of transmission in mathematical models of the SIR (susceptible- infected- r ecovered) type. However, a few recent mathematical models of LPAIV transmission have incorporated both a density-dependent and density-independent form of transmission. The results of these models increasingly support an important role for environmental transmission of persisting LPAIV in the absence of infectious birds in maintaining LPAIV locally and from year to year(18’19).
The transmission mode of HPAIV H5N1 may be different from that of LPAIV. In contrast to LPAIV, HPAIV H5N1 are mainly excreted from the respiratory tract of infected birds. This may favour direct bird-to-bird transmission via respiratory secretions (Figure 2.3B). However, contamination of the environment by respiratory secretions and infected carcasses probably results in indirect oral transmission of the virus. The presence of HPAIV H5N1 in one mallard in January 2009 in Germany, without any report of additional outbreaks in Europe, may indicate environmental persistence of HPAIV H5N1, which have been shown to remain infective for 158 days in fresh water at 17° C, and for 26 days at 28° C(20). In addition, bird species not necessarily associated with aquatic habitats were also found infected with HPAIV H5N1, notably corvids and birds of prey. This strongly suggests that predation and ingestion of infected bird carcasses is another mode of transmission of HPAIV H5N1 in wild birds.
PATHOGENESIS, PATHOLOGY AND IMMUNITY
The pathogenicity of avian influenza viruses in poultry is at least partly determined by the cleavability of the HA protein. The HA glycoprotein contributes to attachment and fusion of the lipid membranes of the virus and host cell. The HA protein of AIV must be cleaved by host proteases to become infectious and undergo a conformational change at low pH, leading to the fusion of the virus and host cell membranes. The HA protein of LPAIV is cleaved by extracellular trypsin-like host proteases present in the digestive and respiratory tracts of birds. By contrast, the HA protein of HPAIV has a polybasic cleavage site that can be cleaved by ubiquitous intracellular subtilisin- like host proteases such as furin, which are present in many cell types, contributing to high virulence.
Ingestion of water contaminated with LPAIV is considered the main route of infection in waterbirds(21), although ‘cloacal drinking’ and inhalation cannot be excluded. Thus, the first sites of LPAIV replication are the intestinal epithelium and to a lesser extent the respiratory epithelium (Figure 2.3A). If ingested, LPAIV must pass through and survive the acidic environment of the proventriculus of ducks; uncleaved HA of LPAIV appears more resistant to acidic pH than that of mammalian influenza viruses, and ingestion of a large amount of water may partly neutralize the acidic pH of duck stomach, which together probably allow infectious doses of LPAIV to reach their target site of replication in the intestine.
Little is known about the infectious dose of LPAIV in wild birds. Doses ranging between 101.2 and 103.3 EID50 have proven to be infectious(22), but higher doses were used in the past for experimental infections. Large quantities of LPAIV are shed in infected bird faeces(16), and LPAIV have been isolated from unconcentrated samples of lake water; thus ingestion of relatively high infectious doses may not be infrequent in nature.
The route of infection of HPAIV H5N1 may be inhalation of respiratory secretions or ingestion of water contaminated by respiratory secretions and carcasses. Ingestion of infected carcasses is another route of infection, based on the occurrence of disease in corvids and birds of prey (Figure 2.3B). Ingestion of meat infected with HPAIV H5N1 resulted in infection and disease in experimentally infected herring gulls (Larus argentatus')1'i∖ The first site of HPAIV H5N1 replication appears to be the respiratory epithelium. In contrast to LPAIV infection, cells along the intestinal epithelium are not, or are rarely reported to be, infected by HPAIV H5N1. A possible explanation is that HPAIV H5N1 is unable to survive the acid barrier of the proventriculus: HPAIV H5N1 is released from the host cell with cleaved HA protein, which is known to be sensitive to, and lose infectivity at, low pH[4].
Relatively low infectious doses of HPAIV H5N1 (around 104 median tissue-culture infectious dose (TCID50)) result in productive infection of various wild bird species(23-25). Inoculation of various doses of HPAIV H5N1 into mute swans(26), house sparrows (Passer domes- ticus) and rock pigeons (Columbia lVα)(2τ) demonstrated a correlation between infectious dose, clinical outcome and duration of viral shedding. Lower doses typically resulted in less severe disease and longer viral shedding, although the total amount of excreted virus did not differ substantially in birds inoculated with different infectious doses.
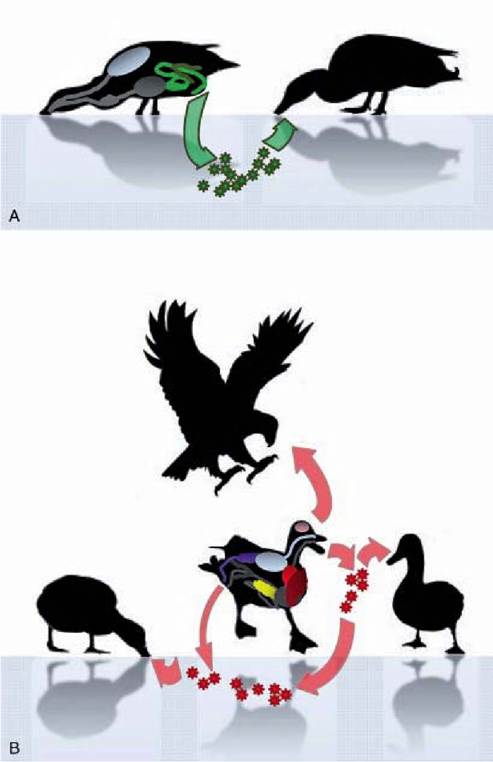
FIGURE 2.3 Routes of transmission and tissue tropism of LPAIV and HPAIV H5N1 in wild birds. A. LPAIV (green stars) are thought to be mainly transmitted via a faecal-oral route, and ingestion of contaminated water is considered the main route of infection in wild birds. LPAIV are recovered from the respiratory tract (blue) and digestive tract (green), infecting mostly intestinal epithelial cells. Infected birds do not present clinical signs. B. HPAIV H5N1 (red stars) are mostly shed via the respiratory tract, yet the exact route of infection in wild birds is not exactly known, and probably includes inhalation, and ingestion of contaminated water or infected carcasses. HPAIV H5N1 replicate in many organs in highly susceptible species, including the central nervous system (pink), the respiratory tract (blue), the heart (red), the liver (brown), the pancreas (yellow) and the kidneys (purple). In susceptible species, clinical signs include laboured breathing and neurological signs, and disease is often fatal. The intestinal tract (grey) is not infected.
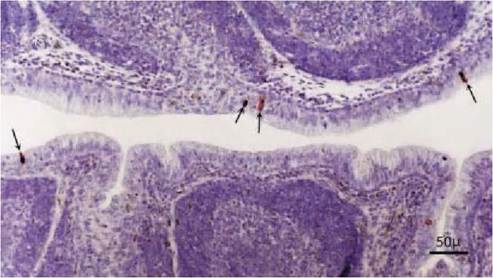
FIGURE 2.4 Cloacal bursa of a wild mallard duck naturally infected with LPAIV, subtype H2N3. Positive cells are scattered along the surface epithelium (arrows). Note the lack of histologic changes associated with the expression of influenza virus antigen in Cloacal bursa (bursa of Fabricius) of a wild mallard. Immunoperoxidase method for nucleoprotein of AI virus counterstained with hematoxylin. Reprinted with permission from Daoust P-Y et al., 2001.
mal cells are the main target for HPAIV H5N1 replication, e.g. respiratory epithelial cells, pancreatic parenchymal cells, hepatocytes, adrenocortical cells, renal epithelial cells, myocytes and neurons. In contrast to HPAIV H5N1 infection in chickens, HPAIV H5N1 rarely, if ever, infects endothelial cells in most wild bird species. Exceptions are mute and whooper swans and tufted ducks(26’33’34), in which HPAIV H5N1 shows a clear endotheliotropism.
In contrast to LPAIV, HPAIV H5N1 is shed more from the respiratory tract than from the digestive tract, based on virus titres in pharyngeal and cloacal swabs, respectively. Pharyngeal shedding of HPAIV H5N1 starts at least 1 dpi and can last 5-7 days in experimental infection of most European species of wild ducks and swans(24’25). In mute swans infected with low doses, HPAIV H5N1 were shed for up to 11 days[5]. In contrast, herring gulls shed HPAIV H5N1 for up to 3 days only(23). Virus shedding from the cloaca is typically shorter and at titres of one to two logs lower than from the pharynx(23-25’[6] [7] [8].
AIV infection in wild birds is typically acute and resolves with the development of a specific immune response. Regeneration of intestinal epithelial cells of wild birds infected with LPAIV is rapid, with histological evidence of regeneration as soon as 72 hours post-infection[9]. Damage caused by HPAIV H5N1 infection varies greatly among species.
Disease following infection with AIV may result from direct cytopathogenic damage caused by the virus replicating in host cells, from damage resulting from the host immune response, or from a combination of these. LPAIV infection in domestic ducks and wild birds causes no, or at most very mild, clinical disease, and the only recorded lesions are very mild degenerative changes of intestinal and bursal epithelium (see below).
By contrast, the effect of HPAIV H5N1 infection in wild birds ranges from subclinical disease to acute death, largely dependent on host species affected. Replication of HPAIV H5N1 in parenchymal cells of an organ results in marked necrosis and inflammation. Clinical disease is highly variable in timing and presentation. It may start between 4 and 9 dpi, and involve respiratory, nervous or general signs. Development of disease differs in those species, such as whooper swans, in which HPAIV H5N1 is endotheliotropic. Infection of endothelial cells causes blood vessel damage and results in haemorrhages and oedema, leading to clinical disease as early as 24 hours post-infection.
LPAIV infection in wild birds generally does not cause gross or microscopic lesions (Figure 2.4). However, very
TABLE 2.3 Infectious period, incubation time and clinical signs following natural and experimental infection with LPAIV and HPAIV
| H5N1 of the sublineage that spread to bird populations in Europe, in wild bird species. | |||||||
| Wild bird species | AIV | Type of infection | Maximum pharyngeal shedding (duration; N/ Total)* | Maximum cloacal shedding (duration; N/Total)* | Clinical signs (incubation time; N/Total) | Mean time to death (N/Total) | Ref |
| Mallard | LPAIV | Natural | ND | ND (3-8 days; 59) | None | None | 32 |
| Mallard | LPAIV | Experimental | 5.9 (7 days; NA) | 5.8 (7 days; NA) | None | None | 28,16,29,30,31,32 |
| Mallard | HPAIV H5N1 | Experimental | 3.2 (4 days; 8/8) | 0 | None | None | 25 |
| Common teal | HPAIV H5N1 | Experimental | 1 (5 days; 3/8) | 0.5 (1 day; 1/8) | None | None | 25 |
| Eurasian wigeon | HPAIV H5N1 | Experimental | 1.2 (2 days; 4/8) | 0 | None | None | 25 |
| Gadwall | HPAIV H5N1 | Experimental | 2.2 (6 days; 7/8) | 0 | None | None | 25 |
| Tufted duck | HPAIV H5N1 | Experimental | 3.2 (5 days; 6/8) | 0 | Laboured breathing, increased recumbency Neurologic signs: torticollis, circling, loss of balance, head tremors (3-4 days; 7/8) | 4 days (3/8) | 25 |
| Tufted duck | HPAIV H5N1 | Natural | ND | ND | Neurological signs: head tilt, circling, loss of balance, drooping wings | ND | 34 |
| Common pochard | HPAIV H5N1 | Experimental | 2.8 (4 days; 7/8) | 0.7 (2 days; 2/8) | Laboured breathing, increased recumbency Neurologic signs: torticollis, circling, loss of balance, head tremors (3-4 days; 4/8) | 4 days (1/8) | 25 |
| Mute swan | HPAIV H5N1 | Experimental | 5.6 (7 days; 5/5) | 4.5 (4 days; 5/5) | Listlessness Neurological signs: seizures, tremors, marked inco-ordination (5-7 days; 5/5) | 5-8 days (5/5) | 24 |
| Whooper swan | HPAIV H5N1 | Experimental | 6.3 (6 days; 4/4) | 4.2 (5 days; 4/4) | Listlessness Neurological signs: seizures, tremors, marked inco-ordination (2-4 days; 4/4) | 4 days (4/4) | 24 |
| Whooper swan | HPAIV H5N1 | Experimental | 4.2 (based on Ct values) (6—11 days; 12/12) | 3.3 (based on Ct values) (6-10 days; 12/12) | Neurological signs: opisthotonus, torticollis, ataxia (4-7 days; 3/12) Sudden death (4-7 days; | 5-9 days (5/5) 8-14 days (6/7) | 26 |
3/12)
TABLE 2.3 (Continued)
| Wild bird species | AIV | Type of infection | Maximum pharyngeal shedding (duration; N/ Total)[†††] [‡‡‡] | Maximum cloacal shedding (duration; N/Total)* | Clinical signs (incubation time; N/Total) | Mean time to death (N/Total) | Ref |
| Herring gu∏ | hpaiv H5N1 | Experimental | 3.8—4.3 (2.5—3 days; 5/5) | < 1.96 (1.5—3 days; 5/5) | Lethargy Neurological signs: seizures, head-tilt, head tremors, torticollis, severe imbalance (1—5 days; 5/5) | 5 days (3/5) | 23 |
| House sparrow | HPAIV H5N1 | Experimental | 5 (4—9 days; 13/15) | 4.3 (4—9 days; 13/15) | Lethargy, pulled-up feathers Neurological signs: loss of balance, head tremors, seizures Sudden death (4—7 days; rare) | 5—8 days (13/15) | 27 |
| HPAIV H5N1 | Experimental | bgcolor=white> | most parenchyma | Parenchymal cells of other organs | |||
| Whooper | HPAIV | Experimental | ND | Multifocal to coalescing necrosis with mild to | Neurons, astrocytes, and glial | 24 | |
| swan | H5N1 | moderate heterophilic inflammation in brain and | cells | ||||
| most parenchyma | Parenchymal cells of other organs | ||||||
| Whooper | HPAIV | Experimental | Widespread haemorrhages | Necrotizing rhinitis | Respiratory epithelial cells | 26 | |
| swan | H5N1 | (myocardium, brain, lungs) | Multifocal pancreatic necrosis | Endothelial cells | |||
| and petechiae in pancreas, | Multifocal hepatic necrosis | Pancreatic cells | |||||
| liver | Multifocal encephalitis | Hepatocytes | |||||
| Pancreatic necrosis | Necrotizing dermatitis | Neurons, glial cells and ependymal cells Adrenocortical cells | |||||
| Mute and | HPAIV | Experimental | Pulmonary oedema and | Hemorrhagic broncho-interstitial pneumonia | Respiratory epithelial cells | 33 | |
| whooper | H5N1 | congestion | Multifocal pancreatic necrosis | Endothelial cells | |||
| swan | Multifocal hemorrhagic | Multifocal to coalescing coagulative hepatic necrosis | Pancreatic cells | ||||
| pancreatic necrosis | Multifocal Iymphoplasmacytic encephalitis | Hepatocytes | |||||
| Subepicardial haemorrhages | Spleen | Neurons, glial cells and | |||||
| Multifocal lymphocyte necrosis in spleen and Peyers | ependymal cells | ||||||
| patches | Intestinal epithelial cells | ||||||
| Multifocal adrenalitis | (proventriculus) Mononuclear cells (Peyers patches) Adrenocortical cells Kidney tubular epithelial cells | ||||||
| Herring | HPAIV | Experimental | ND | Multifocal pancreatic necrosis | Pancreatic cells | 23 | |
| gull | H5N1 | Multifocal encephalitis | Neurons and glial cells | ||||
| Heterophilic and necrotizing myocarditis | Cardiomyocytes | ||||||
| Multifocal necrotizing adrenalitis | Adrenocortical cells | ||||||
| House | HPAIV | Experimental | Pulmonary oedema and | Multifocal to coalescing pancreatic necrosis | Pancreatic cells | 27 | |
| sparrow | H5N1 | congestion | Multifocal to coalescing necrotizing adrenalitis | Adrenocortical cells | |||
| Pancreatic necrosis | Lymphocytic air sacculitis | Neurons and glial cells | |||||
| Splenomegaly | Lymphoplasmacytic and necrotizing encephalitis | Cardiomyocytes | |||||
| Lymphocytic ganglioneuritis | Hepatocytes and Kupffer cells | ||||||
| Myocardial necrosis and Iymphoplasmacytic | Kidney tubular epithelial cells | ||||||
| myocarditis | Thecal epithelial cells, Sertoli | ||||||
| Lymphocytic and heterophilic nephritis | cells, testis tubular epithelial | ||||||
| Lymphocytic orchitis | cells | ||||||
| Heterophilic oophoritis Necrotizing myositis | Myocytes | ||||||
| Rock | Pancreatic necrosis | Multifocal lymphocytic encephalitis and | Neurons and glial cells | 27 | |||
| pigeon | meningoencephalitis | bgcolor=white>Hepatocytes||||||
| Multifocal hepatic necrosis | Thecal epithelial cells |
ND - not determined
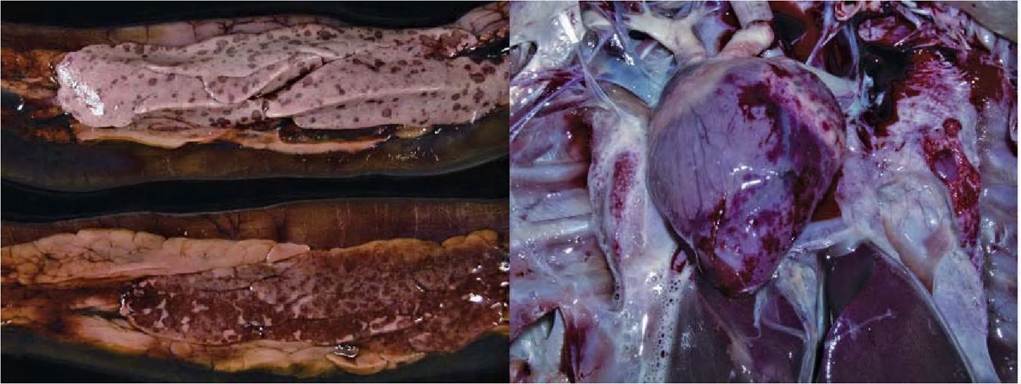
FIGURE 2.5 Gross lesions of HPAIV H5N1 infection. Left panel: Pancreas showing multiple foci of necrosis and haemorrhage in a whooper swan naturally infected with HPAIV H5N1. Right panel: Heart showing subepicardial haemorrhages in a whooper swan naturally infected with HPAIV H5N1. Reprinted with permission from Teifke, J.P. et al., 2007.
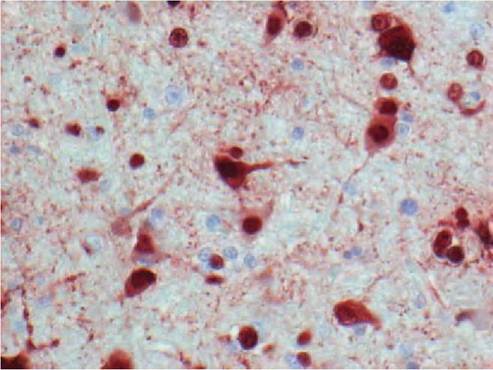
FIGURE 2.6 Histological lesions of HPAIV H5N1 infection. Severe encephalitis in a tufted duck experimentally infected with HPAIV H5N1, characterized by abundant influenza virus expression in neurons and glial cells (stained by immunohistochemistry that used a monoclonal antibody against the nucleoprotein of influenza A virus as primary antibody). Reprinted with permission from Keawcharoen, J. et al., 2008.
annual reinfection. However, lower prevalence of LPAIV infection in adult waterbirds strongly suggests that immunity, in particular heterosubtypic immunity, may reduce the susceptibility of older birds to reinfection.
Immunity to LPAIV protected domestic ducks and mallards against disease and greatly reduced virus shedding upon challenge with the same subtype(29'38). Immunity to another subtype of AIV also reduced disease and virus shedding from challenge with HPAIV H5N1, but to a lesser degree(39).
CLINICAL SIGNS
In general, neither natural nor experimental LPAIV infection causes clinical signs in wild birds(40). Likewise, experimental infection of domestic ducks and mallards with LPAIV typically results in subclinical infection. Despite the general absence of clinical disease, it has recently been suggested that LPAIV infection may cause physiological costs to wild birds(41). Natural LPAIV infection of two Bewick’s swans (Cygnus columbianus bewickii) was associated with reduced feeding and migrating performances, resulting in shorter and delayed spring migration1-42). Mallards, naturally infected with LPAIV, were significantly lighter than uninfected mallards, and virus titres negatively correlated with body mass of juvenile birds(32). It remains unknown whether LPAIV infections are the cause or consequence in these observed correlations.
I n contrast to LPAIV, the clinical effects of HPAIV H5N1 infection of wild birds range from subclinical infection to sudden death, both in natural and experimental infections. In natural infections, clinical signs are rarely reported, because most infected wild birds are found dead. In Sweden, seven naturally infected tufted ducks were found alive and presented with neurologic signs, including head tilt, circling, loss of balance and drooping wings(34).
In experimental infections with HPAIV H5N1, clinical signs depend largely on host species but also on virus strain and inoculation dose(23-27). Mute swans, whooper swans and house sparrows showed severe clinical signs and a high mortality rate, whereas dabbling duck species — mallard, common teal, Eurasian wigeon (Anas penelope), gadwall (A. strepera) — showed no clinical signs and no deaths. Rock pigeons, diving ducks — tufted duck, common pochard — and herring gulls were intermediate in severity of clinical signs and death rate. Mean time to death ranged from 4 to 14 dpi (Table 2.3). Clinical signs were first seen between 2 and 7 dpi. Clinical signs were lethargy, weakness, listlessness, dyspnoea and neurological signs, such as loss of balance, incoordination, circling, ataxia, opisthotonos, head tilt, head tremors, torticollis and seizures1-23-27), and sudden deaths with no clinical signs were also reported (26,43). Neurological signs were particularly conspicuous and were associated with cerebellar infection in experimentally infected birds.
DIAGNOSIS
LPAIV infection is generally subclinical, and the diagnosis in a live wild bird can be made by virus culture or by reverse transcription polymerase chain reaction (RT- PCR)(12,44). Samples of choice are cloacal swabs and, to a lesser extent, pharyngeal swabs, which are placed in virus transport medium and either analysed directly or stored at —70°C until analysis. For virus isolation, specific pathogen- free embryonated chicken eggs are more sensitive than cell culture. Virus culture allows for full characterization of the virus isolate. For RT-PCR, primer pairs based on a conserved gene, such as the matrix gene, are used for initial detection. This may be followed by RT-PCR developed to detect specific HA subtypes, such as H5 and H7, or by virus culture.
The diagnosis of LPAIV infection in a dead wild bird can be made by virus culture, RT-PCR or immunohisto- chemistry1-12,44). Besides cloacal and pharyngeal swabs, samples of choice for virus culture and RT-PCR are tissue samples from intestine, bursa of Fabricius and lung. For immunohistochemistry, a primary antibody against one or more AIV proteins is used to detect AIV antigen in infected cells(45). Immunohistochemistry makes it possible to colocalize microscopic lesions with the presence of AIV antigen, but only allows a diagnosis to the genus level. Samples of choice for immunohistochemistry are tissue samples from the same organs as above, but fixed in 10% neutral-buffered formalin. Only fresh tissues should be used, because AIV antigen expression is limited to epithelial cells, which are the first to desquamate during autolysis of the carcass.
HPAIV H5N1 infection should be included in the differential diagnosis of any unexplained mortality of wild birds, particularly of the species known to be susceptible to disease from these viruses, and also when birds show neurological signs. Other diseases to consider in such cases include Newcastle disease, botulism, avian cholera and blue-green algal poisoning(46). The gross and microscopic lesions are too non-specific to make a presumptive diagnosis, but suspicion is increased with the gross observations of multiple necrotic foci in the pancreas and — in swans — multi-organ haemorrhage; and for the microscopic observation of acute necrosis and inflammation in multiple organs, including brain, pancreas, liver and adrenal gland. The differential diagnosis for such microscopic lesions should include Newcastle disease.
The diagnosis of HPAIV H5N1 infection in a live wild bird can be made by virus culture or RT-PCR, as for LPAIV, except that pharyngeal swabs are preferred to cloacal swabs. The diagnosis of HPAIV H5N1 in a dead wild bird can be made by virus culture, RT-PCR or immunohistochemistry, as for LPAIV. Again, pharyngeal swabs are preferred to cloacal swabs. Tissue samples of choice also are different: they should include brain, lung, air sac, pancreas, liver and kidney.
Screening of wild bird populations for both LPAIV and HPAIV is most commonly done by Taqman PCR of pharyngeal and cloacal swabs, by use of a probe for the matrix gene. Positive samples are subsequently analysed by virus culture to obtain the virus, to identify HA and NA subtype and to determine pathogenicity1-12,47).
Other diagnostic tests available for AIV diagnosis include: nucleic acid sequence-based amplification (NASBA), loop- mediated isothermal polymerase chain reaction (LAMP- PCR), pyrosequencing and in situ hybridization for RNA detection; antigen-capture enzyme-linked immunosorbent assay (ELISA) for antigen detection; and agarose gel immunodiffusion (AGID), ELISA, hemagglutinin inhibition (HI) and neuraminidase inhibition (NI) tests for antibody detection1-48).
MANAGEMENT, CONTROL, AND REGULATIONS
LPAIV infection is endemic in wild birds and management or control is not required for the purpose of wild bird health. However, wild birds are the source of LPAIV infecting poultry and potentially evolving into HPAIV (in the case of LPAIV of the H5 and H7 subtypes), which causes extensive mortality and production loss. Management and control measures thus are applied to reduce the risk of transmission of LPAIV from wild bird to poultry populations by improving biosecurity.
Surveillance of wild birds for AIV infection consists of sampling live birds for detection of AIV or AIV RNA, and necropsy of dead birds for detection of AIV, AIV RNA or AIV antigen.
Three main objectives of such a surveillance programme are: i) to identify areas considered at higher risk for introduction and occurrence of AIV in poultry owing to proximity of poultry holdings to mixing and resting areas of migratory birds; ii) to prevent the disease from entering poultry holdings by limiting contact between wild birds and poultry and by enhancing biosecurity measures; and iii) to detect and report AIV of the H5 and H7 subtypes in wildlife, and to adapt disease-prevention measures in areas where they are detected. Prevention measures, such as limiting contact between farmed animals and wild birds, and enhancing biosecurity measures, such as ensuring that the source of water for farmed animals is not used or contaminated by wild birds, are also applied for susceptible species of farmed mammals that can acquire AIV infection from wild birds, such as pigs and mink.
Wild birds historically played no epidemiological role in HPAIV epidemiology. However, they probably contributed to the geographical spread of HPAIV H5N1, and early detection of HPAIV H5N1 currently is one of the functions of AIV surveillance in wild birds. In the event of wild bird die-offs, necropsy of highly susceptible species, such as swans, diving ducks and grebes, should be reinforced. Detection of HPAI H5N1 in wild birds requires European member states to establish a 3-km control area and a 10-km monitoring area, where restrictions on movements of live poultry and poultry products apply, and where hunting of wild birds is banned. Poultry must be kept indoors to prevent contact with infected wild birds. Clinical inspections and laboratory investigations are required in poultry holdings in order to detect possible virus introduction. Biosecurity and disease awareness are raised. The European Commission also approved preventive vaccination programmes for birds kept in zoos in 17 member states to avoid stamping-out measures for captive wild bird species(49).
Domestic cats and other carnivores may be at risk of HPAIV H5N1 infection in areas where outbreaks in wild and domestic birds occur and should be monitored for infection. Measures to prevent contact between domestic carnivores and infected birds, and surveillance of unusual morbidity and mortality in domestic and wild carnivores, should be implemented(50).
PUBLIC HEALTH CONCERN
Transmission of AIV from wild birds to humans is rare. There is serological evidence that people in close contact with wild birds, such as duck hunters, can become infected with LPAIV(51). However, the only reported transmission of AIV from wild birds to humans concerns two clusters of human infection with HPAIV H5N1 and six human deaths in Azerbaijan, where close contact with and de-feathering of infected wild swans was considered to be the most probable source of exposure1-52). Other sources of AIV infection in humans are most commonly from poultry, but also laboratory accident and pre- and post mortem examination of infected seals and possibly wild birds(53).
AIV infection in humans typically causes mild clinical disease, characterized by conjunctivitis or influenza-like illness, which resolves within 2 weeks However, HPAIV H5N1 infection in humans, although rare, causes severe pneumonia that often progresses to acute respiratory distress syndrome, with a case fatality rate of about 60%(53’54).
The protective measures when handling apparently healthy wild birds in areas where HPAIV H5N1 infection is not suspected are routine hygienic measures such as using gloves and protective eyewear when possible, washing hands with soap and water and disinfecting equipment and work surfaces(55). In areas where HPAIV H5N1 infection has been identified in wild birds, wearing protective suits and respiratory masks and disinfecting tools and vehicles must be implemented to limit contamination and chains of transmission(49).
SIGNIFICANCE AND IMPLICATIONS FOR ANIMAL HEALTH
Both LPAIV and HPAIV have had implications for domestic and wild animal health. LPAIV may cross from wild birds to poultry. Poultry may then present with clinical disease upon infection or show reduction in egg or meat production(56). Furthermore, LPAIV of the H5 and H7 subtypes may become highly pathogenic once introduced into poultry populations, potentially bearing high economical costs and a zoonotic risk. Poultry infected with LPAIV may become bridge species, transmitting LPAIV to other species, including domestic animals and humans. LPAIV have also resulted in outbreaks of severe respiratory disease in farmed mink and outbreaks of influenza-like illness in domestic pigs(2), potentially producing high economic costs. LPAIV have been transmitted from wild birds to free- ranging wild mammals and have resulted in outbreaks of severe respiratory disease in harbour seals. LPAIV outbreaks in farmed mink also call for awareness in the event of cross- species transmission of LPAIV to endangered European mink (Mustela lutreola).
Poultry is the major source of HPAIV in other domestic animals and in wild animals. In particular, HPAIV H5N1, which displays an unusually broad host range, have infected and caused severe systemic disease and death in domestic cats and dogs. In addition, HPAIV H5N1 causes death and severe disease in a wide range of wild birds, and in free-ranging mammals, i.e. a stone marten (Martes foina) and an American mink (Neovison vison) in Europe. The potential of AIV to cross the species barrier and to cause disease in novel host species thus calls for continued and extended surveillance of AIV in wild bird populations1-57).
More on the topic AVIAN INFLUENZA:
- INTRODUCTION
- 5 Appendices
- 3 SELECTED SOCIO-ECONOMICALLY IMPORTANT WILDLIFE RELATED PATHOGENS AND DISEASES IN EUROPE
- 2 SELECTED ZOONOTIC PATHOGENS WITH EUROPEAN WILDLIFE RESERVOIRS/HOSTS
- Conclusions
- HERPESVIRUS INFECTIONS IN WILD BIRDS
- Agency Planning to Ensure Continuity of Essential Veterinary Functions during a Pandemic Is Incomplete
- Introduction